
EPA-Enriched Phospholipids Alleviate Renal Interstitial Fibrosis in Spontaneously Hypertensive Rats by Regulating TGF-β Signaling Pathways

Abstract
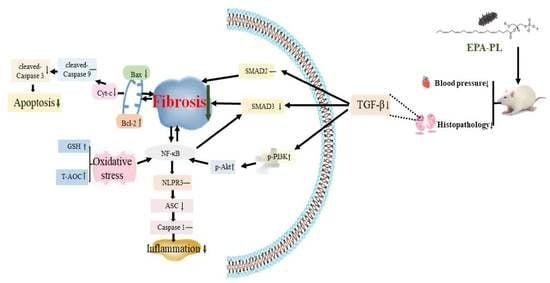
:
1. Introduction
2. Results and Discussion
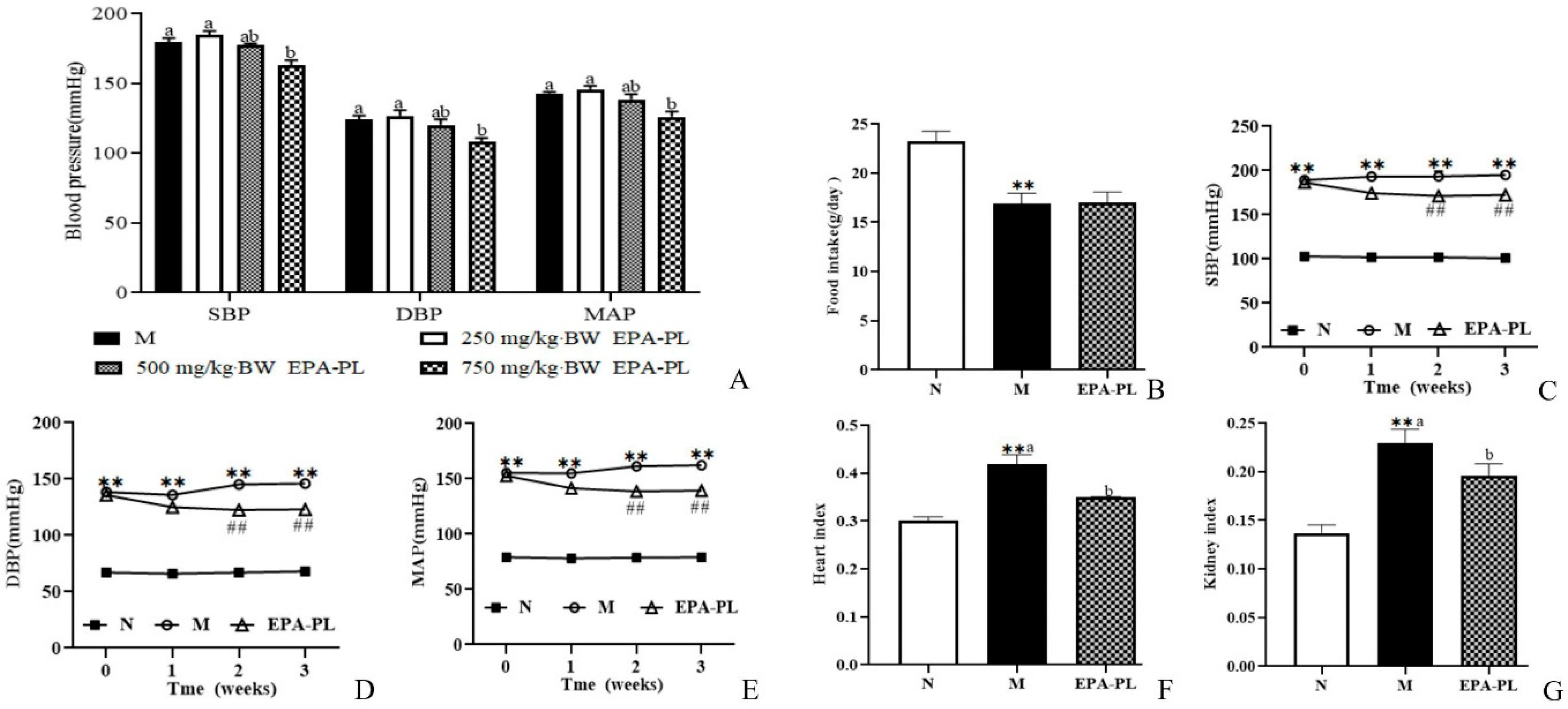
2.1. The Change in Blood Pressure in SHRs after Single Oral Administration of EPA-PL
2.2. The Effects of EPA-PL on Growth Parameters in SHRs
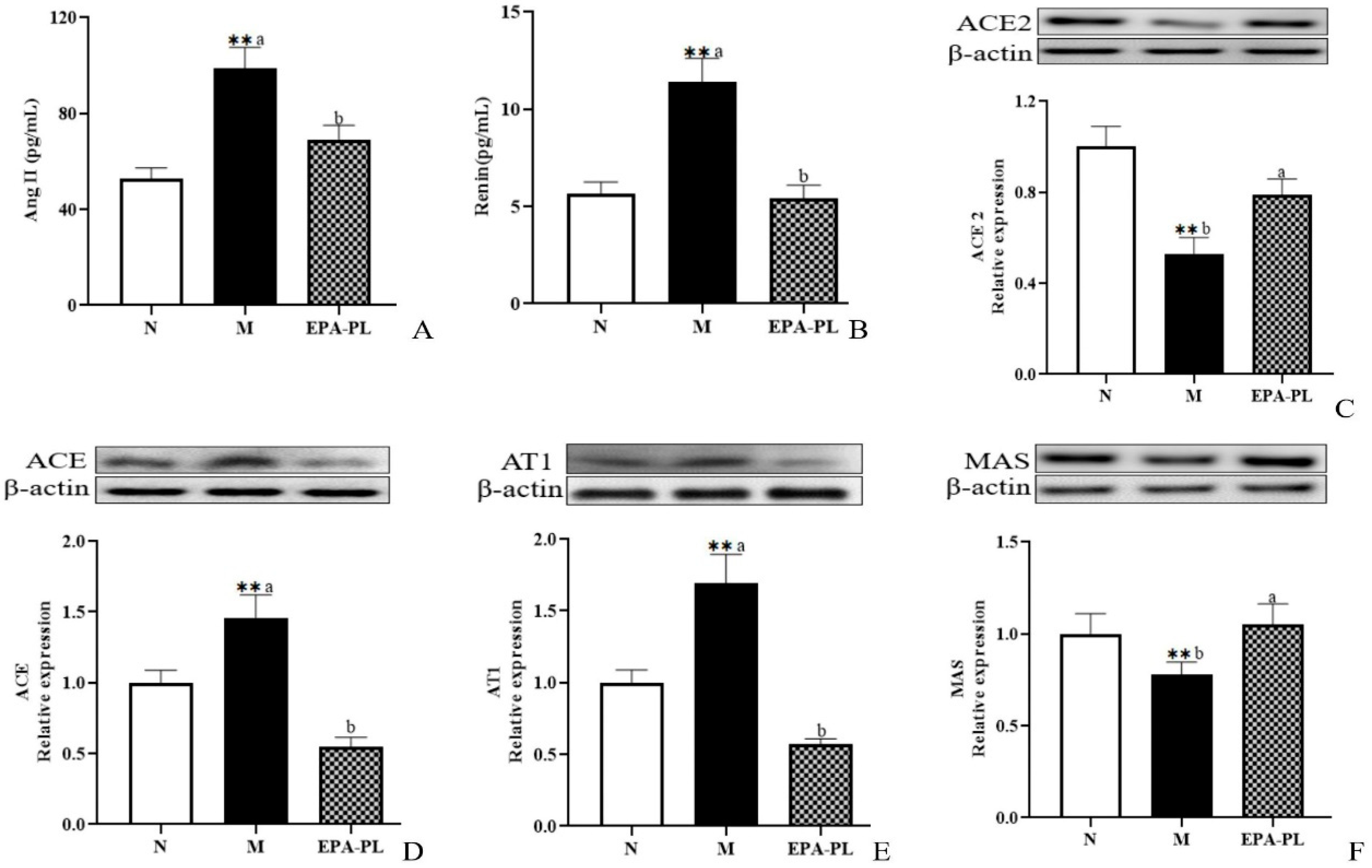
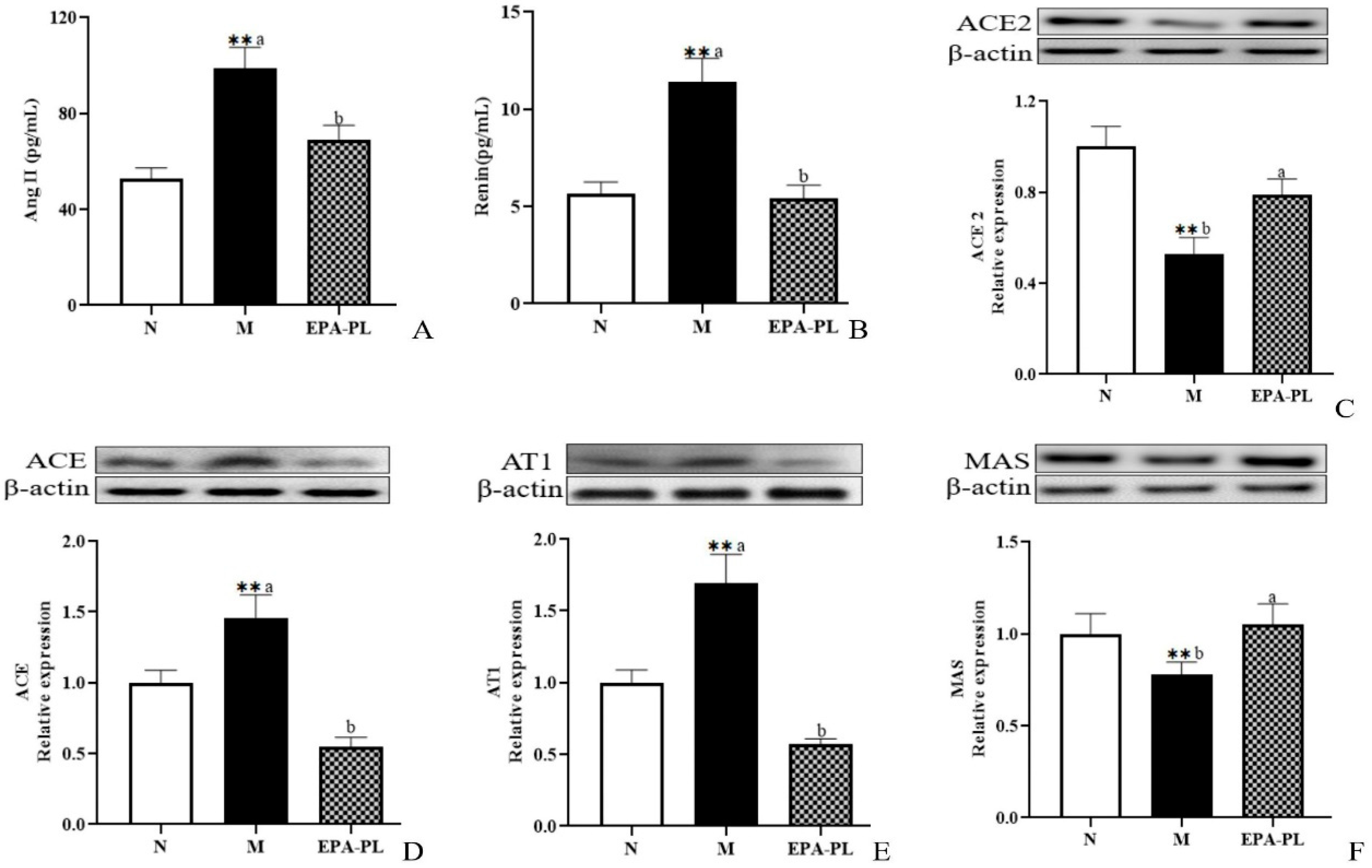
2.3. The Effects of EPA-PL on The Renin–Angiotensin System in SHRs
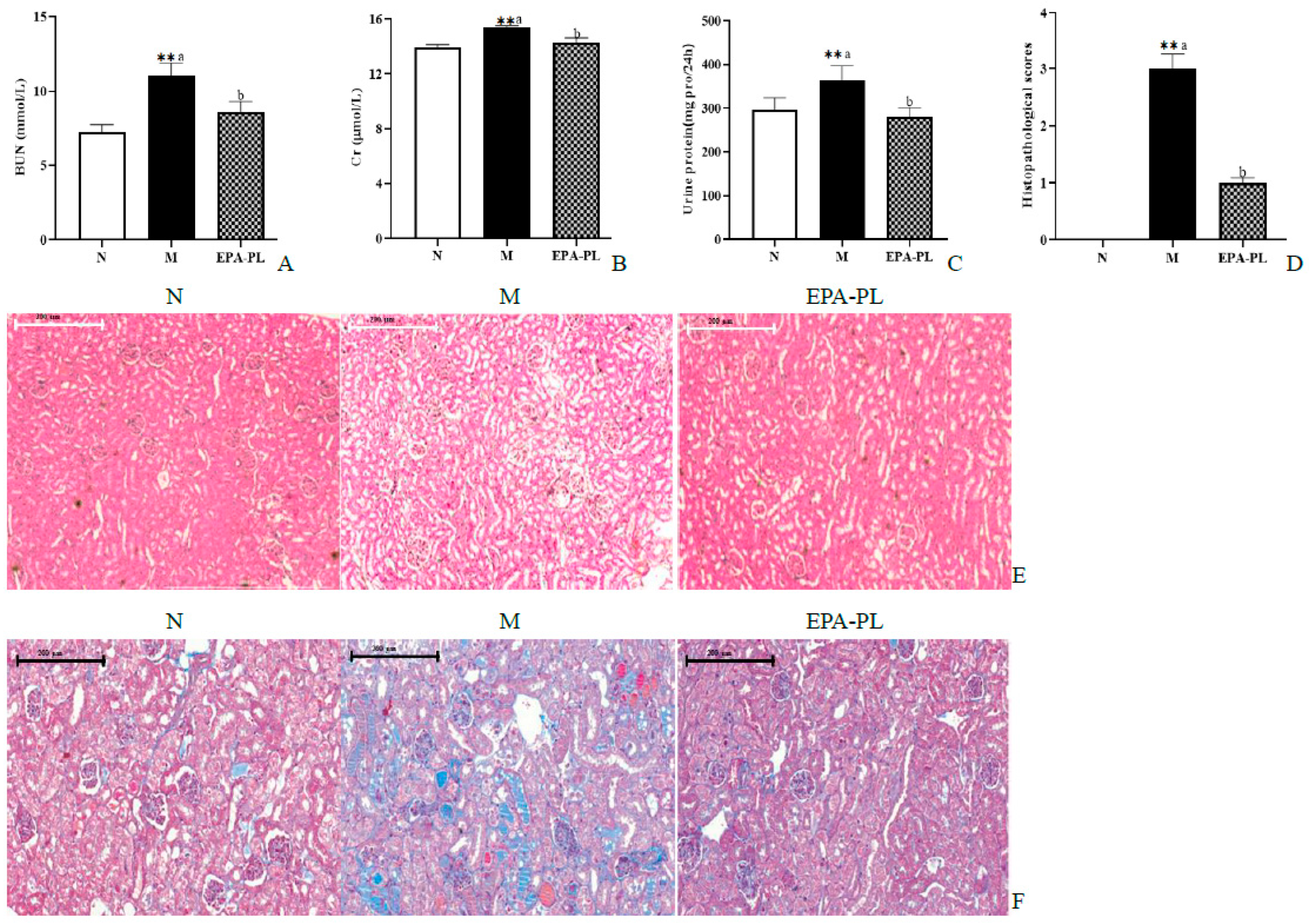
2.4. The Effects of EPA-PL on Serum Parameters and Renal Pathological Changes in SHRs
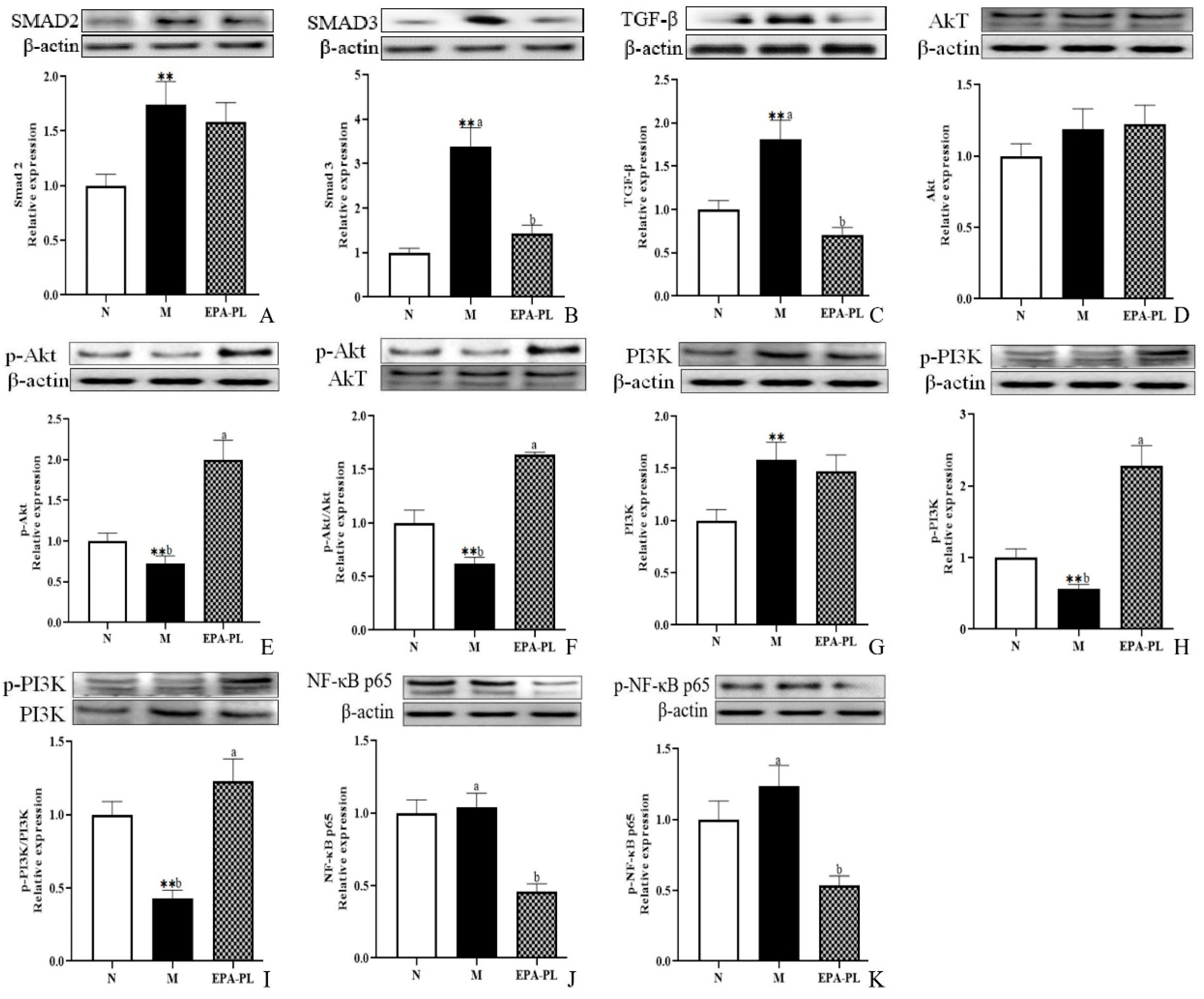
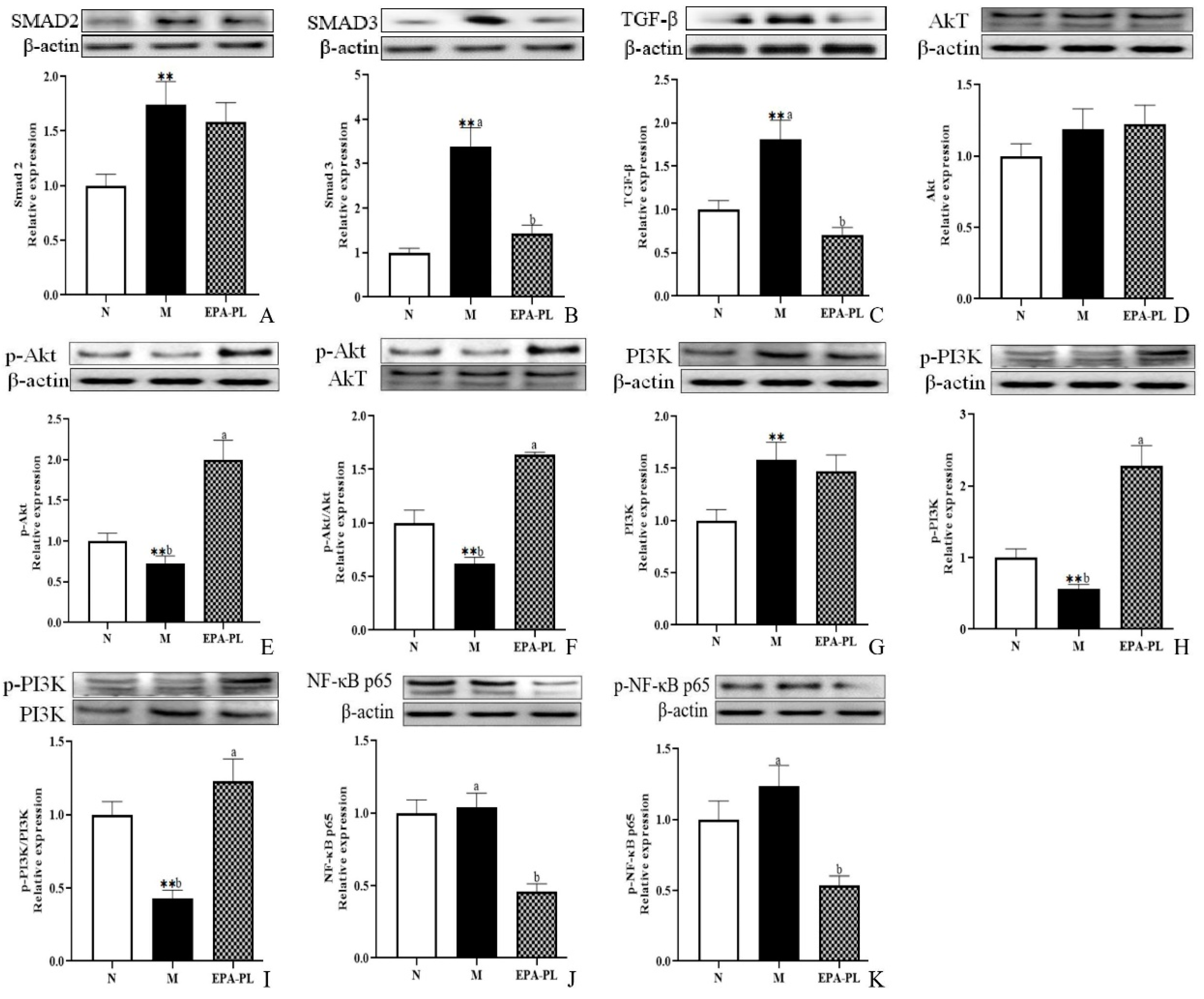
2.5. The Effects of EPA-PL on TGF-β/Smad Signaling Pathway in SHRs
2.6. The Effects of EPA-PL on PI3K/Akt/NF-κB Signaling Pathway in SHRs
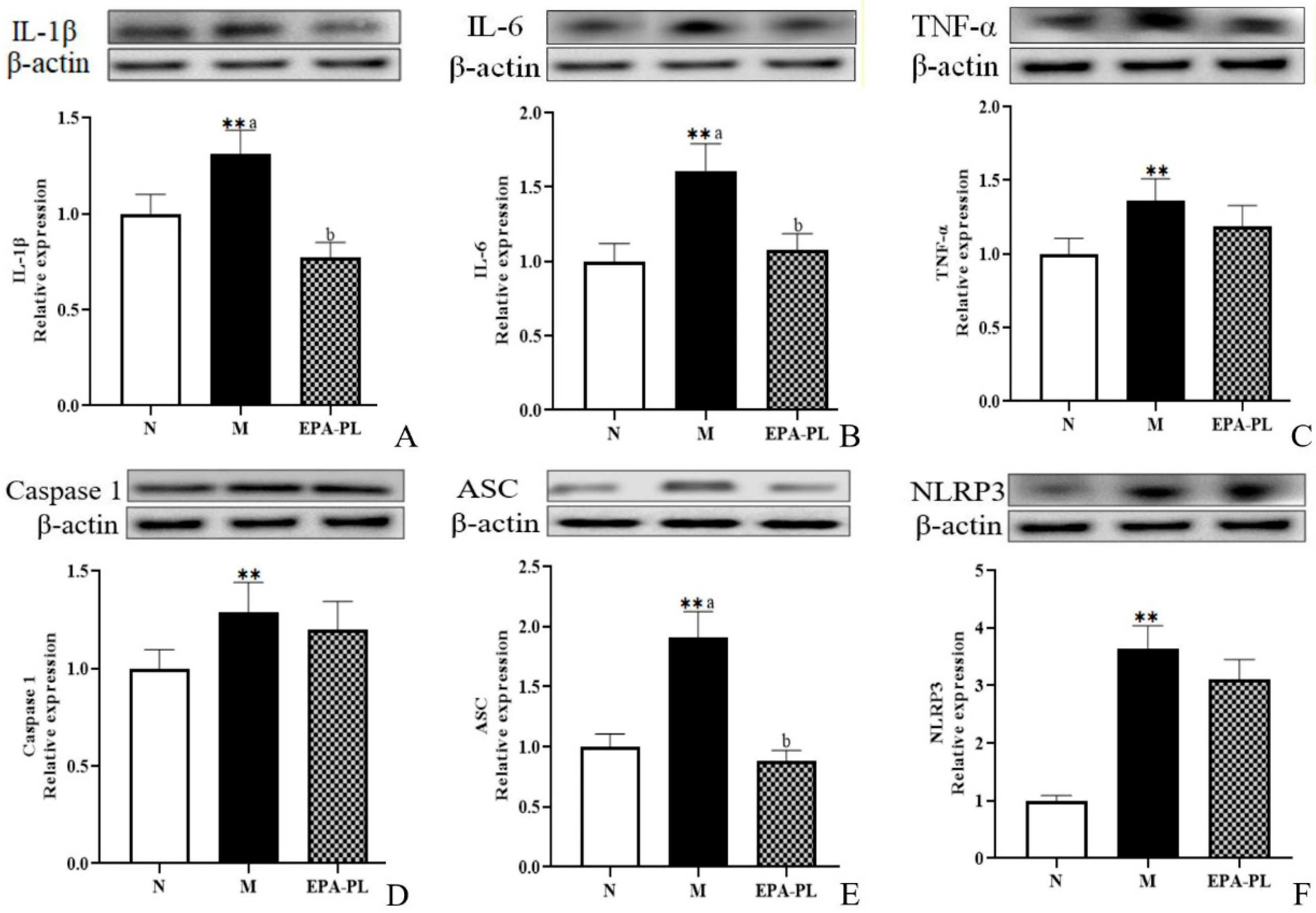
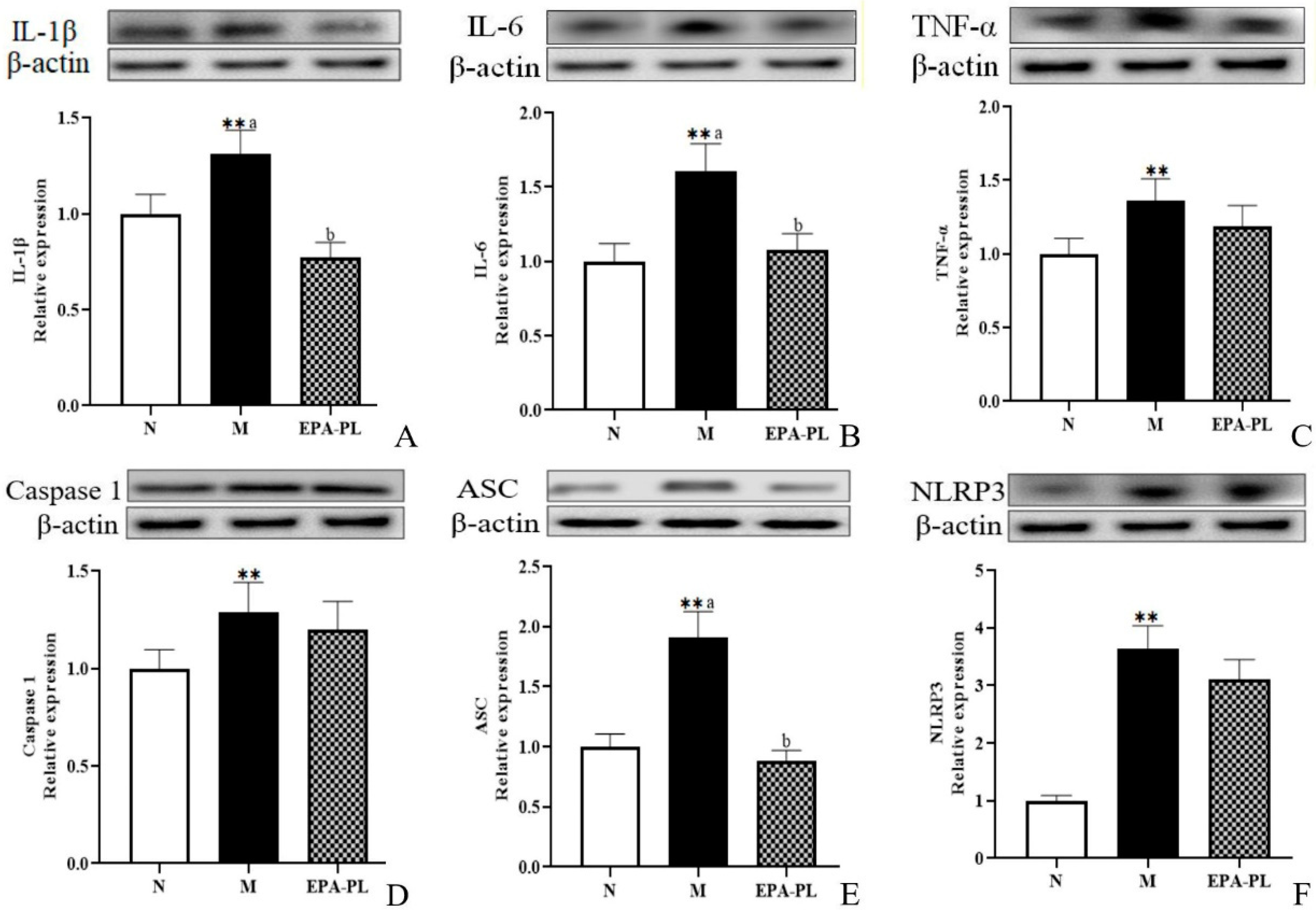
2.7. The Effects of EPA-PL on NF-κB-Meditated Inflammation Signaling Pathway in SHRs
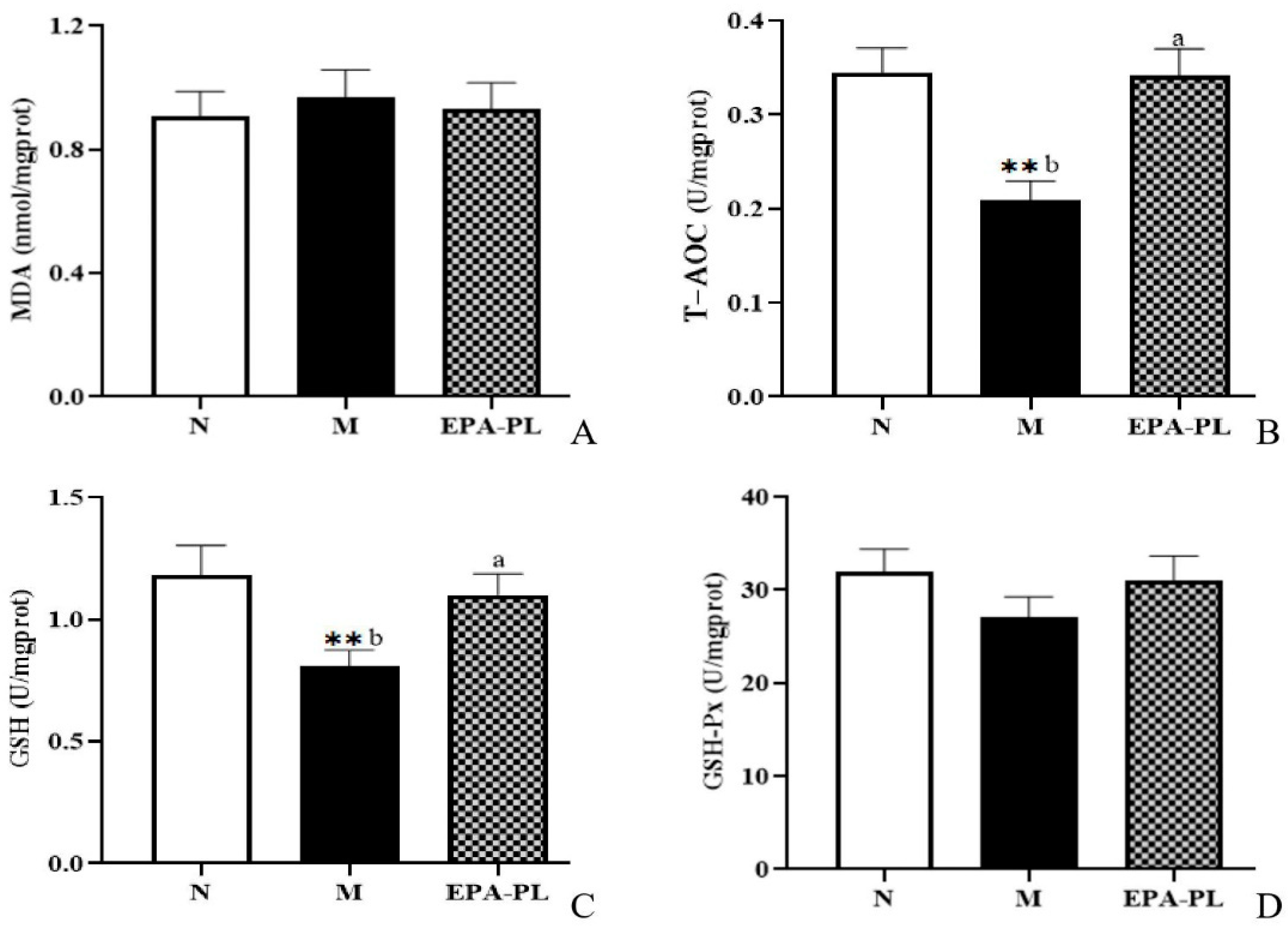
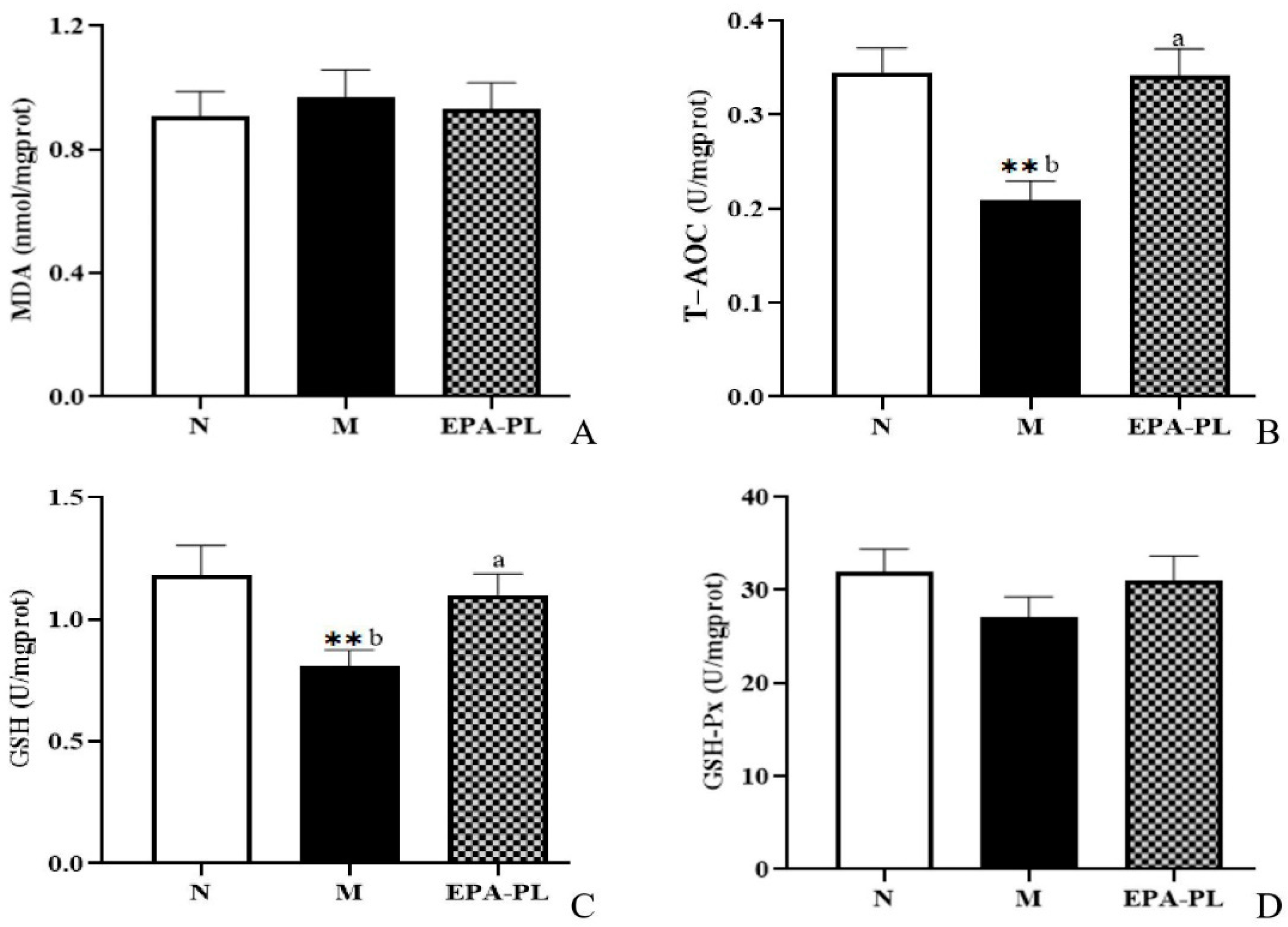
2.8. The Effects of EPA-PL on Oxidative Stress in SHRs
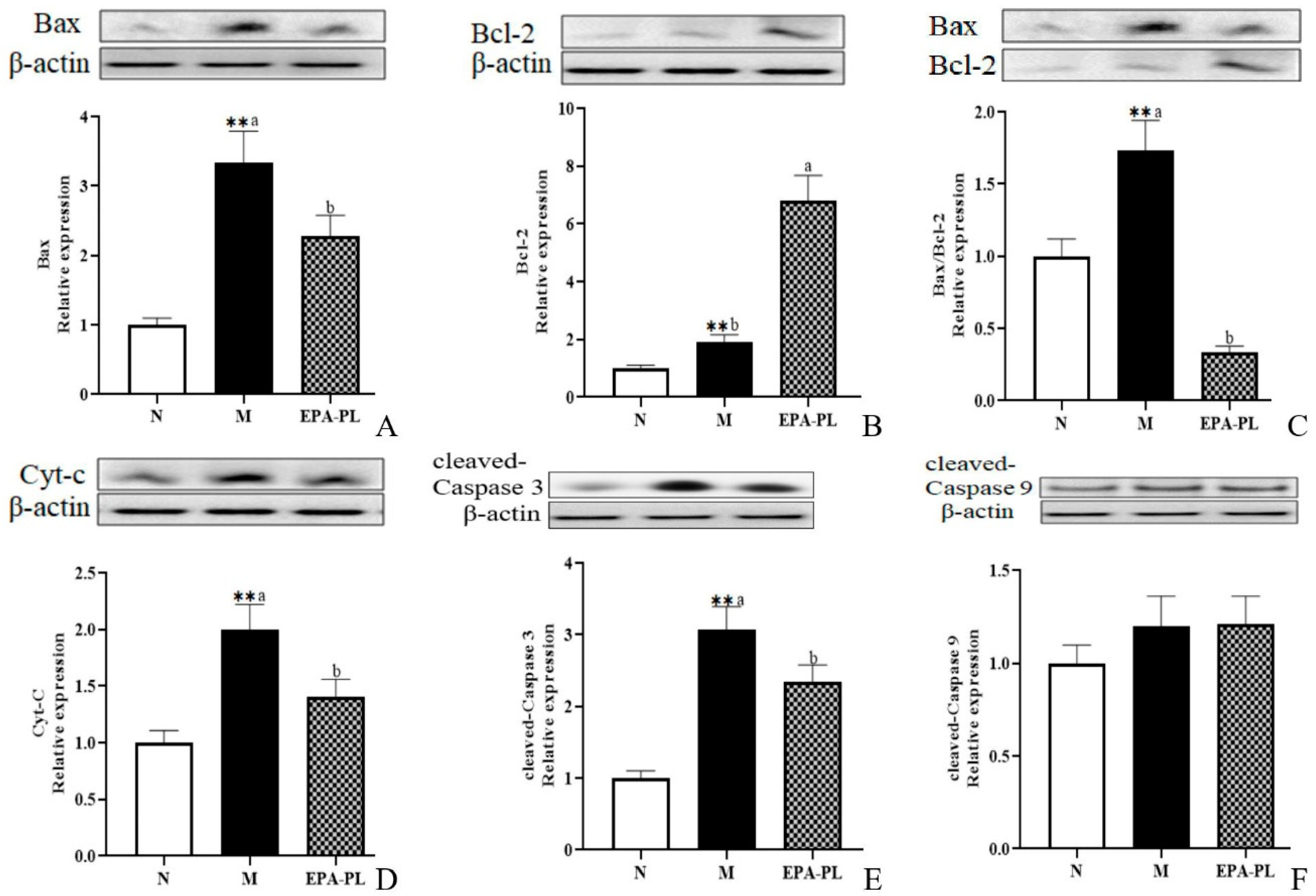
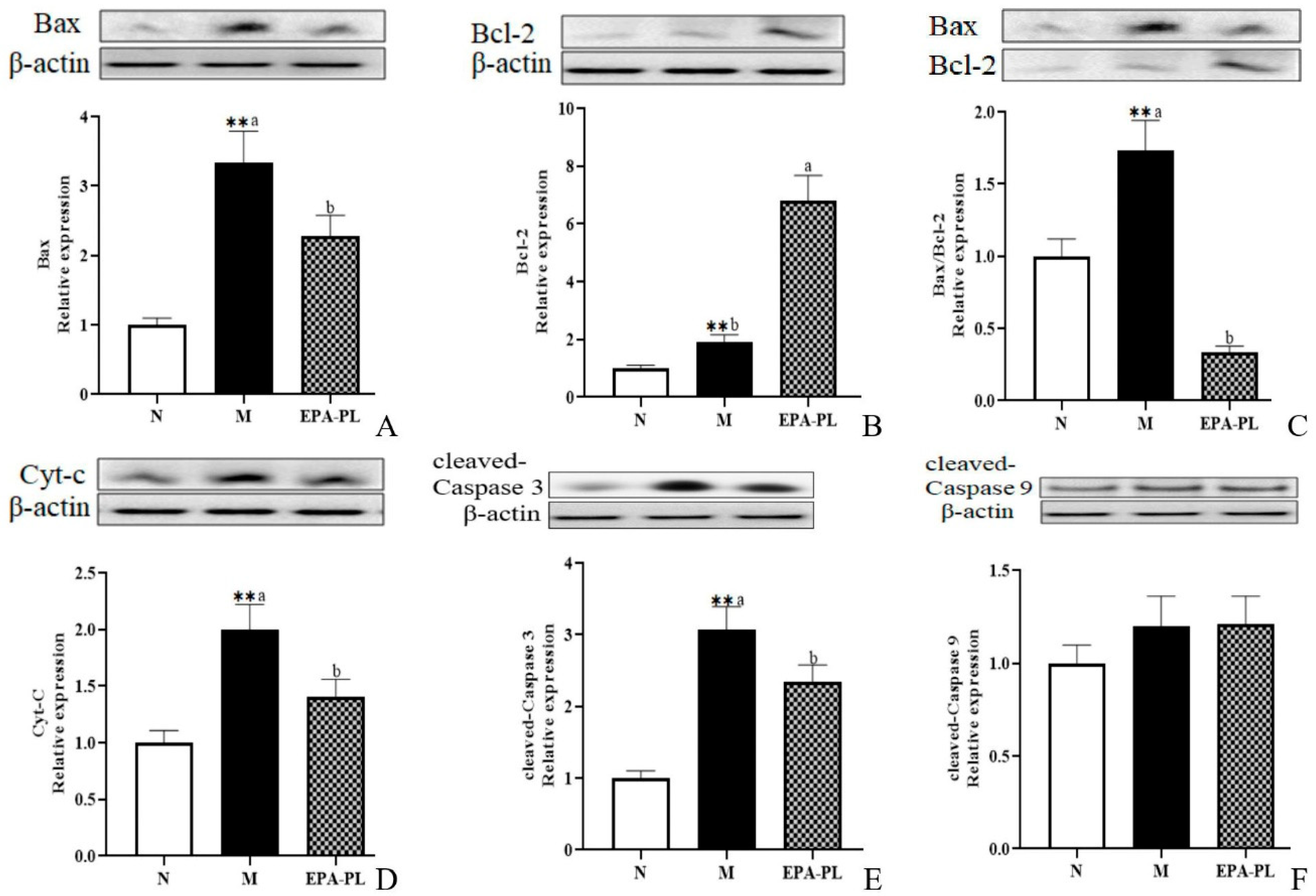
2.9. The Effects of EPA-PL on Mitochondria-Mediated Apoptosis Signaling Pathway in SHRs
3. Materials and Methods
3.1. Materials
3.2. Preparation of EPA-PL
3.3. Animal Experiment Design
3.4. Biochemical Analysis
3.5. Histological Analysis
3.6. Western Blotting Analysis
3.7. Statistical Analysis
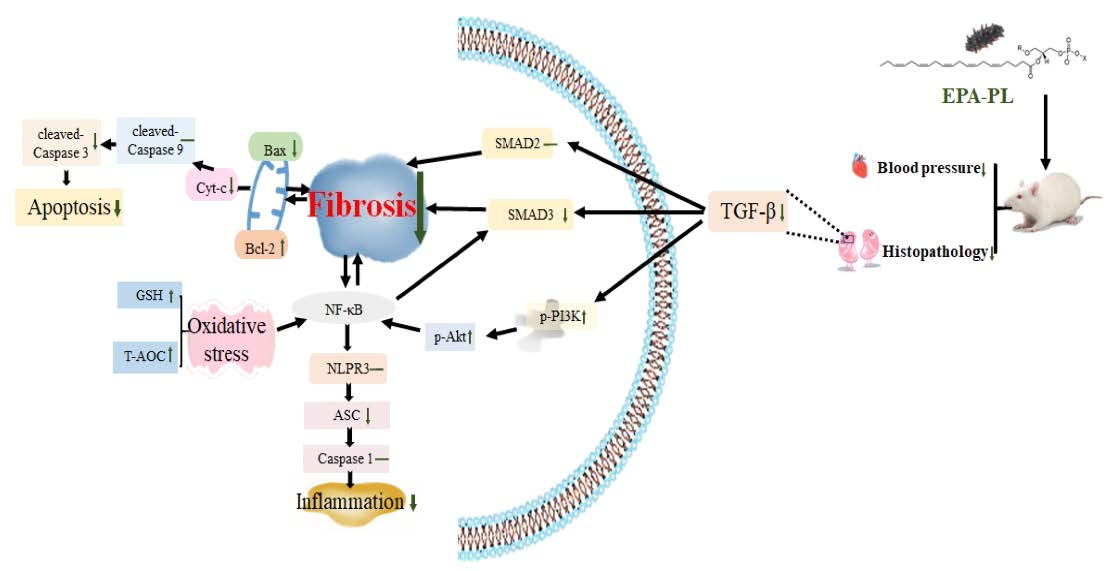
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tsioufis, C.; Thomopoulos, C. Combination drug treatment in hypertension. Pharmacol. Res. 2017, 125, 266–271. [Google Scholar] [CrossRef]
- Raine, A.E. Hypertension and the kidney. Br. Med. Bull. 1994, 50, 322–341. [Google Scholar] [CrossRef]
- Takenaka, T.; Inoue, T.; Miyazaki, T.; Kobori, H.; Nishiyama, A.; Ishii, N.; Hayashi, M.; Suzuki, H. Klotho Ameliorates Medullary Fibrosis and Pressure Natriuresis in Hypertensive Rat Kidneys. Hypertension 2018, 72, 1151–1159. [Google Scholar] [CrossRef]
- Worou, M.E.; Belmokhtar, K.; Bonnet, P.; Vourc’h, P.; Machet, M.C.; Khamis, G.; Eder, V. Hemin decreases cardiac oxidative stress and fibrosis in a rat model of systemic hypertension via PI3K/Akt signalling. Cardiovasc. Res. 2011, 91, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Gan, Z.; Huang, D.; Jiang, J.; Li, Y.; Li, H.; Ke, Y. Captopril alleviates hypertension-induced renal damage, inflammation, and NF-κB activation. Braz. J. Med. Biol. Res. 2018, 51, e7338. [Google Scholar] [CrossRef]
- Fantin, F.; Macchi, F.; Giani, A.; Bissoli, L. The Importance of Nutrition in Hypertension. Nutrients 2019, 11, 2542. [Google Scholar] [CrossRef] [Green Version]
- Ajeigbe, O.F.; Ademosun, A.O.; Oboh, G. Relieving the tension in hypertension: Food-drug interactions and anti-hypertensive mechanisms of food bioactive compounds. J. Food Biochem. 2021, 45, e13317. [Google Scholar] [CrossRef]
- Martínez-Martos, J.M.; Ramírez-Expósito, M.J. Dietary fat and hypertension: A novel approach through the proteolytic regulatory enzymes of the renin-angiotensin-system. Cardiovasc. Hematol. Agents Med. Chem. 2006, 4, 263–276. [Google Scholar] [CrossRef]
- Abraham, C.; Speth, R.C. The relationship between omega-3 fatty acids and blood pressure. J. Pharm. Health Serv. Res. 2018, 10, 161–168. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Guo, X.F.; Li, K.L.; Li, J.M.; Li, D. Effects of EPA and DHA on blood pressure and inflammatory factors: A meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2019, 59, 3380–3393. [Google Scholar] [CrossRef]
- Zhang, T.T.; Xu, J.; Wang, Y.M.; Xue, C.H. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids. Prog. Lipid Res. 2019, 75, 100997. [Google Scholar] [CrossRef]
- Ding, L.; Wang, D.; Zhou, M.; Du, L.; Xu, J.; Xue, C.; Wang, Y. Comparative Study of EPA-enriched Phosphatidylcholine and EPA-enriched Phosphatidylserine on Lipid Metabolism in Mice. J. Oleo Sci. 2016, 65, 593. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ding, L.; Shi, H.; Wang, C.; Xue, C.; Zhang, T.; Wang, Y. The Different Protective Effects of Phospholipids Against Obesity-Induced Renal Injury Mainly Associate with Fatty Acid Composition. Eur. J. Lipid Sci. Technol. 2021, 123, 2100011. [Google Scholar] [CrossRef]
- Shi, H.H.; Wang, C.C.; Guo, Y.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. DHA-PC protects kidneys against cisplatin-induced toxicity and its underlying mechanisms in mice. Food Funct. 2019, 10, 1571–1581. [Google Scholar] [CrossRef]
- Deckers, K.; Schievink, S.H.J.; Rodriquez, M.M.F.; van Oostenbrugge, R.J.; van Boxtel, M.P.J.; Verhey, F.R.J.; Köhler, S. Coronary heart disease and risk for cognitive impairment or dementia: Systematic review and meta-analysis. PLoS ONE 2017, 12, e0184244. [Google Scholar] [CrossRef]
- Yang, B.; Shi, L.; Wang, A.M.; Shi, M.Q.; Li, Z.H.; Zhao, F.; Guo, X.J.; Li, D. Lowering Effects of n-3 Fatty Acid Supplements on Blood Pressure by Reducing Plasma Angiotensin II in Inner Mongolia Hypertensive Patients: A Double-Blind Randomized Controlled Trial. J. Agric. Food Chem. 2019, 67, 184–192. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, W.; Chen, Q.; Cao, Q.; Di, W.; Lan, R.; Chen, Z.; Bai, J.; Han, Z.; Xu, W. Inhibition of RAGE by FPS-ZM1 alleviates renal injury in spontaneously hypertensive rats. Eur. J. Pharmacol. 2020, 882, 173228. [Google Scholar] [CrossRef]
- Almeida, L.F.; Tofteng, S.S.; Madsen, K.; Jensen, B.L. Role of the renin-angiotensin system in kidney development and programming of adult blood pressure. Clin. Sci. 2020, 134, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Ulu, A.; Stephen Lee, K.S.; Miyabe, C.; Yang, J.; Hammock, B.G.; Dong, H.; Hammock, B.D. An omega-3 epoxide of docosahexaenoic acid lowers blood pressure in angiotensin-II-dependent hypertension. J. Cardiovasc. Pharmacol. 2014, 64, 87–99. [Google Scholar] [CrossRef]
- Ajami, M.; Davoodi, S.H.; Habibey, R.; Namazi, N.; Soleimani, M.; Pazoki-Toroudi, H. Effect of DHA plus EPA on oxidative stress and apoptosis induced by ischemia-reperfusion in rat kidneys. Fundam. Clin. Pharmacol. 2013, 27, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.H.; Wang, C.C.; Ding, L.; Mao, X.Z.; Xue, C.H.; Yanagita, T.; Zhang, T.T.; Wang, Y.M. Comparative evaluation of phosphatidylcholine and phosphatidylserine with different fatty acids on nephrotoxicity in vancomycin-induced mice. Biosci. Biotechnol. Biochem. 2021, 85, 1873–1884. [Google Scholar] [CrossRef]
- Hultström, M. Development of structural kidney damage in spontaneously hypertensive rats. J. Hypertens. 2012, 30, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zou, J.; Zhang, T.; Che, H.; Gao, X.; Wang, C.; Wang, Y.; Xue, C. Protective Effects of DHA-PC against Vancomycin-Induced Nephrotoxicity through the Inhibition of Oxidative Stress and Apoptosis in BALB/c Mice. J. Agric. Food Chem. 2018, 66, 475–484. [Google Scholar] [CrossRef]
- Diaz Encarnacion, M.M.; Warner, G.M.; Gray, C.E.; Cheng, J.; Keryakos, H.K.; Nath, K.A.; Grande, J.P. Signaling pathways modulated by fish oil in salt-sensitive hypertension. Am. J. Physiol.-Ren. Physiol. 2008, 294, F1323–F1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elblehi, S.S.; Hafez, M.H.; El-Sayed, Y.S. L-α-Phosphatidylcholine attenuates mercury-induced hepato-renal damage through suppressing oxidative stress and inflammation. Environ. Sci. Pollut. Res. Int. 2019, 26, 9333–9342. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Chung, Y.P.; Liu, C.H.; Huang, K.T.; Guan, S.S.; Chiang, C.K.; Wu, C.T.; Liu, S.H. Withaferin A protects against endoplasmic reticulum stress-associated apoptosis, inflammation, and fibrosis in the kidney of a mouse model of unilateral ureteral obstruction. Phytomedicine Int. J. Phytother. Phytopharm. 2020, 79, 153352. [Google Scholar] [CrossRef]
- Giribabu, N.; Karim, K.; Kilari, E.K.; Salleh, N. Phyllanthus niruri leaves aqueous extract improves kidney functions, ameliorates kidney oxidative stress, inflammation, fibrosis and apoptosis and enhances kidney cell proliferation in adult male rats with diabetes mellitus. J. Ethnopharmacol. 2017, 205, 123–137. [Google Scholar] [CrossRef]
- Wei, Z.; Cao, J.; Zhang, X.; Yin, D.; Xu, D.; Lu, G. EPA attenuates epithelial-mesenchymal transition and fibrosis through the TGF-β1/Smad3/ILK pathway in renal tubular epithelial HK-2 cells by up-regulating miR-541. Int. J. Clin. Exp. Pathol. 2019, 12, 2516–2525. [Google Scholar]
- Cao, W.; Wang, C.; Chin, Y.; Chen, X.; Gao, Y.; Yuan, S.; Xue, C.; Wang, Y.; Tang, Q. DHA-phospholipids (DHA-PL) and EPA-phospholipids (EPA-PL) prevent intestinal dysfunction induced by chronic stress. Food Funct. 2019, 10, 277–288. [Google Scholar] [CrossRef]
- Pathil, A.; Mueller, J.; Ludwig, J.M.; Wang, J.; Warth, A.; Chamulitrat, W.; Stremmel, W. Ursodeoxycholyl lysophosphatidylethanolamide attenuates hepatofibrogenesis by impairment of TGF-β1/Smad2/3 signalling. Br. J. Pharmacol. 2014, 171, 5113–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, X.J.; Hou, J.G.; Jiang, S.; Liu, Z.; Tang, S.; Liu, X.X.; Wang, Y.P.; Chen, C.; Wang, Z.; Li, W. Maltol Mitigates Thioacetamide-induced Liver Fibrosis through TGF-β1-mediated Activation of PI3K/Akt Signaling Pathway. J. Agric. Food Chem. 2019, 67, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gao, X.; Li, K.; Liu, Y.; Hettiarachichi, D.S.; Sunderland, B.; Li, D. Sandalwood seed oil ameliorates hepatic insulin resistance by regulating the JNK/NF-κB inflammatory and PI3K/AKT insulin signaling pathways. Food Funct. 2021, 12, 2312–2322. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.P.; Fu, W.; Liu, J. Omega-3 PUFA Attenuates LPS-Induced Neuro-Injury of Neonatal Rats through the PI3K/AKT Pathway. Neuroscience 2019, 414, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yu, J.; Song, J.; Wang, S.; Cao, T.; Liu, Z.; Gao, X.; Wei, Y. The antihypertensive effect and mechanisms of bioactive peptides from Ruditapes philippinarum fermented with Bacillus natto in spontaneously hypertensive rats. J. Funct. Foods 2021, 79, 104411. [Google Scholar] [CrossRef]
- Javkhedkar, A.A.; Quiroz, Y.; Rodriguez-Iturbe, B.; Vaziri, N.D.; Lokhandwala, M.F.; Banday, A.A. Resveratrol restored Nrf2 function, reduced renal inflammation, and mitigated hypertension in spontaneously hypertensive rats. Am. J. Physiology. Regul. Integr. Comp. Physiol. 2015, 308, R840–R846. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Zhang, Z.; Huang, Y.; Sun, H.; Qiu, L. Vaccarin alleviates hypertension and nephropathy in renovascular hypertensive rats. Exp. Ther. Med. 2018, 15, 924–932. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, N.S.; Tostes, R.C.; Paradis, P.; Schiffrin, E.L. Aldosterone, Inflammation, Immune System, and Hypertension. Am. J. Hypertens. 2021, 34, 15–27. [Google Scholar] [CrossRef]
- Zhang, Z.; Xue, Z.; Yang, H.; Zhao, F.; Liu, C.; Chen, J.; Lu, S.; Zou, Z.; Zhou, Y.; Zhang, X. Differential effects of EPA and DHA on DSS-induced colitis in mice and possible mechanisms involved. Food Funct. 2021, 12, 1803–1817. [Google Scholar] [CrossRef]
- Nowacki, D.; Martynowicz, H.; Skoczynska, A.; Wojakowska, A.; Turczyn, B.; Bobak, L.; Trziszka, T.; Szuba, A. Lecithin derived from omega-3 PUFA fortified eggs decreases blood pressure in spontaneously hypertensive rats. Sci. Rep. 2017, 7, 12373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Y.; Ding, L.; Shi, H.H.; Xu, J.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Eicosapentaenoic acid in the form of phospholipids exerts superior anti-atherosclerosis effects to its triglyceride form in ApoE(-/-) mice. Food Funct. 2019, 10, 4177–4188. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Li, Q.; Zhang, T.; Ding, L.; Zhang, L.; Shi, H.H.; Yanagita, T.; Xue, C.; Chang, Y.; Wang, Y. A comparative study of EPA-enriched ethanolamine plasmalogen and EPA-enriched phosphatidylethanolamine on Aβ42 induced cognitive deficiency in a rat model of Alzheimer’s disease. Food Funct. 2018, 9, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, A.M.; Abdel-Dayem, M.A.; Abdel-Aziz, A.; El-Azab, M.F.; Said, S.A. Eicosapentaenoic acid ablates valproate-induced liver oxidative stress and cellular derangement without altering its clearance rate: Dynamic synergy and therapeutic utility. Biochim. Biophys. Acta, Mol. Cell Biol. Lipids. 2011, 1811, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.C.; Shi, L.; Jin, Z.; Hu, M.; Huang, H.; Yan, T.; Zhang, K.R. Swimming Exercise Ameliorates Hypertension-Induced Kidney Dysfunction via Alleviating Renal Interstitial Fibrosis and Apoptosis. Kidney Blood Press. Res. 2021, 46, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Chen, X.; Zhong, G.; Chen, Q.; Hu, C. Mitofusin 2 Downregulation Triggers Pulmonary Artery Smooth Muscle Cell Proliferation and Apoptosis Imbalance in Rats With Hypoxic Pulmonary Hypertension Via the PI3K/Akt and Mitochondrial Apoptosis Pathways. J. Cardiovasc. Pharmacol. 2016, 67, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A. Pathophysiological mechanisms of mineralocorticoid receptor-dependent cardiovascular and chronic kidney disease. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2019, 42, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wang, J.; Shao, W.; Wang, J.; Yao, L.; Li, Z.; Ohno, S. Pathogenesis and Damage Targets of Hypertensive Kidney Injury. J. Transl. Intern. Med. 2020, 8, 205–209. [Google Scholar] [CrossRef]
- Fukui, M.; Kang, K.S.; Okada, K.; Zhu, B.T. EPA, an omega-3 fatty acid, induces apoptosis in human pancreatic cancer cells: Role of ROS accumulation, caspase-8 activation, and autophagy induction. J. Cell. Biochem. 2013, 114, 192–203. [Google Scholar] [CrossRef]
- Sebe, M.; Tsutsumi, R.; Yamaguchi, S.; Horikawa, Y.T.; Harada, N.; Oyama, T.; Kakuta, N.; Tanaka, K.; Tsutsumi, Y.M.; Nakaya, Y.; et al. The synergystic effects of omega-3 fatty acids against 5-fluorouracil-induced mucosal impairment in mice. BMC Nutr. 2016, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Taneda, S.; Honda, K.; Tomidokoro, K.; Uto, K.; Nitta, K.; Oda, H. Eicosapentaenoic acid restores diabetic tubular injury through regulating oxidative stress and mitochondrial apoptosis. Am. J. Physiol. Ren. Physiol. 2010, 299, F1451–F1461. [Google Scholar] [CrossRef] [Green Version]
- Martínez-González, M.A.; Gea, A.; Ruiz-Canela, M. The Mediterranean Diet and Cardiovascular Health. Circ. Res. 2019, 124, 779–798. [Google Scholar] [CrossRef] [PubMed]
- Akhlaghi, M. Dietary Approaches to Stop Hypertension (DASH): Potential mechanisms of action against risk factors of the metabolic syndrome. Nutr. Res. Rev. 2020, 33, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bercea, C.I.; Cottrell, G.S.; Tamagnini, F.; McNeish, A.J. Omega-3 polyunsaturated fatty acids and hypertension: A review of vasodilatory mechanisms of docosahexaenoic acid and eicosapentaenoic acid. Br. J. Pharmacol. 2021, 178, 860–877. [Google Scholar] [CrossRef] [PubMed]
- Vara-Messler, M.; Mukdsi, J.H.; Osieki, N.I.; Benizio, E.; Repossi, G.M.; Ajayi, E.I.O.; García, N.H. Eicosapentaenoic acid prevents salt sensitivity in diabetic rats and decreases oxidative stress. Nutrition 2020, 72, 110644. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | EPA-PL (g/100 g) |
|---|---|
| C14:0 | 3.41 ± 0.23 |
| C16:0 | 3.63 ± 0.29 |
| C16:1 n-9 | 6.69 ± 0.75 |
| C18:0 | 5.32 ± 0.48 |
| C18:1 n-9 | 5.45 ± 0.36 |
| C20:1 n-9 | 6.93 ± 0.59 |
| C20:3 n-6 | 1.95 ± 0.12 |
| C20:3 n-3 | 5.75 ± 0.41 |
| C20:5 n-3 (EPA) | 38.61 ± 2.91 |
| C22:6 n-3 (DHA) | 4.21 ± 0.38 |
| Ingredients (g/kg) | N | M | EPA-PL |
|---|---|---|---|
| Potato starch | 349.5 | 349.5 | 349.5 |
| Casein | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 |
| Soy oil | 50 | 50 | 46 |
| Lard | 200 | 200 | 184 |
| Cellulose | 50 | 50 | 50 |
| Mineral–salt mix | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 |
| l-cystine | 3 | 3 | 3 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 |
| EPA-PL | - | - | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.-H.; Zhang, L.-Y.; Chen, L.-P.; Yang, J.-Y.; Wang, C.-C.; Xue, C.-H.; Wang, Y.-M.; Zhang, T.-T. EPA-Enriched Phospholipids Alleviate Renal Interstitial Fibrosis in Spontaneously Hypertensive Rats by Regulating TGF-β Signaling Pathways. Mar. Drugs 2022, 20, 152. https://doi.org/10.3390/md20020152
Shi H-H, Zhang L-Y, Chen L-P, Yang J-Y, Wang C-C, Xue C-H, Wang Y-M, Zhang T-T. EPA-Enriched Phospholipids Alleviate Renal Interstitial Fibrosis in Spontaneously Hypertensive Rats by Regulating TGF-β Signaling Pathways. Marine Drugs. 2022; 20(2):152. https://doi.org/10.3390/md20020152
Chicago/Turabian StyleShi, Hao-Hao, Ling-Yu Zhang, Li-Pin Chen, Jin-Yue Yang, Cheng-Cheng Wang, Chang-Hu Xue, Yu-Ming Wang, and Tian-Tian Zhang. 2022. "EPA-Enriched Phospholipids Alleviate Renal Interstitial Fibrosis in Spontaneously Hypertensive Rats by Regulating TGF-β Signaling Pathways" Marine Drugs 20, no. 2: 152. https://doi.org/10.3390/md20020152