
Marine versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Chemo-Diversity of (exo)Polysaccharides
3. Polysaccharides as Skincare Cosmetic Ingredients
4. Regulation and Safety of Cosmetic Products
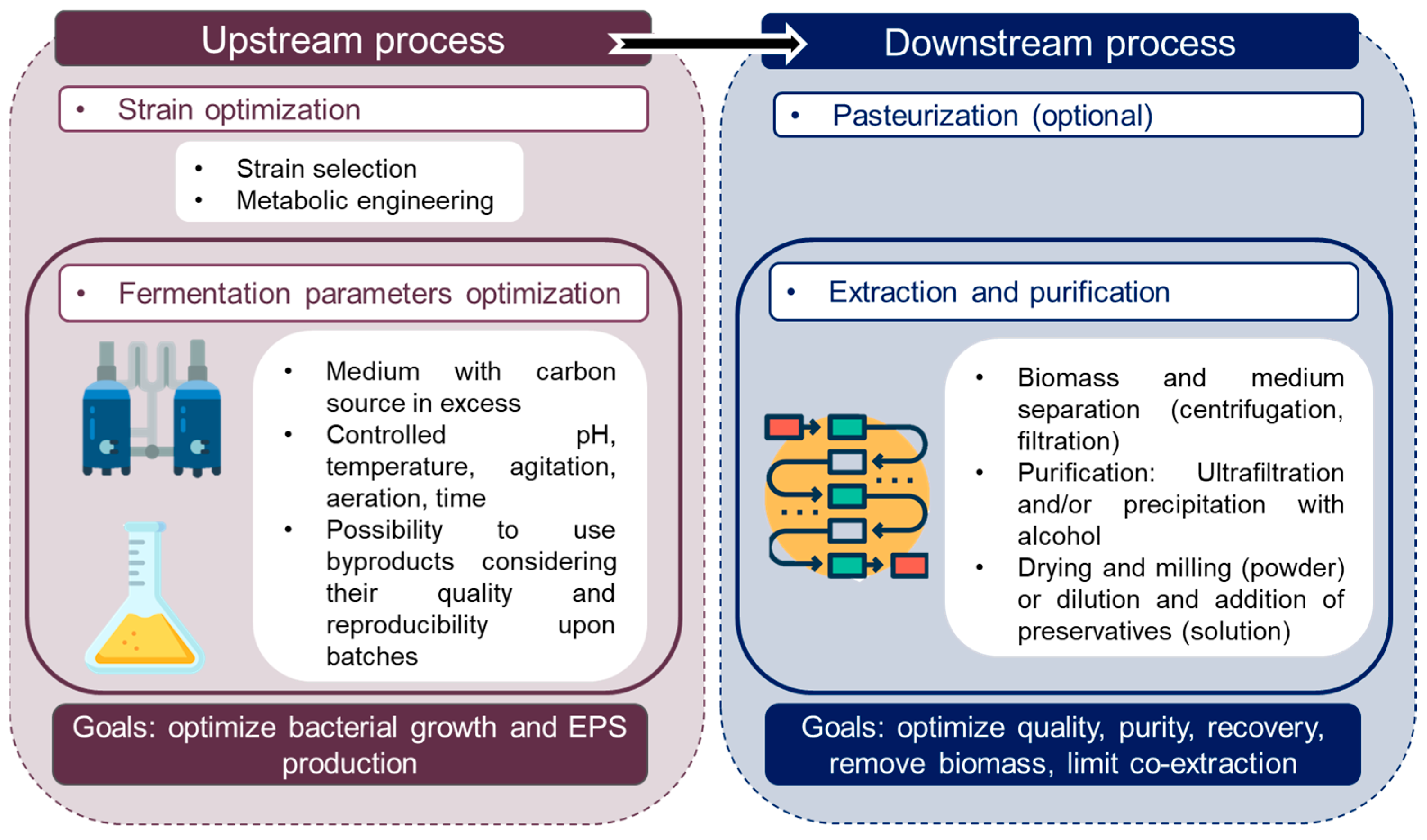
5. Production and Purification Processes for Bacterial EPS
5.1. Upstream Process
5.1.1. Impact of Culture Conditions on EPS Production
5.1.2. Optimization of EPS Production
5.1.3. EPS Production at Large Scale
5.2. Downstream Process

5.3. Bacterial EPS Production for Cosmetic Ingredients Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Medium | Carbon Source | Temperature, pH, Time | Extraction & Purification | Refs. |
|---|---|---|---|---|---|
| X. campestris | Soy proteins 2.5 g/L, calcium carbonate 0.1 g/L | Corn starch 38 g/L Glc, Suc 20–40 g/L | 28–30 °C; pH 6.0–7.5 60–100 h | Broth pasteurization at 90–120 °C for 20 min, cell removal, purification by ethanol precipitation, drying, milling. | [66,67,68] |
| C. marina CNCM I-4353 | Peptone 4 g/L, yeast extract 1 g/L, salts 27 g/L | Glc, Suc or Fru 30 g/L | 25–30 °C | Cell removal by centrifugation and filtration, extraction by ultrafiltration using 300 kDa membranes. | [64] |
| H. anticariensis LMG P-27891 | Yeast extract, malt extract, peptone 0.5–10 g/L, salts | Glc 25 g/L | 32 °C; pH 7.0; 24–72 h | Cell removal by centrifugation and filtration, ultrafiltration, dialysis using 10 kDa PES membranes. Purification by ethanol, acetone or isopropanol precipitation. | [62] |
| H. eurihalina LMG P-28571 | Yeast extract 3 g/L, malt extract 3 g/L, pea peptone 5 g/L, salt solution | Glc 10 g/L | 32 °C; pH 7.0; 40 h | Cell removal by centrifugation and filtration, ultrafiltration 10 kDa cut-off PES membranes or dialysis with 10 kDa cut-off membrane. Purification by ethanol, acetone or isopropanol precipitation. | [4] |
| Pseudoalteromonas sp. CNCM I-4150 | Medium with salts | Glc 20 g/L | 29 °C; pH 7.5; 72 h | Cell removal by centrifugation and filtration, purification with distilled water by ultrafiltration on 100 kDa cut-off PES membranes. | [69] |
| V. alginolyticus CNCM I-5035 | Zobell medium: sea salts 30 g/L, Yeast extract 1 g/L, Peptone 4 g/L | Glc 30 g/L | 25 °C; pH 7.2; 72 h | Cell removal by centrifugation and filtration, ultrafiltration using 100 kDa cut-off membranes. | [65] |
| Vibrio sp. CNCM I-4239; Vibrio sp. CNCM I-4277 | Zobell medium with salts | Glc 20 g/L | 29 °C; pH 7.5; 72 h | Cell removal by centrifugation and filtration, purification with distilled water by ultrafiltration on 100 kDa cut-off PES membranes. | [70,71] |
6. EPS for Cosmetic Applications: Non-Marine versus Marine EPS
6.1. Non-Marine Bacterial EPS in Cosmetics

6.2. Marine Bacterial EPS in Cosmetics
6.3. EPS-Producing Extremophilic Bacteria
| Bacterial Strain | Type | Environnent | In-Lab Production Conditions and Yield | EPS Composition | Mw (g/mol) | Refs. |
|---|---|---|---|---|---|---|
| Bacillus licheniformis T14 | Thermophilic Halophilic | Marine hydrothermal vent | Bioreactor: 50 °C; pH 8.0; 48 h; NaCl 50 g/L Yield: 366 mg/L | Fru, Fuc, Glc, GalN, Man | 1 × 106 | [169,170] |
| Bacillus licheniformis B3-15 | Thermophilic | Shallow hydrothermal vent | Shake flask: 45 °C Yield: 165 mg/L | Man, Glc | 6 × 105 | [171] |
| Geobacillus thermodenitrificans sp. B3-72 | Thermophilic | Shallow hydrothermal vent | Shake flask: 65 °C; 72 h Yield: 70 mg/L | Man, Glc | 4 × 105 | [172] |
| Geobacillus sp. 4004 | Thermophilic | Marine hot spring | Bioreactor: 60 °C; pH 7.0; 28 h Yield: 90 mg/L | Man, Glc, Gal, GlcN, Ara | 1 × 106 | [173] |
| Rhodothermus marinus DSM 4252 and MAT 493 | Thermophilic | Shallow marine hot spring | Shake flask: 65 °C; 48 h Yield: N.I | Ara, Xyl Acetate, sulfate | 8 × 104 | [174] |
| Strain 4001 | Thermophilic | Marine hot spring | Shake flask: 65 °C; 48 h Yield: 60 mg/L | Man, Glc, Gal, ManN | 4 × 105 | [175] |
| Colwellia psychrerythraea 34H | Psychrophilic | Arctic sediments | Shake flask: 4 °C Yield: N.I | GalA, QuiNAc Alanine decoration | N.I | [176] |
| Colwellia sp. GW185 | Psychrophilic | Antarctic sponge | Shake flask: 15 °C; 168 h Yield: 183 mg/L | Glc, Man, Gal, GalN, GlcA, GalA | N.I | [177] |
| Marinobacter sp. W1-16 | Psychrophilic | Antarctic seawater | Shake flask: 15 °C; 192 h Yield: 139 mg/L | Glc, Man, Gal, GalN, GalA, GlcA Sulfate | 3 × 105 | [178] |
| Polaribacter sp. SM1127 | Psychrophilic | Arctic brown algae | Shake flask: 15 °C; 120 h 2110 mg/L | GlcNAc, Man, GlcA | 2 × 105 | [179] |
| Pseudoalteromonas sp. MER144 | Psychrophilic | Antarctic seawater | Shake flask: 4 °C; 336 h Yield: 318 mg/L | Glc, Man, GlcN, Ara, GlcA, GalA, Gal | 3 × 105 | [180] |
| Pseudoalteromonas sp. SM20310 | Psychrophilic | Arctic sea ice | Shake flask: 15 °C; 72 h Yield: 567 mg/L | Man, Glc, Gal, Rha | 2 × 106 | [181] |
| Pseudoalteromonas sp. SM9913 | Psychrophilic | Deep-sea sediment | Shake flask: 15 °C; 52 h Yield: 5250 mg/L | Glc, Ara | 4 × 104 | [182] |
| Pseudomonas sp. ID1 | Psychrophilic | Antarctic marine sediment | Shake flask: 11 °C; 120 h Yield: N.I | Glc, Gal, Fuc | 2 × 106 | [183] |
| Shewanella sp. CAL606 | Psychrophilic | Antarctic sponge | Shake flask: 4 °C; 240 h Yield: 329 mg/L | Glc, Gal, Man, GalN, GlcA, GalA | N.I | [177] |
| Winogradskyella sp. CAL396 | Psychrophilic | Antarctic sponge | Shake flask: 4 °C; 240 h Yield: 397 mg/L | Man, Ara, GalA, GlcA, Gal, Glc, GlcN | N.I | [177] |
| Winogradskyella sp. CAL384 | Psychrophilic | Antarctic sponge | Shake flask: 4 °C; 240 h Yield: 144 mg/L | Glc, Man, GalA, Ara, Gal, GlcN, GlcA | N.I | [177] |
6.4. Bioactivity Evaluation of Marine EPS
6.5. Structure-Function Relationship
7. Challenges and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holzwarth, G. Molecular Weight of Xanthan Polysaccharide. Carbohydr. Res. 1978, 66, 173–186. [Google Scholar] [CrossRef]
- Tanaka, T.; Oi, S.; Yamamoto, T. Synthesis of Levan by Levansucrase. Some Factors Affecting the Rate of Synthesis and Degree of Polymerization of Levan. J. Biochem. 1979, 85, 287–293. [Google Scholar] [CrossRef]
- Raguénès, G.; Christen, R.; Guezennec, J.; Pignet, P.; Barbier, G. Vibrio diabolicus sp. Nov., a New Polysaccharide-Secreting Organism Isolated from a Deep-Sea Hydrothermal Vent Polychaete Annelid, Alvinella pompejana. Int. J. Syst. Bacteriol. 1997, 47, 989–995. [Google Scholar] [CrossRef]
- Laporta Alcántara, O.; Almiñana Domènech, N.; Soley Astals, A.; Ferrer Montiel, A.V.; García Sanz, N. Cosmetic Composition Containing Halomonas Ferment Extract, and Use Thereof. European Patent EP3212291 B1, 22 August 2018. [Google Scholar]
- Kamerling, J.P.; Gerwig, G.J. Strategies for the Structural Analysis of Carbohydrates. In Comprehensive Glycoscience; Elsevier: Oxford, UK, 2007; pp. 1–68. ISBN 978-0-444-51967-2. [Google Scholar]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Letisse, F.; Chevallereau, P.; Simon, J.-L.; Lindley, N. The Influence of Metabolic Network Structures and Energy Requirements on Xanthan Gum Yields. J. Biotechnol. 2002, 99, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Landhuis, E. Glycobiology: Sweet Success. Nature 2017, 547, 127–129. [Google Scholar] [CrossRef]
- Sutherland, I.W. Bacterial Exopolysaccharides. Adv. Microb. Physiol. 1972, 8, 143–213. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Bacterial Surface Polysaccharides: Structure and Function. Int. Rev. Cytol. 1988, 113, 187–231. [Google Scholar] [CrossRef]
- Sutherland, I.W. Structure-Function Relationships in Microbial Exopolysaccharides. Biotechnol. Adv. 1994, 12, 393–448. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Sinquin, C.; Lebellenger, L.; Zykwinska, A.; Colliec-Jouault, S. Exopolysaccharides Produced by Marine Bacteria and Their Applications as Glycosaminoglycan-like Molecules. Front. Chem. 2014, 2, 85. [Google Scholar] [CrossRef]
- Moorhouse, R.; Walkinshaw, M.D.; Arnott, S. Xanthan Gum-Molecular Conformation and Interactions. In Extracellular Microbial Polysaccharides; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1977; Volume 45, pp. 7–90. ISBN 9780841203723. [Google Scholar]
- Haxaire, K.; Braccini, I.; Milas, M.; Rinaudo, M.; Pérez, S. Conformational Behavior of Hyaluronan in Relation to Its Physical Properties as Probed by Molecular Modeling. Glycobiology 2000, 10, 587–594. [Google Scholar] [CrossRef]
- Makshakova, O.; Zykwinska, A.; Cuenot, S.; Colliec-Jouault, S.; Perez, S. Three-Dimensional Structures, Dynamics and Calcium-Mediated Interactions of the Exopolysaccharide, Infernan, Produced by the Deep-Sea Hydrothermal Bacterium Alteromonas infernus. Carbohydr. Polym. 2022, 276, 118732. [Google Scholar] [CrossRef] [PubMed]
- Akoumany, K.; Zykwinska, A.; Sinquin, C.; Marchand, L.; Fanuel, M.; Ropartz, D.; Rogniaux, H.; Pipelier, M.; Delbarre-Ladrat, C.; Colliec-Jouault, S. Characterization of New Oligosaccharides Obtained by an Enzymatic Cleavage of the Exopolysaccharide Produced by the Deep-Sea Bacterium Alteromonas infernus Using Its Cell Extract. Molecules 2019, 24, 3441. [Google Scholar] [CrossRef]
- Mohd-Setapar, S.H.; John, C.P.; Mohd-Nasir, H.; Azim, M.M.; Ahmad, A.; Alshammari, M.B. Application of Nanotechnology Incorporated with Natural Ingredients in Natural Cosmetics. Cosmetics 2022, 9, 110. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Soto, M.L.; Pérez-Armada, L.; Domínguez, H. Cosmetics from Marine Sources. In Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1015–1042. ISBN 978-3-642-53971-8. [Google Scholar]
- Alves, A.; Sousa, E.; Kijjoa, A.; Pinto, M. Marine-Derived Compounds with Potential Use as Cosmeceuticals and Nutricosmetics. Molecules 2020, 25, 2536. [Google Scholar] [CrossRef]
- Courtois, A.; Guezennec, J. Obtention d’exopolysaccharides Bactériens et Applications En Cosmétique. Tech. L’ingénieur Formul. 2015. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, B. Skin Health Promoting Effects of Natural Polysaccharides and Their Potential Application in the Cosmetic Industry. Polysaccharides 2022, 3, 818–830. [Google Scholar] [CrossRef]
- Ferreira, M.; Matos, A.; Couras, A.; Marto, J.; Ribeiro, H. Overview of Cosmetic Regulatory Frameworks around the World. Cosmetics 2022, 9, 72. [Google Scholar] [CrossRef]
- CosIng Database. Available online: http://ec.europa.eu/growth/tools-databases/cosing/ (accessed on 9 May 2023).
- Rizzi, V.; Gubitosa, J.; Fini, P.; Cosma, P. Neurocosmetics in Skincare-The Fascinating World of Skin-Brain Connection: A Review to Explore Ingredients, Commercial Products for Skin Aging, and Cosmetic Regulation. Cosmetics 2021, 8, 66. [Google Scholar] [CrossRef]
- Regulation (EC) N°1223/2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32009R1223 (accessed on 27 March 2023).
- Desmeth, P. The Nagoya Protocol Applied to Microbial Genetic Resources. In Microbial Resources; Kurtböke, I., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 205–217. ISBN 978-0-12-804765-1. [Google Scholar]
- Humphries, F.; Rabone, M.; Jaspars, M. Traceability Approaches for Marine Genetic Resources under the Proposed Ocean (BBNJ) Treaty. Front. Mar. Sci. 2021, 8, 430. [Google Scholar] [CrossRef]
- Personal Care Products Council INCI Nomenclature (PCPC). Available online: https://www.personalcarecouncil.org/wp-content/uploads/2021/02/Conventions2021_v1.pdf (accessed on 11 July 2023).
- Freitas, F.; Alves, V.D.; Reis, M.A.M. Bacterial Polysaccharides: Production and Applications in Cosmetic Industry. In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 1–24. ISBN 978-3-319-03751-6. [Google Scholar]
- Almeida, A.; Sarmento, B.; Rodrigues, F. Insights on in vitro Models for Safety and Toxicity Assessment of Cosmetic Ingredients. Int. J. Pharm. 2017, 519, 178–185. [Google Scholar] [CrossRef]
- Pellevoisin, C.; Bouez, C.; Cotovio, J. Cosmetic Industry Requirements Regarding Skin Models for Cosmetic Testing; Marques, A.P., Pirraco, R.P., Cerqueira, M.T., Reis, R.L.B.T.-S.T.M., Eds.; Academic Press: Boston, MA, USA, 2018; pp. 3–37. ISBN 978-0-12-810545-0. [Google Scholar]
- Cornell, E.M.; Janetos, T.M.; Xu, S. Time for a Makeover-Cosmetics Regulation in the United States. J. Cosmet. Dermatol. 2019, 18, 2041–2047. [Google Scholar] [CrossRef] [PubMed]
- Fiume, M.M.; Heldreth, B.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C.; Slaga, T.J.; et al. Safety Assessment of Microbial Polysaccharide Gums as Used in Cosmetics. Int. J. Toxicol. 2016, 35, 5S–49S. [Google Scholar] [CrossRef] [PubMed]
- Becker, L.C.; Bergfeld, W.F.; Belsito, D.V.; Klaassen, C.D.; Marks, J.G.J.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; Andersen, F.A. Final Report of the Safety Assessment of Hyaluronic Acid, Potassium Hyaluronate, and Sodium Hyaluronate. Int. J. Toxicol. 2009, 28, 5–67. [Google Scholar] [CrossRef]
- Jeong, S.I.; Lee, S.E.; Yang, H.; Jin, Y.-H.; Park, C.-S.; Park, Y.S. Toxicologic Evaluation of Bacterial Synthesized Cellulose in Endothelial Cells and Animals. Mol. Cell. Toxicol. 2010, 6, 370–377. [Google Scholar] [CrossRef]
- Moreira, S.; Silva, N.B.; Almeida-Lima, J.; Rocha, H.A.O.; Medeiros, S.R.B.; Alves, C.J.; Gama, F.M. BC Nanofibres: In Vitro Study of Genotoxicity and Cell Proliferation. Toxicol. Lett. 2009, 189, 235–241. [Google Scholar] [CrossRef]
- García, A.; Fernández-Sandoval, M.T.; Morales-Guzmán, D.; Martínez-Morales, F.; Trejo-Hernández, M.R. Advances in Exopolysaccharide Production from Marine Bacteria. J. Chem. Technol. Biotechnol. 2022, 97, 2694–2705. [Google Scholar] [CrossRef]
- Liu, L.; Wang, M.; Du, G.; Chen, J. Enhanced Hyaluronic Acid Production of Streptococcus zooepidemicus by an Intermittent Alkaline-Stress Strategy. Lett. Appl. Microbiol. 2008, 46, 383–388. [Google Scholar] [CrossRef]
- Shu, C.-H.; Yang, S.-T. Effects of Temperature on Cell Growth and Xanthan Production in Batch Cultures of Xanthomonas campestris. Biotechnol. Bioeng. 1990, 35, 454–468. [Google Scholar] [CrossRef]
- Freitas, F.; Alves, V.D.; Reis, M.A. Advances in Bacterial Exopolysaccharides: From Production to Biotechnological Applications. Trends Biotechnol. 2011, 29, 388–398. [Google Scholar] [CrossRef]
- de Souza, A.M.; Sutherland, I.W. Exopolysaccharide and Storage Polymer Production in Enterobacter aerogenes Type 8 Strains. J. Appl. Bacteriol. 1994, 76, 463–468. [Google Scholar] [CrossRef]
- Sutherland, I.W. Biosynthesis of Microbial Exopolysaccharides. Adv. Microb. Physiol. 1982, 23, 79–150. [Google Scholar] [CrossRef] [PubMed]
- Delbarre-Ladrat, C.; Sinquin, C.; Marchand, L.; Bonnetot, S.; Zykwinska, A.; Verrez-Bagnis, V.; Colliec-Jouault, S. Influence of the Carbon and Nitrogen Sources on Diabolican Production by the Marine Vibrio diabolicus Strain CNCM I-1629. Polymers 2022, 14, 1994. [Google Scholar] [CrossRef]
- Shukla, A.; Mehta, K.; Parmar, J.; Pandya, J.; Saraf, M. Depicting the Exemplary Knowledge of Microbial Exopolysaccharides in a Nutshell. Eur. Polym. J. 2019, 119, 298–310. [Google Scholar] [CrossRef]
- Özcan, E.; Öner, E.T. Microbial of Extracellular Polysaccharide Production from Biomass Sources. In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–21. ISBN 978-3-319-03751-6. [Google Scholar]
- Barcelos, M.C.S.; Vespermann, K.A.C.; Pelissari, F.M.; Molina, G. Current Status of Biotechnological Production and Applications of Microbial Exopolysaccharides. Crit. Rev. Food Sci. Nutr. 2020, 60, 1475–1495. [Google Scholar] [CrossRef]
- Peña, C.; Trujillo-Roldán, M.A.; Galindo, E. Influence of Dissolved Oxygen Tension and Agitation Speed on Alginate Production and Its Molecular Weight in Cultures of Azotobacter vinelandii. Enzym. Microb. Technol. 2000, 27, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Amanullah, A.; Tuttiett, B.; Nienow, A.W. Agitator Speed and Dissolved Oxygen Effects in Xanthan Fermentations. Biotechnol. Bioeng. 1998, 57, 198–210. [Google Scholar] [CrossRef]
- Finore, I.; Di Donato, P.; Mastascusa, V.; Nicolaus, B.; Poli, A. Fermentation Technologies for the Optimization of Marine Microbial Exopolysaccharide Production. Mar. Drugs 2014, 12, 3005–3024. [Google Scholar] [CrossRef]
- Teworte, S.; Malcı, K.; Walls, L.E.; Halim, M.; Rios-Solis, L. Recent Advances in Fed-Batch Microscale Bioreactor Design. Biotechnol. Adv. 2022, 55, 107888. [Google Scholar] [CrossRef]
- Haby, B.; Hans, S.; Anane, E.; Sawatzki, A.; Krausch, N.; Neubauer, P.; Cruz Bournazou, M.N. Integrated Robotic Mini Bioreactor Platform for Automated, Parallel Microbial Cultivation with Online Data Handling and Process Control. SLAS Technol. 2019, 24, 569–582. [Google Scholar] [CrossRef]
- Von den Eichen, N.; Bromig, L.; Sidarava, V.; Marienberg, H.; Weuster-Botz, D. Automated Multi-Scale Cascade of Parallel Stirred-Tank Bioreactors for Fast Protein Expression Studies. J. Biotechnol. 2021, 332, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Rühmann, B.; Schmid, J.; Sieber, V. High Throughput Exopolysaccharide Screening Platform: From Strain Cultivation to Monosaccharide Composition and Carbohydrate Fingerprinting in One Day. Carbohydr. Polym. 2015, 122, 212–220. [Google Scholar] [CrossRef]
- Widner, B.; Behr, R.; Von Dollen, S.; Tang, M.; Heu, T.; Sloma, A.; Sternberg, D.; Deangelis, P.L.; Weigel, P.H.; Brown, S. Hyaluronic Acid Production in Bacillus subtilis. Appl. Environ. Microbiol. 2005, 71, 3747–3752. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Roldán, M.; Moreno, S.; Segura, D.; Galindo, E.; Espín, G. Alginate Production by an Azotobacter vinelandii Mutant Unable to Produce Alginate Lyase. Appl. Microbiol. Biotechnol. 2003, 60, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Luozhong, S.; Guo, Z.; Yu, H.; Stephanopoulos, G. Enhanced Biosynthesis of Hyaluronic Acid Using Engineered Corynebacterium glutamicum via Metabolic Pathway Regulation. Biotechnol. J. 2017, 12, 1700191. [Google Scholar] [CrossRef]
- Raguénès, G.; Cambon-Bonavita, M.A.; Lohier, J.F.; Boisset, C.; Guezennec, J. A Novel, Highly Viscous Polysaccharide Excreted by an Alteromonas Isolated from a Deep-Sea Hydrothermal Vent Shrimp. Curr. Microbiol. 2003, 46, 448–452. [Google Scholar] [CrossRef]
- Guézennec, J.; Moppert, X.; Raguénès, G.; Richert, L.; Costa, B.; Simon-Colin, C. Microbial Mats in French Polynesia and Their Biotechnological Applications. Process Biochem. 2011, 46, 16–22. [Google Scholar] [CrossRef]
- Raguénès, G.; Pignet, P.; Gauthier, G.; Peres, A.; Christen, R.; Rougeaux, H.; Barbier, G.; Guezennec, J. Description of a New Polymer-Secreting Bacterium from a Deep-Sea Hydrothermal Vent, Alteromonas macleodii subsp. fijiensis, and Preliminary Characterization of the Polymer. Appl. Environ. Microbiol. 1996, 62, 67–73. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Salas, M.L.; Sinquin, C.; Zykwinska, A.; Colliec-Jouault, S. Bioprospecting for Exopolysaccharides from Deep-Sea Hydrothermal Vent Bacteria: Relationship between Bacterial Diversity and Chemical Diversity. Microorganisms 2017, 5, 63. [Google Scholar] [CrossRef]
- Website Freepik. Available online: https://www.freepik.com/ (accessed on 18 October 2023).
- García Sanz, A.; Ferrer Montiel, A.V.; Soley Astals, A.; Almiñana Domènech, N. Expolysaccharide for the Treatment and/or Care of Skin, Culture Media and Compositions Thereof. European Patent EP3062759 B1, 14 August 2019. [Google Scholar]
- Wołosowska, S.; Synowiecki, J. Thermostable β-Glucosidase with a Broad Substrate Specifity Suitable for Processing of Lactose-Containing Products. Food Chem. 2004, 85, 181–187. [Google Scholar] [CrossRef]
- Gedouin, P.A.; Vallée, R.; Morvan, P.Y. Compound of Marine Origin and Its Use for Improving the Appearance of the Skin. France Patent FR3108847 B1, 22 July 2022. [Google Scholar]
- Drouillard, S.; Chambon, R.; Jeacomine, I.; Buon, L.; Boisset, C.; Courtois, A.; Thollas, B.; Morvan, P.-Y.; Vallée, R.; Helbert, W. Structure of the Polysaccharide Secreted by Vibrio alginolyticus CNCM I-5035 (Epidermist 4.0TM). Mar. Drugs 2020, 18, 509. [Google Scholar] [CrossRef]
- Clark, R.; Hayden, H. Xanthan Gum with Fast Hydration and High Viscosity. U.S. Patent US20120053339 A1, 1 March 2012. [Google Scholar]
- Sworn, G.; Kerdavid, E.; Chevallereau, P.; Fayos, J. Improved Xanthan Gum. U.S. Patent US9380803 B2, 5 July 2016. [Google Scholar]
- Funahashi, H.; Yoshida, T.; Taguchi, H. Effect of Glucose Concentrations on Xanthan Gum Production by Xanthomonas campestris. J. Ferment. Technol. 1987, 65, 603–606. [Google Scholar] [CrossRef]
- Courtois, A.; Thollas, B.; Delgado Gonzalez, R.; Cebrian, J.; Soley Astals, A. Exopolysaccharide for the Treatment and/or Care of the Skin, Mucous Membranes, Hair and/or Nails. European Patent EP2646115 B1, 8 November 2017. [Google Scholar]
- Delgado Gonzalez, R.; Soley Astals, A.; Courtois, A.; Thollas, B. Exopolysaccharide for the Treatment and/or Care of the Skin, Mucous Membranes and/or Nails. European Patent EP2827837 B1, 11 March 2020. [Google Scholar]
- Delgado Gonzalez, R.; Soley Astals, A.; Courtois, A.; Thollas, B. Exopolysaccharide for the Treatment and/or Care of the Skin, Mucous Membranes and/or Nails. European Patent EP2976060 B1, 26 October 2016. [Google Scholar]
- Website Mintel. Available online: https://clients.mintel.com/ (accessed on 18 October 2023).
- Margaritis, A.; Zajic, J.E. Mixing, Mass Transfer, and Scale-up of Polysaccharide Fermentations. Biotechnol. Bioeng. 1978, 20, 939–1001. [Google Scholar] [CrossRef]
- Jansson, P.; Kenne, L.; Lindberg, B. Structure of the Extracellular Polysaccharide from Xanthomonas campestris. Carbohydr. Res. 1975, 45, 275–282. [Google Scholar] [CrossRef]
- Melton, L.D.; Mindt, L.; Rees, D.A. Covalent Structure of the Extracellular Polysaccharide from Xanthomonas campestris: Evidence from Partial Hydrolysis Studies. Carbohydr. Res. 1976, 46, 245–257. [Google Scholar] [CrossRef]
- Capron, I.; Brigand, G.; Muller, G. About the Native and Renatured Conformation of Xanthan Exopolysaccharide. Polymer 1997, 38, 5289–5295. [Google Scholar] [CrossRef]
- Katzbauer, B. Properties and Applications of Xanthan Gum. Polym. Degrad. Stab. 1998, 59, 81–84. [Google Scholar] [CrossRef]
- Paoletti, S.; Cesàro, A.; Delben, F. Thermally Induced Conformational Transition of Xanthan Polyelectrolyte. Carbohydr. Res. 1983, 123, 173–178. [Google Scholar] [CrossRef]
- García-Ochoa, F.; Santos, V.E.; Casas, J.A.; Gómez, E. Xanthan Gum: Production, Recovery, and Properties. Biotechnol. Adv. 2000, 18, 549–579. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.; Lindberg, B.; Sandford, P.A. Structural Studies of Gellan Gum, an Extracellular Polysaccharide Elaborated by Pseudomonas elodea. Carbohydr. Res. 1983, 124, 135–139. [Google Scholar] [CrossRef]
- Tang, J.; Tung, M.A.; Zeng, Y. Compression Strength and Deformation of Gellan Gels Formed with Mono- and Divalent Cations. Carbohydr. Polym. 1996, 29, 11–16. [Google Scholar] [CrossRef]
- Akkineni, A.R.; Sen Elci, B.; Lode, A.; Gelinsky, M. Addition of High Acyl Gellan Gum to Low Acyl Gellan Gum Enables the Blends 3D Bioprintable. Gels 2022, 8, 199. [Google Scholar] [CrossRef]
- Meyer, K.; Palmer, J.W. The Polysaccharide of the Vitreaous Humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar] [CrossRef]
- Bogdan Allemann, I.; Baumann, L. Hyaluronic Acid Gel (Juvéderm) Preparations in the Treatment of Facial Wrinkles and Folds. Clin. Interv. Aging 2008, 3, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Weissmann, B.; Meyer, K. The Structure of Hyalobiuronic Acid and of Hyaluronic Acid from Umbilical Cord. J. Am. Chem. Soc. 1954, 76, 1753–1757. [Google Scholar] [CrossRef]
- Maclennan, A.P. The Production of Capsules, Hyaluronic Acid and Hyaluronidase by Group A and Group C Streptococci. Microbiology 1956, 14, 134–142. [Google Scholar] [CrossRef]
- Krzyściak, W.; Pluskwa, K.K.; Jurczak, A.; Kościelniak, D. The Pathogenicity of the Streptococcus Genus. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1361–1376. [Google Scholar] [CrossRef]
- Pavicic, T.; Gauglitz, G.G.; Lersch, P.; Schwach-Abdellaoui, K.; Malle, B.; Korting, H.C.; Farwick, M. Efficacy of Cream-Based Novel Formulations of Hyaluronic Acid of Different Molecular Weights in Anti-Wrinkle Treatment. J. Drugs Dermatol. 2011, 10, 990–1000. [Google Scholar]
- Jegasothy, S.M.; Zabolotniaia, V.; Bielfeldt, S. Efficacy of a New Topical Nano-Hyaluronic Acid in Humans. J. Clin. Aesthet. Dermatol. 2014, 7, 27–29. [Google Scholar]
- Nobile, V.; Buonocore, D.; Michelotti, A.; Marzatico, F. Anti-Aging and Filling Efficacy of Six Types Hyaluronic Acid Based Dermo-Cosmetic Treatment: Double Blind, Randomized Clinical Trial of Efficacy and Safety. J. Cosmet. Dermatol. 2014, 13, 277–287. [Google Scholar] [CrossRef]
- Cheng, K.; Zhou, Y.; Neelamegham, S. DrawGlycan-SNFG: A Robust Tool to Render Glycans and Glycopeptides with Fragmentation Information. Glycobiology 2017, 27, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Website Drawglycan. Available online: http://www.virtualglycome.org/DrawGlycan/ (accessed on 15 September 2023).
- Rougeaux, H.; Kervarec, N.; Pichon, R.; Guezennec, J. Structure of the Exopolysaccharide of Vibrio diabolicus Isolated from a Deep-Sea Hydrothermal Vent. Carbohydr. Res. 1999, 322, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Drouillard, S.; Jeacomine, I.; Laurine, B.; Claire, B.; Courtois, A.; Thollas, B.; Morvan, P.-Y.; Vallée, R.; Helbert, W. Structure of the Exopolysaccharide Secreted by a Marine Strain Vibrio alginolyticus. Mar. Drugs 2018, 16, 164. [Google Scholar] [CrossRef] [PubMed]
- Paximada, P.; Koutinas, A.A.; Scholten, E.; Mandala, I.G. Effect of Bacterial Cellulose Addition on Physical Properties of WPI Emulsions. Comparison with Common Thickeners. Food Hydrocoll. 2016, 54, 245–254. [Google Scholar] [CrossRef]
- Okamoto, T.; Kubota, K.; Kuwahara, N. Light Scattering Study of Gellan Gum. Food Hydrocoll. 1993, 7, 363–371. [Google Scholar] [CrossRef]
- Brownsey, G.J.; Chilvers, G.R.; Anson, K.I.; Morris, V.J. Some Observations (or Problems) on the Characterization of Gellan Gum Solutions. Int. J. Biol. Macromol. 1984, 6, 211–214. [Google Scholar] [CrossRef]
- Yadav, M.K.; Chae, S.-W.; Park, K.; Song, J.-J. Hyaluronic Acid Derived from Other Streptococci Supports Streptococcus pneumoniae in vitro Biofilm Formation. BioMed Res. Int. 2013, 2013, 690217. [Google Scholar] [CrossRef]
- Bassis, C.M.; Visick, K.L. The Cyclic-Di-GMP Phosphodiesterase BinA Negatively Regulates Cellulose-Containing Biofilms in Vibrio fischeri. J. Bacteriol. 2010, 192, 1269–1278. [Google Scholar] [CrossRef]
- Matthysse, A.G.; Holmes, K.V.; Gurlitz, R.H. Elaboration of Cellulose Fibrils by Agrobacterium tumefaciens during Attachment to Carrot Cells. J. Bacteriol. 1981, 145, 583–595. [Google Scholar] [CrossRef]
- Hungund, B.S.; Gupta, S.G. Improved Production of Bacterial Cellulose from Gluconacetobacter persimmonis GH-2. J. Microb. Biochem. Technol. 2010, 2, 127–133. [Google Scholar] [CrossRef]
- Bimmer, M.; Reimer, M.; Klingl, A.; Ludwig, C.; Zollfrank, C.; Liebl, W.; Ehrenreich, A. Analysis of Cellulose Synthesis in a High-Producing Acetic Acid Bacterium Komagataeibacter hansenii. Appl. Microbiol. Biotechnol. 2023, 107, 2947–2967. [Google Scholar] [CrossRef]
- Ude, S.; Arnold, D.L.; Moon, C.D.; Timms-Wilson, T.; Spiers, A.J. Biofilm Formation and Cellulose Expression among Diverse Environmental Pseudomonas Isolates. Environ. Microbiol. 2006, 8, 1997–2011. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Dazzo, F.; Hubbell, D. Production of Cellulose Microfibrils by Rhizobium. Appl. Microbiol. 1975, 30, 123–131. [Google Scholar] [CrossRef]
- Pacheco, G.; de Mello, C.V.; Chiari-Andréo, B.G.; Isaac, V.L.B.; Ribeiro, S.J.L.; Pecoraro, É.; Trovatti, E. Bacterial Cellulose Skin Masks-Properties and Sensory Tests. J. Cosmet. Dermatol. 2018, 17, 840–847. [Google Scholar] [CrossRef]
- Jeanes, A.; Haynes, W.C.; Wilham, C.A.; Rankin, J.C.; Melvin, E.H.; Austin, M.J.; Cluskey, J.E.; Fisher, B.E.; Tsuchiya, H.M.; Rist, C.E. Characterization and Classification of Dextrans from Ninety-Six Strains of Bacteria. J. Am. Chem. Soc. 1954, 76, 5041–5052. [Google Scholar] [CrossRef]
- Nácher-Vázquez, M.; Iturria, I.; Zarour, K.; Mohedano, M.L.; Aznar, R.; Pardo, M.Á.; López, P. Dextran Production by Lactobacillus sakei MN1 Coincides with Reduced Autoagglutination, Biofilm Formation and Epithelial Cell Adhesion. Carbohydr. Polym. 2017, 168, 22–31. [Google Scholar] [CrossRef]
- Patel, S.; Kasoju, N.; Bora, U.; Goyal, A. Structural Analysis and Biomedical Applications of Dextran Produced by a New Isolate Pediococcus pentosaceus Screened from Biodiversity Hot Spot Assam. Bioresour. Technol. 2010, 101, 6852–6855. [Google Scholar] [CrossRef]
- Ahmed, R.Z.; Siddiqui, K.; Arman, M.; Ahmed, N. Characterization of High Molecular Weight Dextran Produced by Weissella cibaria CMGDEX3. Carbohydr. Polym. 2012, 90, 441–446. [Google Scholar] [CrossRef]
- Kato, I. The Application of Sodium Dextran Sulfate to the Field of Cosmetics. Int. J. Cosmet. Sci. 2007, 29, 68. [Google Scholar] [CrossRef]
- Sun, G.; Zhang, X.; Shen, Y.-I.; Sebastian, R.; Dickinson, L.E.; Fox-Talbot, K.; Reinblatt, M.; Steenbergen, C.; Harmon, J.W.; Gerecht, S. Dextran Hydrogel Scaffolds Enhance Angiogenic Responses and Promote Complete Skin Regeneration during Burn Wound Healing. Proc. Natl. Acad. Sci. USA 2011, 108, 20976–20981. [Google Scholar] [CrossRef] [PubMed]
- Aman, A.; Siddiqui, N.N.; Qader, S.A.U. Characterization and Potential Applications of High Molecular Weight Dextran Produced by Leuconostoc mesenteroides AA1. Carbohydr. Polym. 2012, 87, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Robert, A.M.; Robert, L. Effect of a Fucose-Rich Polysaccharide Preparation on the Age-Dependent Evolution of the Skin Surface Micro-Relief. Pathol. Biol. 2003, 51, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Péterszegi, G.; Isnard, N.; Robert, A.M.; Robert, L. Studies on Skin Aging. Preparation and Properties of Fucose-Rich Oligo- and Polysaccharides. Effect on Fibroblast Proliferation and Survival. Biomed. Pharmacother. 2003, 57, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.; Alves, V.D.; Gouveia, A.R.; Pinheiro, C.; Torres, C.A.V.; Grandfils, C.; Reis, M.A.M. Controlled Production of Exopolysaccharides from Enterobacter A47 as a Function of Carbon Source with Demonstration of Their Film and Emulsifying Abilities. Appl. Biochem. Biotechnol. 2014, 172, 641–657. [Google Scholar] [CrossRef]
- Freitas, F.; Alves, V.D.; Torres, C.A.V.; Cruz, M.; Sousa, I.; Melo, M.J.; Ramos, A.M.; Reis, M.A.M. Fucose-Containing Exopolysaccharide Produced by the Newly Isolated Enterobacter Strain A47 DSM 23139. Carbohydr. Polym. 2011, 83, 159–165. [Google Scholar] [CrossRef]
- Torres, C.A.V.; Ferreira, A.R.V.; Freitas, F.; Reis, M.A.M.; Coelhoso, I.; Sousa, I.; Alves, V.D. Rheological Studies of the Fucose-Rich Exopolysaccharide FucoPol. Int. J. Biol. Macromol. 2015, 79, 611–617. [Google Scholar] [CrossRef]
- Baptista, S.; Pereira, J.R.; Guerreiro, B.M.; Baptista, F.; Silva, J.C.; Freitas, F. Cosmetic Emulsion Based on the Fucose-Rich Polysaccharide FucoPol: Bioactive Properties and Sensorial Evaluation. Colloids Surf. B Biointerfaces 2023, 225, 113252. [Google Scholar] [CrossRef]
- Guerreiro, B.M.; Freitas, F.; Lima, J.C.; Silva, J.C.; Reis, M.A.M. Photoprotective Effect of the Fucose-Containing Polysaccharide FucoPol. Carbohydr. Polym. 2021, 259, 117761. [Google Scholar] [CrossRef]
- Baptista, S.; Torres, C.A.V.; Sevrin, C.; Grandfils, C.; Reis, M.A.M.; Freitas, F. Extraction of the Bacterial Extracellular Polysaccharide FucoPol by Membrane-Based Methods: Efficiency and Impact on Biopolymer Properties. Polymers 2022, 14, 390. [Google Scholar] [CrossRef]
- Freitas, F.; Alves, V.D.; Pais, J.; Costa, N.; Oliveira, C.; Mafra, L.; Hilliou, L.; Oliveira, R.; Reis, M.A.M. Characterization of an Extracellular Polysaccharide Produced by a Pseudomonas Strain Grown on Glycerol. Bioresour. Technol. 2009, 100, 859–865. [Google Scholar] [CrossRef]
- Balkrishna, A.; Agarwal, V.; Kumar, G.; Gupta, A.K. Applications of Bacterial Polysaccharides with Special Reference to the Cosmetic Industry. In Microbial Bioprospecting for Sustainable Development; Singh, J., Sharma, D., Kumar, G., Sharma, N.R., Eds.; Springer: Singapore, 2018; pp. 189–202. ISBN 978-981-13-0053-0. [Google Scholar]
- Shih, I.-L.; Yu, Y.-T.; Shieh, C.-J.; Hsieh, C.-Y. Selective Production and Characterization of Levan by Bacillus subtilis (Natto) Takahashi. J. Agric. Food Chem. 2005, 53, 8211–8215. [Google Scholar] [CrossRef]
- Feingold, D.S.; Gehatia, M. The Structure and Properties of Levan, a Polymer of D-Fructose Produced by Cultures and Cell-Free Extracts of Aerobacter levanicum. J. Polym. Sci. 1957, 23, 783–790. [Google Scholar] [CrossRef]
- Poli, A.; Kazak, H.; Gürleyendağ, B.; Tommonaro, G.; Pieretti, G.; Öner, E.T.; Nicolaus, B. High Level Synthesis of Levan by a Novel Halomonas Species Growing on Defined Media. Carbohydr. Polym. 2009, 78, 651–657. [Google Scholar] [CrossRef]
- Jathore, N.R.; Bule, M.V.; Tilay, A.V.; Annapure, U.S. Microbial Levan from Pseudomonas fluorescens: Characterization and Medium Optimization for Enhanced Production. Food Sci. Biotechnol. 2012, 21, 1045–1053. [Google Scholar] [CrossRef]
- Newbrun, E.; Baker, S. Physico-Chemical Characteristics of the Levan Produced by Streptococcus salivarius. Carbohydr. Res. 1968, 6, 165–170. [Google Scholar] [CrossRef]
- Kim, K.H.; Chung, C.B.; Kim, Y.H.; Kim, K.S.; Han, C.S.; Kim, C.H. Cosmeceutical Properties of Levan Produced by Zymomonas mobilis. J. Cosmet. Sci. 2005, 56, 395–406. [Google Scholar] [CrossRef]
- Cambon-Bonavita, M.A.; Raguénès, G.; Jean, J.; Vincent, P.; Guezennec, J. A Novel Polymer Produced by a Bacterium Isolated from a Deep-sea Hydrothermal Vent Polychaete Annelid. J. Appl. Microbiol. 2002, 93, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Mancuso Nichols, C.A.; Garon, S.; Bowman, J.P.; Raguénès, G.; Guézennec, J. Production of Exopolysaccharides by Antarctic Marine Bacterial Isolates. J. Appl. Microbiol. 2004, 96, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Raguénès, G.; Peres, A.; Ruimy, R.; Pignet, P.; Christen, R.; Loaec, M.; Rougeaux, H.; Barbier, G.; Guezennec, J.G. Alteromonas infernus sp. Nov., a New Polysaccharide-Producing Bacterium Isolated from a Deep-Sea Hydrothermal Vent. J. Appl. Microbiol. 1997, 82, 422–430. [Google Scholar] [CrossRef]
- Rougeaux, H.; Guezennec, J.; Carlson, R.W.; Kervarec, N.; Pichon, R.; Talaga, P. Structural Determination of the Exopolysaccharide of Pseudoalteromonas Strain HYD 721 Isolated from a Deep-Sea Hydrothermal Vent. Carbohydr. Res. 1999, 315, 273–285. [Google Scholar] [CrossRef]
- Vincent, P.; Pignet, P.; Talmont, F.; Bozzi, L.; Fournet, B.; Guezennec, J.; Jeanthon, C.; Prieur, D. Production and Characterization of an Exopolysaccharide Excreted by a Deep-Sea Hydrothermal Vent Bacterium Isolated from the Polychaete Annelid Alvinella pompejana. Appl. Environ. Microbiol. 1994, 60, 4134–4141. [Google Scholar] [CrossRef]
- Zykwinska, A.; Marchand, L.; Bonnetot, S.; Sinquin, C.; Colliec-Jouault, S.; Delbarre-Ladrat, C. Deep-Sea Hydrothermal Vent Bacteria as a Source of Glycosaminoglycan-Mimetic Exopolysaccharides. Molecules 2019, 24, 1703. [Google Scholar] [CrossRef]
- Gedouin, P.A.; Vallée, R. Cosmetic or Pharmaceutical Composition, Useful for Treating Inflammation Including Erythema, Edema, Itching and Pain, Comprises an Exopolysaccharide Obtained from Cobetia marina Strain. France Patent FR2981847 B1, 15 November 2013. [Google Scholar]
- Gedouin, P.A.; Vallée, R.; Morvan, P.Y. An Ingredient of Marine Origin for Use in Cosmetic Compositions for Aged or Acne Skin. France Patent FR3100982 B1, 24 September 2021. [Google Scholar]
- Le Costaouëc, T.; Cérantola, S.; Ropartz, D.; Ratiskol, J.; Sinquin, C.; Colliec-Jouault, S.; Boisset, C. Structural Data on a Bacterial Exopolysaccharide Produced by a Deep-Sea Alteromonas macleodii Strain. Carbohydr. Polym. 2012, 90, 49–59. [Google Scholar] [CrossRef]
- Thibodeau, A.; Takeoka, A. The Applications and Functions of New Exopolysaccharide “Deepsane” from the Deepest Oceans. Fragr. J. 2006, 34, 61–68. [Google Scholar]
- Barbier, G.; Guezennec, J.; Pignet, P.; Bozzi, L.; Rinaudo, M.; Milas, M.; Leroy, Y.; Dubreucq, G.; Prieur, D.; Jeanthon, C.; et al. Alteromonas-Type Bacteria, Polysaccharides Produced by Said Bacteria, Ose Contained in Said Polysaccharides and Applications. Patent EP0682713 B1, 15 September 1999. [Google Scholar]
- Rougeaux, H.; Guezennec, J. Purified Alteromonas macleodii Polysaccharide and Its Uses. Patent EP1171625 B1, 19 January 2005. [Google Scholar]
- Fristsch, M.C.; Vacher, A.M. Cosmetic Composition Containing at Least a Polysaccharide from an Hydrothermal Bacterium. Patent EP0987010 B1, 8 September 2004. [Google Scholar]
- Gedouin, P.A.; Vallée, R. Antiwrinkle Cosmetic Composition Comprising an Exopolysaccharide from a Marine Bacterium. France Patent FR2975910 B1, 21 June 2013. [Google Scholar]
- Senni, K.; Sinquin, C.; Colliec-Jouault, S.; Godeau, G.-J.; Guezennec, J. Use of a Polysaccharide Which Is Excreted by the Vibrio diabolicus Species for the Engineering of Non-Mineralised Connective Tissue. Patent EP1960011 B1, 6 April 2016. [Google Scholar]
- Senni, K.; Gueniche, F.; Changotade, S.; Septier, D.; Sinquin, C.; Ratiskol, J.; Lutomski, D.; Godeau, G.; Guezennec, J.; Colliec-Jouault, S. Unusual Glycosaminoglycans from a Deep Sea Hydrothermal Bacterium Improve Fibrillar Collagen Structuring and Fibroblast Activities in Engineered Connective Tissues. Mar. Drugs 2013, 11, 1351–1369. [Google Scholar] [CrossRef]
- Colliec Jouault, S.; Chevolot, L.; Helley, D.; Ratiskol, J.; Bros, A.; Sinquin, C.; Roger, O.; Fischer, A.M. Characterization, Chemical Modifications and in vitro Anticoagulant Properties of an Exopolysaccharide Produced by Alteromonas infernus. Biochim. Biophys. Acta 2001, 1528, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.C.; Noiret, N.; Sinquin, C.; Ratiskol, J.; Guézennec, J.; Colliec-Jouault, S. Free-Radical Depolymerization with Metallic Catalysts of an Exopolysaccharide Produced by a Bacterium Isolated from a Deep-Sea Hydrothermal Vent Polychaete Annelid. Carbohydr. Polym. 2006, 64, 597–602. [Google Scholar] [CrossRef]
- Boisset, C.; Cozien, J.; Le Costaouec, T.; Sinquin, C.; Ratiskol, J.; Helbert, W.; Correc, G. Method of Depolymerisation of Natural Polysaccharides by Mechanical Grinding. France Patent FR2953217 B1, 5 October 2012. [Google Scholar]
- Petit, A.C.; Noiret, N.; Guezennec, J.; Gondrexon, N.; Colliec-Jouault, S. Ultrasonic Depolymerization of an Exopolysaccharide Produced by a Bacterium Isolated from a Deep-Sea Hydrothermal Vent Polychaete Annelid. Ultrason. Sonochem. 2007, 14, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Guezennec, J.; Pignet, P.; Lijour, Y.; Gentric, E.; Ratiskol, J.; Colliec-Jouault, S. Sulfation and Depolymerization of a Bacterial Exopolysaccharide of Hydrothermal Origin. Carbohydr. Polym. 1998, 37, 19–24. [Google Scholar] [CrossRef]
- Zykwinska, A.; Berre, L.T.-L.; Sinquin, C.; Ropartz, D.; Rogniaux, H.; Colliec-Jouault, S.; Delbarre-Ladrat, C. Enzymatic Depolymerization of the GY785 Exopolysaccharide Produced by the Deep-Sea Hydrothermal Bacterium Alteromonas infernus: Structural Study and Enzyme Activity Assessment. Carbohydr. Polym. 2018, 188, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Gedouin, P.A.; Vallée, R.; Gedouin, A.; Brehu, L. Method and Installation for the Controlled Partial Depolymerization of Polysaccharides. France Patent FR3053340 B1, 12 July 2019. [Google Scholar]
- Website CP Kelco. Available online: https://www.cpkelco.com/products/ (accessed on 18 October 2023).
- Website BASF. Available online: https://products.basf.com/global/en.html (accessed on 18 October 2023).
- Website Cargill. Available online: https://www.cargill.com/products (accessed on 18 October 2023).
- Website Safic Alcan. Available online: https://www.safic-alcan.com/en/fr/product-catalog/cosmetics-personal-care (accessed on 18 October 2023).
- Website Solabia. Available online: https://www.solabia.com/cosmetics/ (accessed on 18 October 2023).
- Website Codif Technologie Naturelle. Available online: https://www.codif-tn.com/nos-principes-actifs/ (accessed on 18 October 2023).
- Website Lubrizol. Available online: https://www.lubrizol.com/Personal-Care/Products/Product-Finder (accessed on 18 October 2023).
- Website Lucas-Meyer. Available online: https://lucasmeyercosmetics.com/fr/produits (accessed on 18 October 2023).
- Desbruyeres Daniel, L.L. Alvinella pompejana Gen. Sp. Nov., Ampharetidae Aberrant Des Sources Hydrothermales de La Ride Est-Pacifique. Oceanol. Acta 1980, 3, 267–274. [Google Scholar]
- Arahal, D.R.; Castillo, A.M.; Ludwig, W.; Schleifer, K.H.; Ventosa, A. Proposal of Cobetia Marina Gen. Nov., Comb. Nov., within the Family Halomonadaceae, to Include the Species Halomonas marina. Syst. Appl. Microbiol. 2002, 25, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant Production and Characterization of a Highly Active Alkaline Phosphatase from Marine Bacterium Cobetia marina. Mar. Biotechnol. 2015, 17, 130–143. [Google Scholar] [CrossRef]
- Mellado, E.; Moore, E.R.B.; Nieto, J.J.; Ventosa, A. Phylogenetic Inferences and Taxonomic Consequences of 16S Ribosomal DNA Sequence Comparison of Chromohalobacter marismortui, Volcaniella eurihalina, and Deleya salina and Reclassification of V. eurihalina as Halomonas eurihalina Comb. Nov. Int. J. Syst. Evol. Microbiol. 1995, 45, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Quesada, E.; Valderrama, M.J.; Bejar, V.; Ventosa, A.; Gutierrez, M.C.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Volcaniella eurihalina Gen. Nov., Sp. Nov., a Moderately Halophilic Nonmotile Gram-Negative Rod. Int. J. Syst. Evol. Microbiol. 1990, 40, 261–267. [Google Scholar] [CrossRef]
- Mata, J.A.; Béjar, V.; Llamas, I.; Arias, S.; Bressollier, P.; Tallon, R.; Urdaci, M.C.; Quesada, E. Exopolysaccharides Produced by the Recently Described Halophilic Bacteria Halomonas ventosae and Halomonas anticariensis. Res. Microbiol. 2006, 157, 827–835. [Google Scholar] [CrossRef]
- Martínez-Cánovas, M.J.; Béjar, V.; Martínez-Checa, F.; Quesada, E. Halomonas anticariensis Sp. Nov., from Fuente de Piedra, a Saline-Wetland Wildfowl Reserve in Málaga, Southern Spain. Int. J. Syst. Evol. Microbiol. 2004, 54, 1329–1332. [Google Scholar] [CrossRef]
- Yu, Y.; Li, H.; Wang, Y.; Zhang, Z.; Liao, M.; Rong, X.; Li, B.; Wang, C.; Ge, J.; Zhang, X. Antibiotic Resistance, Virulence and Genetic Characteristics of Vibrio alginolyticus Isolates from Aquatic Environment in Costal Mariculture Areas in China. Mar. Pollut. Bull. 2022, 185, 114219. [Google Scholar] [CrossRef]
- Radchenkova, N.; Vassilev, S.; Panchev, I.; Anzelmo, G.; Tomova, I.; Nicolaus, B.; Kuncheva, M.; Petrov, K.; Kambourova, M. Production and Properties of Two Novel Exopolysaccharides Synthesized by a Thermophilic Bacterium Aeribacillus pallidus 418. Appl. Biochem. Biotechnol. 2013, 171, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Spanò, A.; Laganà, P.; Visalli, G.; Maugeri, T.L.; Gugliandolo, C. In vitro Antibiofilm Activity of an Exopolysaccharide from the Marine Thermophilic Bacillus licheniformis T14. Curr. Microbiol. 2016, 72, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Spanò, A.; Gugliandolo, C.; Lentini, V.; Maugeri, T.L.; Anzelmo, G.; Poli, A.; Nicolaus, B. A Novel EPS-Producing Strain of Bacillus licheniformis Isolated from a Shallow Vent off Panarea Island (Italy). Curr. Microbiol. 2013, 67, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Panico, A.; Lama, L.; Gambacorta, A.; Nicolaus, B. A Halophilic Thermotolerant Bacillus Isolated from a Marine Hot Spring Able to Produce a New Exopolysaccharide. Biotechnol. Lett. 2002, 24, 515–519. [Google Scholar] [CrossRef]
- Nicolaus, B.; Panico, A.; Manca, M.C.; Lama, L.; Gambacorta, A.; Maugeri, T.; Gugliandolo, C.; Caccamo, D. A Thermophilic Bacillus Isolated from an Eolian Shallow Hydrothermal Vent Able to Produce Exopolysaccharides. Syst. Appl. Microbiol. 2000, 23, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Schiano Moriello, V.; Lama, L.; Poli, A.; Gugliandolo, C.; Maugeri, T.L.; Gambacorta, A.; Nicolaus, B. Production of Exopolysaccharides from a Thermophilic Microorganism Isolated from a Marine Hot Spring in Flegrean Areas. J. Ind. Microbiol. Biotechnol. 2003, 30, 95–101. [Google Scholar] [CrossRef]
- Sardari, R.R.R.; Kulcinskaja, E.; Ron, E.Y.C.; Björnsdóttir, S.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the Production of Exopolysaccharides by Two Strains of the Thermophilic Bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef]
- Nicolaus, B.; Schiano Moriello, V.; Lama, L.; Poli, A.; Gambacorta, A. Polysaccharides from Extremophilic Microorganisms. Orig. Life Evol. Biosph. 2004, 34, 159–169. [Google Scholar] [CrossRef]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-Activity Relationship of the Exopolysaccharide from a Psychrophilic Bacterium: A Strategy for Cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Finore, I.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Giudice, A. Lo Production and Biotechnological Potential of Extracellular Polymeric Substances from Sponge-Associated Antarctic Bacteria. Appl. Environ. Microbiol. 2018, 84, e01624-17. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Finore, I.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Isolation, Characterization and Optimization of EPSs Produced by a Cold-Adapted Marinobacter Isolate from Antarctic Seawater. Antarct. Sci. 2019, 31, 69–79. [Google Scholar] [CrossRef]
- Sun, M.-L.; Zhao, F.; Shi, M.; Zhang, X.-Y.; Zhou, B.-C.; Zhang, Y.-Z.; Chen, X.-L. Characterization and Biotechnological Potential Analysis of a New Exopolysaccharide from the Arctic Marine Bacterium Polaribacter sp. SM1127. Sci. Rep. 2016, 5, 18435. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular Polymeric Substances with Metal Adsorption Capacity Produced by Pseudoalteromonas sp. MER144 from Antarctic Seawater. Environ. Sci. Pollut. Res. 2018, 25, 4667–4677. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-B.; Chen, X.-L.; He, H.-L.; Zhang, X.-Y.; Xie, B.-B.; Yu, Y.; Chen, B.; Zhou, B.-C.; Zhang, Y.-Z. Structure and Ecological Roles of a Novel Exopolysaccharide from the Arctic Sea Ice Bacterium Pseudoalteromonas sp. Strain SM20310. Appl. Environ. Microbiol. 2013, 79, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Zhu, L.; Chen, X.; Wang, P.G.; Zhang, Y. Structural Characterization and Ecological Roles of a Novel Exopolysaccharide from the Deep-Sea Psychrotolerant Bacterium Pseudoalteromonas sp. SM9913. Microbiology 2007, 153, 1566–1572. [Google Scholar] [CrossRef]
- Carrión, O.; Delgado, L.; Mercade, E. New Emulsifying and Cryoprotective Exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Somayaji, A.; Dhanjal, C.R.; Lingamsetty, R.; Vinayagam, R.; Selvaraj, R.; Varadavenkatesan, T.; Govarthanan, M. An Insight into the Mechanisms of Homeostasis in Extremophiles. Microbiol. Res. 2022, 263, 127115. [Google Scholar] [CrossRef]
- López-Ortega, M.A.; Chavarría-Hernández, N.; del Rocío López-Cuellar, M.; Rodríguez-Hernández, A.I. A Review of Extracellular Polysaccharides from Extreme Niches: An Emerging Natural Source for the Biotechnology. From the Adverse to Diverse. Int. J. Biol. Macromol. 2021, 177, 559–577. [Google Scholar] [CrossRef]
- Marx, J.G.; Carpenter, S.D.; Deming, J.W. Production of Cryoprotectant Extracellular Polysaccharide Substances (EPS) by the Marine Psychrophilic Bacterium Colwellia psychrerythraea Strain 34H under Extreme Conditions. Can. J. Microbiol. 2009, 55, 63–72. [Google Scholar] [CrossRef]
- Nichols, C.M.; Bowman, J.P.; Guezennec, J. Effects of Incubation Temperature on Growth and Production of Exopolysaccharides by an Antarctic Sea Ice Bacterium Grown in Batch Culture. Appl. Environ. Microbiol. 2005, 71, 3519–3523. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.; Makshakova, O.; Angulo, J.; Bedini, E.; Bisio, A.; de Paz, J.L.; Fadda, E.; Guerrini, M.; Hricovini, M.; Hricovini, M.; et al. Glycosaminoglycans: What Remains to Be Deciphered? JACS Au 2023, 3, 628–656. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant and Antiradical Activities of L-Carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Tubia, C.; Fernández-Botello, A.; Dupont, J.; Gómez, E.; Desroches, J.; Attia, J.; Loing, E. A New Ex vivo Model to Evaluate the Hair Protective Effect of a Biomimetic Exopolysaccharide against Water Pollution. Cosmetics 2020, 7, 78. [Google Scholar] [CrossRef]
- Eom, S.J.; Lee, J.-A.; Kim, J.H.; Park, J.-T.; Lee, N.H.; Kim, B.-K.; Kang, M.-C.; Song, K.-M. Skin-Protective Effect of Polysaccharide from Ultrasonicated Sesame Oil Cake. Ind. Crops Prod. 2023, 203, 117123. [Google Scholar] [CrossRef]
- Mancuso Nichols, C.A.; Guezennec, J.; Bowman, J.P. Bacterial Exopolysaccharides from Extreme Marine Environments with Special Consideration of the Southern Ocean, Sea Ice, and Deep-Sea Hydrothermal Vents: A Review. Mar. Biotechnol. 2005, 7, 253–271. [Google Scholar] [CrossRef]
- Ruffing, A.; Chen, R.R. Metabolic Engineering of Microbes for Oligosaccharide and Polysaccharide Synthesis. Microb. Cell Factories 2006, 5, 25. [Google Scholar] [CrossRef]
- Essendoubi, M.; Gobinet, C.; Reynaud, R.; Angiboust, J.F.; Manfait, M.; Piot, O. Human Skin Penetration of Hyaluronic Acid of Different Molecular Weights as Probed by Raman Spectroscopy. Skin Res. Technol. 2016, 22, 55–62. [Google Scholar] [CrossRef]




| Bacterial EPS | Bacterial Strain | EPS Composition | Mw (g/mol) | Functional Properties | Ref. |
|---|---|---|---|---|---|
| Xanthan | Xanthomonas sp. | Anionic, branched Glc, Man, GlcA, Pyruvate, acetate | 0.4–15 × 106 | Hydrocolloid, binder, emulsion stabilizer, viscosity enhancer, thickening agent Skin conditioning agent | [1,33,73,74] |
| Gellan | Sphingomonas sp. | Anionic, linear Glc, Rha, GlcA Acetate, glycerate | 0.24–2.2 × 106 | Hydrocolloid, emulsion stabilizer, viscosity enhancer | [33,80,96,97] |
| Hyaluronic acid (HA) | Streptococcus sp. | Anionic, linear GlcA, GlcNAc | 2 × 106 | Viscosity enhancer, high water retention capacity Skin conditioning agent Bioactive: anti-wrinkle, moisturizing, skin elasticity enhancer, dermal filler | [34,84,85,86,88,89,98] |
| Cellulose (β-glucan) | Aliivibrio sp., Agrobacterium sp., Gluconacetobacter sp., Komagataeibacter sp., Pseudomonas sp., Rhizobium sp. | Neutral, linear Glc | 1 × 106 | Insoluble in aqueous solvents, highly crystalline, high degree of hydration, emulsion stabilizer Bioactive: moisturizer | [95,99,100,101,102,103,104,105] |
| Dextran | Lactobacillus sp., Leuconostoc sp., Pediococcus sp., Streptococcus sp., Weissella sp. | Neutral, linear Glc | 2–40 × 106 | Binder, bulking agent Bioactive: skin smoothing, brightening agent, anti-inflammatory | [106,107,108,109,110,111,112] |
| Fucogel | Klebsiella sp. | Anionic, linear Fuc, Gal, GalA Acetate | 4 × 104 | Skin conditioning agent Bioactive: skin moisturizing, anti-aging | [113,114] |
| FucoPol | Enterobacter A47 | Anionic, branched Fuc, Gal, Glc, GlcA Succinate, pyruvate, acetate | 2–6 × 106 | Hydrocolloid, emulsifying, flocculating and film-forming agent Bioactive: antioxidant, wound healing, photoprotection | [115,116,117,118,119,120] |
| GalactoPol | Pseudomonas sp. | Anionic, linear Gal, Man, Glc, Rha Succinate, pyruvate, acetate | 1–5 × 106 | Hydrocolloid, emulsifying, flocculating and film-forming agent | [121,122] |
| Levan | Aerobacter sp., Bacillus sp., Halomonas sp., Pseudomonas sp., Streptococcus sp., Zymomonas sp. | Neutral, linear or branched Fru | 2 × 106 | Water-soluble, strongly adhesive, film former, viscosity enhancer Skin conditioning agent Bioactive: anti-inflammatory, cell proliferative | [2,33,123,124,125,126,127,128] |
| Bacterial Strain | EPS Composition | Mw (g/mol) | Scopes of Action | Bioactivities | Refs. |
|---|---|---|---|---|---|
| A. macleodii subsp. fijiensis biovar deepsane HYD657 CNCM I-1285 | Anionic Gal, Glc, Rha, GlcA, GalA, Man, Fuc Sulfate, lactate, pyruvate | 1 × 106 | Soothing Irritation | Soothing effect; reduction of sensitive skin irritation by chemical, mechanical and UVB aggression; promotion of skin repair. | [129,137,138,139,140,141] |
| Alteromonas sp. CNCM I-4354 | Anionic GlcA, Glc, Gal, GalA, Man | 1 × 106 | Wrinkles | Wrinkle depth reduction; collagen fibers contraction inducing a tensing effect. | [142] |
| C. marina CNCM I-4353 | Anionic Glc, Rha, Gal, GlcA, GalA Sulfate | 1 × 106 | Soothing Inflammation | Inhibition and prevention of inflammation. | [135] |
| C. marina CNCM I-4353 | Anionic Glc, Rha, GlcNAc, GalA, Gal Sulfate 2 amino acids (threonine and serine) | 2 × 105 * | Barrier function Skin appearance Aging | Improvement of barrier function and moisturizing of the skin in the treatment of aged skin; improvement of skin repair kinetics against external aggressions. | [64] |
| H. anticariensis LMG P-27891 | Neutral or anionic Man, Rha, Glc Optional: GalA, Xyl | 1 × 104 | Inflammation Aging Wrinkles Skin firming | Treatment of cellulite; reduction of skin lipid accumulation; stimulation of lipolysis and collagen synthesis; reduction of the amount of nocturnin in cells. | [62] |
| H. eurihalina LMG P-28571 | Neutral or anionic Glc, GlcN, Man, Rha, Gal Optional: Fuc, GlcA Sulfate | 1 × 104 | Aging Wrinkles Skin firming | Promotion of collagen synthesis and connexins levels. | [4] |
| Pseudoalteromonas sp. CNCM I-4150 | Anionic Glc, Gal, GlcA, GlcNAc, GalA, Man | 8 × 105 * | Aging Wrinkles | Improvement of skin moisturizing due to the water retention capacity. | [69] |
| V. alginolyticus CNCM I-4151 | Anionic GalA, GlcNAc 2 amino acids (alanine and serine) | 2 × 105 * | Aging Inflammation Acne | Reduction of inflammation reduced; improvement of quality of the superficial layers of the epidermis; degradation of the extracellular matrix reduced. | [136] |
| V. alginolyticus CNCM I-5035 | Anionic Gal, GlcNAc, GulNAcA | 5 × 105 | Barrier function Acne | Improvement of physical and chemical barriers function by increasing the keratinocyte differentiation and epidermal renewal. Increasement of immune defense against pathogens involved in acne. | [65] |
| Vibrio sp. CNCM I-4239 | Anionic GlcNAc, GlcA, GalNAc | 1 × 105–1 × 106 * | Hydration Inflammation | Promotion of the healing process; inhibition of neuronal exocytosis (inflammation; acne; wrinkle reduction). | [71] |
| Vibrio sp. CNCM I-4277 | Anionic GlcA, GlcNAc, Glc, Fuc Sulfate | 1 × 106 | Aging Wrinkles | Increase of hyaluronic acid synthesis. | [70] |
| V. diabolicus CNCM I-1629 | GlcA, GlcNAc, GalNAc | 1 × 106 | Skin regeneration | Collagen structuring and extracellular matrix establishment by dermal fibroblasts | [93,143,144] |
| EPS | Examples of Trademarks | Refs. |
|---|---|---|
| Non-marine bacteria | Xanthan: Keldent®, Keltrol® (CP Kelco, Atlanta, GA, USA), Rheocare® (BASF, Ludwigshafen, Germany), Satiaxane (Cargill, Minneapolis, MN, USA), Safic’ Care T XG 80® (Safic Alcan, Puteaux, France) Gellan: Kelcogel® (CP Kelco, Atlanta, GA, USA) Fucogel® (Solabia, Pantin, France) | [152,153,154,155,156] |
| Marine bacteria | Epidermist 4.0TM, EPS BrightTM, EPS SeafillTM, EPS SeamatTM, EPS SeaglowTM, EPS SeapurTM (Codif Technologie Naturelle, Saint-Malo, France) Hyadisine®, Hyanify® (Lubrizol, Wickliffe, OH, USA) AbyssineTM PF, Exo-HTM, Exo-PTM, Exo-TTM (Lucas-Meyer, Québec, QC, Canada) | [157,158,159] |
| Strain | Claims | Cell Culture Model | Activity and Analysis | Refs. |
|---|---|---|---|---|
| C. marina CNCM I-4353 | Vascularization | Co-culture of human dermal fibroblasts (NHDF) and human umbilical vein endothelial cells (HUVECs) infected with a lentivirus that express green fluorescent protein | Quantification of angiogenesis (fluorescence levels expressed by HUVECs) | [64] |
| H. anticariensis LMG P-27891 | Anti-aging | Human dermal fibroblasts (2D) | Type I collagen synthesis (ELISA assay) | [62] |
| H. anticariensis LMG P-27891 | Slimming | Human subcutaneous pre-adipocytes in a complete differentiation medium (2D) | Reduction of the lipid accumulation “adipogenesis” (fluorescence assay) | [62] |
| V. alginolyticus CNCM I-4151 | Anti-inflammation | Skin explants inflamed by lipopolysaccharides addition | Interleukin production quantification (IL-8 levels of expression) | [136] |
| V. alginolyticus CNCM I-4151 | Anti-inflammation Anti-acne | Inflamed reconstructed human skin (3D) | Inflammation level studied by metalloproteinase expression (MMP3 mRNAs levels of expression) | [136] |
| V. alginolyticus CNCM I-4151 | Barrier function | Reconstructed aged human skin (3D) | Late Cornified Envelop Proteins (LCEs) proteins of the stratum corneum (gene expression of LCE3) | [136] |
| Vibrio sp. CNCM I-4277 | Moisturizing | Human dermal fibroblasts (2D) | Hyaluronic acid synthesis (ELISA assay) | [70] |
| Vibrio sp. CNCM I-4239 | Barrier function | Human keratinocytes (2D) | Healing test (microscopic observations of cells compared before and after treatment on the scrap region) | [71] |
| Vibrio sp. CNCM I-4239 | Cytotoxicity | Human dermal fibroblasts (2D) | Proliferation assay to measure cell viability (fluorescence assay) | [71] |
| V. diabolicus CNCM I-1629 | Promotion of fibroblast proliferation | Dermal equivalent matrices with human dermal fibroblasts (3D) | Proliferation and migration of fibroblasts and production of an extracellular matrix | [143,144] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benhadda, F.; Zykwinska, A.; Colliec-Jouault, S.; Sinquin, C.; Thollas, B.; Courtois, A.; Fuzzati, N.; Toribio, A.; Delbarre-Ladrat, C. Marine versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications. Mar. Drugs 2023, 21, 582. https://doi.org/10.3390/md21110582
Benhadda F, Zykwinska A, Colliec-Jouault S, Sinquin C, Thollas B, Courtois A, Fuzzati N, Toribio A, Delbarre-Ladrat C. Marine versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications. Marine Drugs. 2023; 21(11):582. https://doi.org/10.3390/md21110582
Chicago/Turabian StyleBenhadda, Fanny, Agata Zykwinska, Sylvia Colliec-Jouault, Corinne Sinquin, Bertrand Thollas, Anthony Courtois, Nicola Fuzzati, Alix Toribio, and Christine Delbarre-Ladrat. 2023. "Marine versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications" Marine Drugs 21, no. 11: 582. https://doi.org/10.3390/md21110582
APA StyleBenhadda, F., Zykwinska, A., Colliec-Jouault, S., Sinquin, C., Thollas, B., Courtois, A., Fuzzati, N., Toribio, A., & Delbarre-Ladrat, C. (2023). Marine versus Non-Marine Bacterial Exopolysaccharides and Their Skincare Applications. Marine Drugs, 21(11), 582. https://doi.org/10.3390/md21110582