Antioxidant Activities of Hydrolysates of Arca Subcrenata Prepared with Three Proteases

Abstract
:1. Introduction
2. Materials and methods
2.1 Materials
2.2 Enzymatic hydrolysis
2.3 Optimization of hydrolysis conditions
2.4 Analytical methods
2.4.1 Determination of the degree of hydrolysis
2.4.2 Determination of molecular mass distributions of hydrolysates
2.4.3 Determination of antioxidant activities
2.4.4 Size exclusion chromatography
2.4.5 Determination of free amino acids [20] and total amino acids of hydrolysates
2.4.6 Statistical analysis
3. Results and Discussion
3.1 Optimization of enzymatic hydrolysis conditions
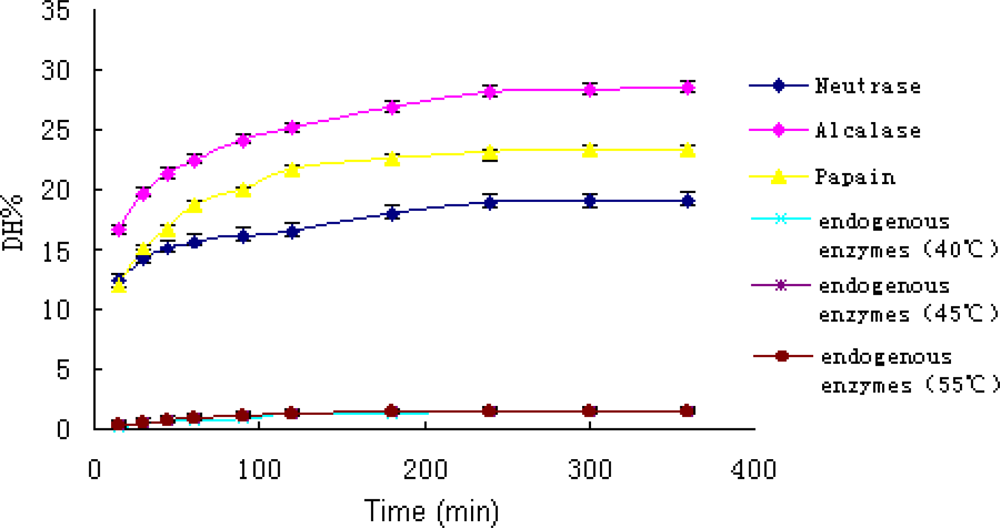
3.2 Effect of different enzymes on the degree of hydrolysis
3.3 Hydrolysate analysis by SDS-PAGE
3.4 Molecular mass distributions of three hydrolysates
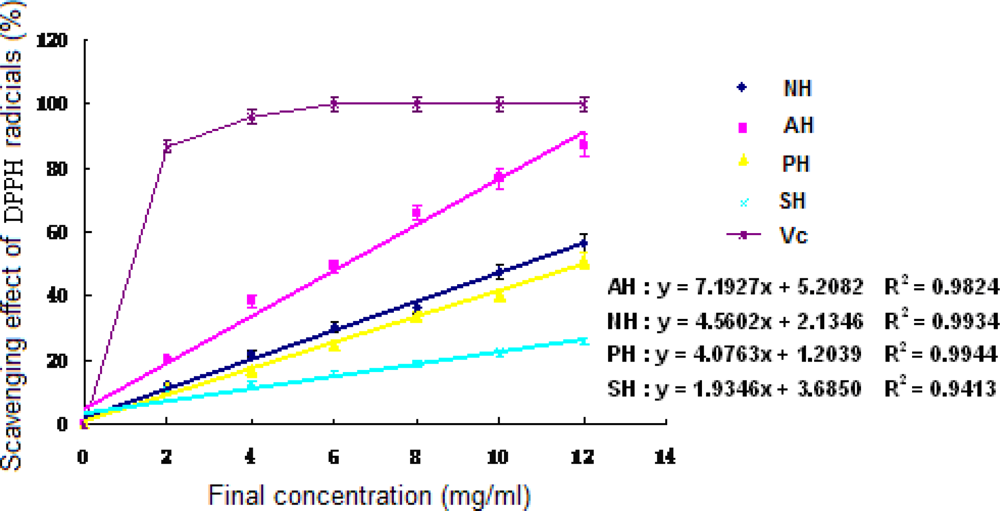
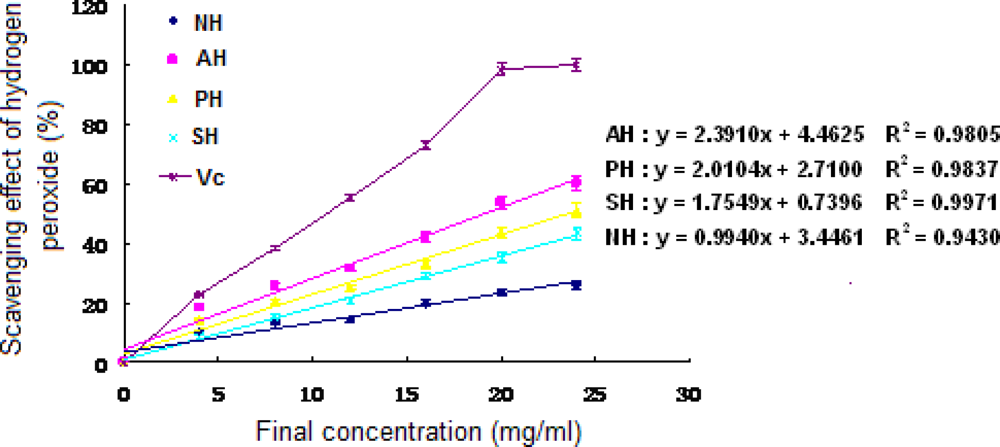
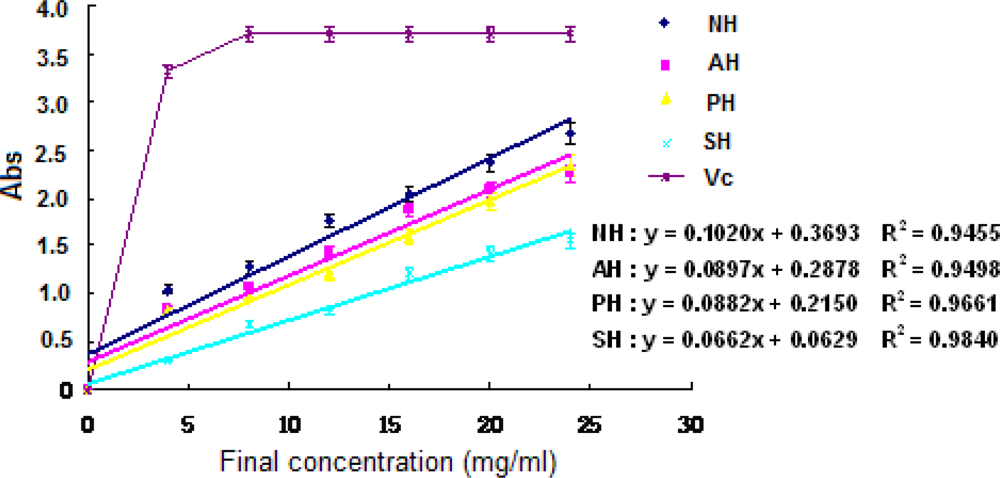
3.5 Antioxidant activities of hydrolysates
3.6 Determination of amino acid composition of hydrolysates
4. Conclusion
5. Acknowledgements
6. References
- Becker, GL. Preserving food and health: antioxidants make functional, nutritious preservatives. Food Processing 1993, 12, 54–56. [Google Scholar]
- Kitts, DD; Weiler, K. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Curr. Pharm. Design 2003, 9, 1309–1323. [Google Scholar]
- Suetsuna, K. Antioxidant peptides from the protease digest of prawn (Penaeus Japonicus) muscle. Mar. Biotechnol 2000, 2, 5–10. [Google Scholar]
- Jao, CL; Ko, WC. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) radical scavenging by protein hydrolysates from tuna cooking juice. Fisheries Sci 2002, 68, 430–435. [Google Scholar]
- Je, JY; Park, PJ; Kim, SK. Antioxidant activity of peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int 2005, 38, 45–50. [Google Scholar]
- Kim, SK; Kim, YT; Byun, HG; Nam, KS; Joo, DS; Shahidi, F. Isolation and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin. J. Agr. Food Chem 2001, 49, 1984–1989. [Google Scholar]
- Korhonen, H; Pihlanto-Leppala, A; Rantamaki, P; Tupasela, T. Impact of processing on bioactive proteins and peptides. Trends Food Sci. Tech 1998, 9, 307–319. [Google Scholar]
- Guo, XS; Li, Y. Marine Chinese Traditional Medicines; Sciences Press: Beijing. P.R. China, 2003; pp. 138–141. [Google Scholar]
- Li, Q; Li, TM; Wang, XQ; Huang, Q; Wu, WJ. Biochemical properties analysis of Arca subcrenata extractive. Pharm. Biotechnol 1998, 5, 245–247. [Google Scholar]
- Dou, CG; Yan, YQ; Zhang, Z. Experimental studies on hypoglycemia and hypolipid effects of hydrolysate of Arca subcrenata. Chin. J Mar. Drugs 1996, 15(1), 13–15. [Google Scholar]
- Song, LY; Ren, SF; Yu, RM; Yan, CY; Li, TF; Zhao, Y. Purification, characterization and in vitro anti-tumor activity of proteins from Arca subcrenata Lischke. Mar. Drugs 2008, 6, 418–430. [Google Scholar]
- Adler-Nissen, J. Enzymic hydrolysis of food proteins.; Elsevier Applied Science Publishers Ltd: London, 1986; p. 427. [Google Scholar]
- Mullally, MM; Meisel, H; FitzGerald, RJ. Angiotensin-I converting enzyme inhibitory activities of gastric and pancreatic proteinase digests of whey proteins. Int. Dairy J 1997, 7, 299–303. [Google Scholar]
- Guerard, F; Guimas, L; Binet, A. Production of tuna waste hydrolysates by a commercial neutral protease preparation. J. Mol. Catal. B-Enzym 2002, 19–20, 489–498. [Google Scholar]
- Schagger, H; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem 1987, 166, 368–379. [Google Scholar]
- Shimada, K; Fujikawa, K; Yahara, K; Nakamura, T. Antioxidative properties of xanthan on the antioxidation of soybean oil in cyclodextrin emulsion. J. Agr. Food Chem 1992, 40, 945–948. [Google Scholar]
- Ruch, RJ; Cheng, SJ; Klauning, JE. Prevention of cytotoxity and inhibition of intercellular communication by antioxidant catechin isolated from Chinese green tea. Carcinogenesis 1989, 10, 1003–1008. [Google Scholar]
- Oyaizu, M. Studies on products of browning reactions: antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutrition 1986, 44, 307–315. [Google Scholar]
- Tsai, JS; Lin, TC; Chen, JL; Pan, BS. The inhibitory effects of freshwater clam (Corbicula fluminea Muller) muscle protein hydrolysates on angiotensin I converting enzyme. Process Biochem 2006, 41, 2276–2281. [Google Scholar]
- He, HL; Chen, XL; Sun, CY; Zhang, YZ; Gao, PJ. Preparation and functional evaluation of oligopeptide-enriched hydrolysate from shrimp (Acetes chinensis) treated with crude protease from Bacillus sp. SM98011. Bioresource Technol 2006, 97, 385–390. [Google Scholar]
- Wang, J; Yuan, XP; Jin, ZY; Tian, Y; Song, HL. Free radical and reactive oxygen species scavenging activities of peanut skins extract. Food Chem 2007, 104, 242–250. [Google Scholar]
- Shahidi, F; Han, XQ; Synowiecki, J. Production and characteristics of protein hydrolysates from capelin (Mallotus villosus). Food Chem 1995, 53, 285–293. [Google Scholar]
- Yamaguchi, T; Takamura, H; Matoba, T; Terao, J. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1-diphenyl-2-picrylhydrazyl. Biosci. Biotech. Bioch 1998, 62, 1201–1204. [Google Scholar]
- Wang, JS; Zhao, MM; Zhao, QZ; Jiang, YM. Antioxidant properties of Papain hydrolysates of wheat gluten in different oxidation systems. Food Chem 2007, 101, 1658–1663. [Google Scholar]
- Gbogouri, GA; Linder, M; Fanni, J; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproduct hydrolysates. J. Food Sci 2004, 69, 615–622. [Google Scholar]
- Wu, HC; Chen, HM; Shiau, CY. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int 2003, 36, 949–957. [Google Scholar]
- Jun, SY; Park, PJ; Jung, WK; Kim, SK. Purification and characterization of an antioxidant from enzymatic hydrolysates of yellowfin sole (Limanda aspera) frame protein. Eur. Food Res. Technol 2004, 219, 20–26. [Google Scholar]
- Meisel, H. Overview on milk protein-derived peptides. Int. Dairy J 1998, 8, 363–373. [Google Scholar]
- Marcuse, R. Antioxidative effect of amino-acids. Nature 1960, 186, 886–887. [Google Scholar]
- Chen, HM; Muramoto, K; Yamauchi, F; Fujimoto, K; Nokihara, K. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. J. Agr. Food Chem 1998, 46, 49–53. [Google Scholar]
- Decker, EA; Crum, AD; Calvert, JT. Differences in the antioxidant mechanism of carnosine in the presence of copper and iron. J. Agr. Food Chem 1992, 40, 756–759. [Google Scholar]
- Chen, HM; Muramoto, K; Yamauchi, F. Structural analysis of antioxidative peptides from soybean β-conglycinin. J. Agr. Food Chem 1995, 43, 574–578. [Google Scholar]
- Saiga, A; Tanabe, S; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agr. Food Chem 2003, 51, 3661–3667. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Variable
| ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C)
| Time (h)
| E/S (%)
| pH
| DH (%)
| |||||||||||
| N | A | P | N | A | P | N | A | P | N | A | P | N | A | P | |
| 1 | 1(40) | 1(45) | 1(50) | 1(4) | 1(4) | 1(4) | 1(4.0) | 1(3.0) | 1(3.0) | 1(6.5) | 1(8.0) | 1(6.5) | 13.00 | 17.20 | 8.10 |
| 2 | 1 | 1 | 1 | 2(5) | 2(5) | 2(5) | 2(5.0) | 2(4.0) | 2(4.0) | 2(7.0) | 2(8.5) | 2(7.0) | 13.80 | 16.10 | 3.90 |
| 3 | 1 | 1 | 1 | 3(6) | 3(6) | 3(6) | 3(6.0) | 3(5.0) | 3(5.0) | 3(7.5) | 3(9.0) | 3(7.5) | 19.40 | 23.70 | 7.70 |
| 4 | 2(45) | 2(50) | 2(55) | 1 | 1 | 1 | 2 | 2 | 2 | 3 | 3 | 3 | 12.00 | 19.70 | 5.40 |
| 5 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 1 | 1 | 1 | 14.30 | 12.90 | 12.20 |
| 6 | 2 | 2 | 2 | 3 | 3 | 3 | 1 | 1 | 1 | 2 | 2 | 2 | 12.50 | 6.40 | 7.80 |
| 7 | 3(50) | 3(55) | 3(60) | 1 | 1 | 1 | 3 | 3 | 3 | 2 | 2 | 2 | 16.20 | 16.10 | 9.80 |
| 8 | 3 | 3 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 3 | 3 | 3 | 17.30 | 17.80 | 5.20 |
| 9 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 9.10 | 9.20 | 8.10 |
| K1 | 46.20 | 57.00 | 19.70 | 41.20 | 53.00 | 23.30 | 42.80 | 41.40 | 21.10 | 36.40 | 39.30 | 28.40 | |||
| K2 | 38.80 | 39.00 | 25.40 | 45.40 | 46.80 | 21.30 | 34.90 | 45.00 | 17.40 | 42.50 | 38.60 | 21.50 | |||
| K3 | 42.60 | 43.10 | 23.10 | 41.00 | 39.3 | 23.6 | 49.90 | 52.70 | 29.70 | 48.70 | 61.20 | 18.30 | |||
| R1 | 15.40 | 19.00 | 6.57 | 13.73 | 17.67 | 7.77 | 14.27 | 13.80 | 7.03 | 12.13 | 13.10 | 9.47 | |||
| R2 | 12.93 | 13.00 | 8.47 | 15.13 | 15.6 | 7.10 | 11.63 | 15.00 | 5.80 | 14.17 | 12.87 | 7.17 | |||
| R3 | 14.20 | 14.37 | 7.70 | 13.67 | 13.1 | 7.87 | 16.63 | 17.57 | 9.90 | 16.23 | 20.40 | 6.10 | |||
| S | 3.05 | 19.77 | 1.83 | 1.36 | 10.47 | 0.35 | 12.51 | 7.42 | 8.85 | 8.41 | 36.68 | 5.93 | |||
| Amino acid | Supernatant | NH | AH | PH | ||||||
| Ta | Fb | AAPc | Ta | Fb | AAPc | Ta | Fb | AAPc | ||
| Arg | 19.38 | 20.79 | 14.53 | 6.26 | 23.56 | 11.93 | 11.64 | 19.40 | 9.85 | 9.48 |
| Lys | 10.67 | 16.04 | 4.82 | 11.22 | 24.31 | 3.42 | 20.89 | 20.13 | 2.92 | 17.21 |
| Alad | 15.87 | 16.32 | 9.70 | 6.62 | 27.94 | 8.74 | 19.20 | 20.65 | 1.30 | 19.35 |
| Thr | 10.88 | 12.01 | 9.38 | 2.63 | 22.19 | 13.77 | 8.42 | 15.89 | 11.25 | 4.64 |
| Gly | 26.22 | 26.68 | 4.37 | 22.31 | 32.97 | 17.17 | 15.80 | 26.28 | 3.15 | 20.13 |
| Vald | 7.88 | 11.18 | 6.73 | 4.46 | 18.32 | - | 18.32 | 11.81 | 2.06 | 9.75 |
| Ser | 11.47 | 12.85 | - | 12.88 | 27.80 | - | 27.80 | 16.23 | - | 16.23 |
| Prod | 5.48 | 7.60 | 5.78 | 1.82 | 9.18 | 4.43 | 4.75 | 11.08 | 2.31 | 8.77 |
| Iled | 5.90 | 7.68 | 5.42 | 2.26 | 14.84 | 6.12 | 8.72 | 10.69 | 2.77 | 7.92 |
| Leud | 16.31 | 22.34 | 10.85 | 11.50 | 37.58 | 5.80 | 31.78 | 28.53 | 3.96 | 24.57 |
| Metd | 4.09 | 6.16 | 3.87 | 2.29 | 11.26 | 2.75 | 8.51 | 8.04 | 1.74 | 6.29 |
| His | 3.15 | 4.09 | 3.83 | 0.26 | 6.61 | 0.85 | 5.76 | 5.03 | 3.68 | 1.35 |
| Phed | 5.46 | 6.64 | 3.54 | 3.09 | 11.60 | - | 11.60 | 6.56 | 2.99 | 3.57 |
| Gln | 34.26 | 42.04 | 7.18 | 34.87 | 66.05 | 6.15 | 59.90 | 50.41 | 2.99 | 47.42 |
| Asn | 19.56 | 26.31 | 1.40 | 24.91 | 40.99 | 0.64 | 40.36 | 33.03 | 0.75 | 32.28 |
| Cys | 0.23 | 0.96 | 0.11 | 0.84 | 1.15 | - | 1.15 | 0.68 | - | 0.68 |
| Tyr | 3.61 | 3.77 | 0.33 | 3.44 | 8.82 | 2.61 | 6.22 | 5.24 | 1.95 | 3.29 |
| Total | 200.42 | 243.46 | 91.84 | 151.66 | 385.17 | 84.38 | 300.82 | 289.68 | 53.67 | 232.93 |
Share and Cite
Song, L.; Li, T.; Yu, R.; Yan, C.; Ren, S.; Zhao, Y. Antioxidant Activities of Hydrolysates of Arca Subcrenata Prepared with Three Proteases. Mar. Drugs 2008, 6, 607-619. https://doi.org/10.3390/md6040607
Song L, Li T, Yu R, Yan C, Ren S, Zhao Y. Antioxidant Activities of Hydrolysates of Arca Subcrenata Prepared with Three Proteases. Marine Drugs. 2008; 6(4):607-619. https://doi.org/10.3390/md6040607
Chicago/Turabian StyleSong, Liyan, Tingfei Li, Rongmin Yu, Chunyan Yan, Shengfang Ren, and Yu Zhao. 2008. "Antioxidant Activities of Hydrolysates of Arca Subcrenata Prepared with Three Proteases" Marine Drugs 6, no. 4: 607-619. https://doi.org/10.3390/md6040607
APA StyleSong, L., Li, T., Yu, R., Yan, C., Ren, S., & Zhao, Y. (2008). Antioxidant Activities of Hydrolysates of Arca Subcrenata Prepared with Three Proteases. Marine Drugs, 6(4), 607-619. https://doi.org/10.3390/md6040607