Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research

Abstract
:1. Introduction
2. Cylindrospermopsin
2.1. Properties
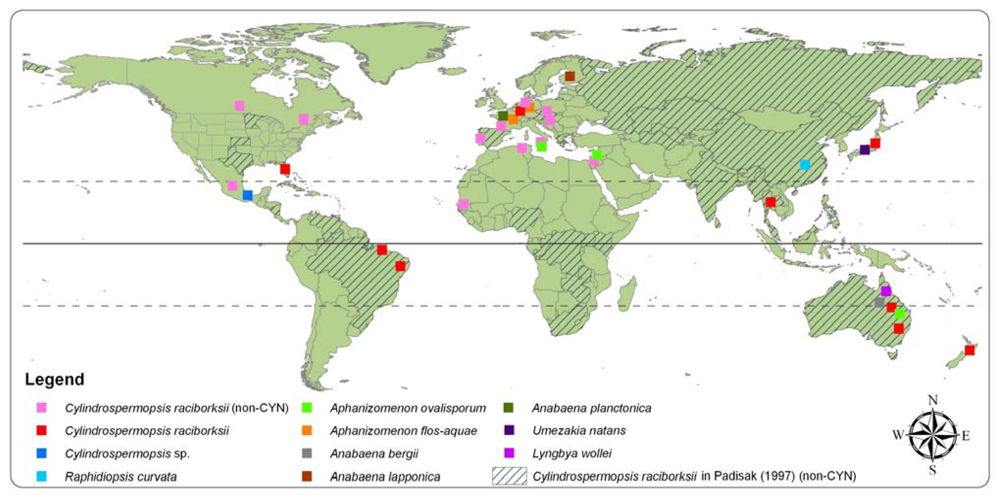
2.2. Distribution and detection
2.3. Studies on mechanism of toxicity
2.4. Human health effects
2.5. Ecological effects
3. Bioaccumulation of CYN
3.1. Existing evidence
3.2. Patterns of toxin uptake and deposition
3.3. Factors influencing bioaccumulation rates and potential
3.4. Field monitoring of bioaccumulation
3.5. Risk assessments and bioaccumulation
4. Where to Next?
5. Conclusions
Acknowledgements
References and Notes
- Komárek, J; Anagnostidis, K. Modern approach to the classification system of cyanophytes. Arch Hydrobiol 1986, 157–226. [Google Scholar]
- Whitton, BA; Potts, M. The Ecology of Cyanobacteria Their Diversity in Time and Space; Kluwer Academic Publishers: Dordrecht, The Netherland, 2000; p. 669. [Google Scholar]
- Shaw, G; Lam, PKS. Health aspects of freshwater cyanobacterial toxins. Water Sci Technol: Water Supply 2007, 7, 193–203. [Google Scholar]
- Rellán, S; Osswald, J; Saker, M; Gago-Martinez, A; Vasconcelos, V. First detection of anatoxin-a in human and animal dietary supplements containing cyanobacteria. Food Chem Toxicol 2009, 47, 2189–2195. [Google Scholar]
- Chorus, I. Water Safety Plans A better regulatory approach to prevent human exposure to harmful cyanobacteria. In Harmful Cyanobacteria; Huisman, J, Matthijs, HCP, Visser, PM, Eds.; Springer: Dordrecht, The Netherlands, 2005b; pp. 201–226. [Google Scholar]
- Chorus, I; Falconer, IR; Salas, HJ; Bartram, J. Health risks caused by freshwater cyanobacteria in recreational waters. J Toxicol Environ Health: Part B 2000, 3, 323–347. [Google Scholar]
- Schwimmer, M; Schwimmer, D. Medial aspects of phycology. In Algae, Man and the Environment; Jackson, DF, Ed.; Syracuse University Press: New York, NY, USA, 1968; pp. 279–358. [Google Scholar]
- Beasley, VR; Cook, WO; Dahlem, AM; Hooser, SB; Lovell, RA; Valentine, WM. Algae intoxication in livestock and waterfowl. Vet Clin North Am Food Anim Pract 1989, 5, 345–361. [Google Scholar]
- Carmichael, WW; Falconer, IR. Diseases related to freshwater blue-green algal toxins, and control measures. In Algal Toxins in Seafood and Drinking Water; Falconer, IR, Ed.; Academic Press: Sydney, Australia, 1993; pp. 187–209. [Google Scholar]
- Duy, TN; Lam, PKS; Shaw, GR; Connell, DW. Toxicology and risk assessment of freshwater cyanobacterial (blue-green algal) toxins in water. Rev Environ Contam Toxicol 2000, 163, 113–186. [Google Scholar]
- Ibelings, BW; Havens, KE. Cyanobacterial toxins: a qualitative meta-analysis of concentrations, dosage and effects in freshwater, estuarine and marine biota. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer Press: New York, NY, USA, 2008; pp. 675–732. [Google Scholar]
- Ibelings, BW; Chorus, I. Accumulation of cyanobacterial toxins in freshwater “seafood” and its consequences for public health: A review. Environ Pollut 2007, 150, 177–192. [Google Scholar]
- Ohtani, I; Moore, RE; Runnegar, MTC. Cylindrospermopsin: a potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J Am Chem Soc 1992, 114, 7941–7942. [Google Scholar]
- Norris, RLG; Eaglesham, GK; Pierens, G; Shaw, GR; Smith, MJ; Chiswell, RK; Seawright, AA; Moore, MR. Deoxycylindropermopsin, an analog of cylindropermopsin from Cylindrospermopsis raciborskii. Environ Toxicol 1999, 14, 163–165. [Google Scholar]
- Banker, R; Carmeli, S; Teltsch, B; Sukenik, A. 7-epicylindrospermopsin, a toxic minor metabolite of the cyanobacterium Aphanizomenon ovalisporum from Lake Kinneret, Israel. J Nat Prod 2000, 63, 387–389. [Google Scholar]
- Norris, RL; Eaglesham, G; Pierens, G; Shaw, G; Smith, MJ; Chiswell, RK; Seawright, AA; Moore, MR. Deoxycylindrospermopsin, an analog of cylindrospermopsin from Cylindrospermopsis raciborskii. Environ Toxicol 1999, 14, 163–165. [Google Scholar]
- Chiswell, RK; Shaw, GR; Eaglesham, G; Smith, MJ; Norris, KR; Seawright, AA; Moore, MR. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature and sunlight on decomposition. Environ Toxicol 1999, 14, 155–161. [Google Scholar]
- Sivonen, K; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water; Chorus, I, Bartram, J, Eds.; E & FN Spoon: London, UK, 1999. [Google Scholar]
- Shaw, G; Seawright, AA; Moore, MR; Lam, PKS. Cylindrospermopsin, a cyanobacterial alkaloid: evaluation of its toxicologic activity. Ther Drug Monit 2000, 22, 89–92. [Google Scholar]
- Seifert, M; McGregor, G; Eaglesham, G; Wickramasinghe, W; Shaw, G. First evidence for the production of cylindrospermopsin and deoxy-cylindrospermopsin by the freshwater benthic cyanobacterium, Lyngbya wollei (Farlow ex Gomont) Speziale and Dyck. Harmful Algae 2007, 6, 73–80. [Google Scholar]
- Padisák, J. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding highly adaptive cyanobacterium: worldwide distribution and review of its ecology. Arch Hydrobiol 1997, 4, 563–593, Supplement 107 (Monographic Studies). [Google Scholar]
- Rücker, J; Stüken, A; Nixdorf, B; Fastner, J; Chorus, I; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar]
- Kling, HJ. Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria): A brief historic overview and recent discovery in the assiniboine river (Canada). Fottea 2009, 9, 45–47. [Google Scholar]
- Griffiths, DJ; Saker, ML. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ Toxicol 2003, 18, 78–93. [Google Scholar]
- McGregor, GB; Fabbro, LD. Dominance of Cylindrospermopsis raciborskii (Nostocales, Cyanoprokaryota) in Queensland tropical and subtropical reservoirs: Implications for monitoring and management. Lakes Reserv Res Manage 2000, 5, 195–205. [Google Scholar]
- Hawkins, PR; Putt, E; Falconer, IR; Humpage, AR. Phenotypical variation in a toxic strain of the phytoplankter, Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) during batch culture. Environ Toxicol 2001, 16, 460–467. [Google Scholar]
- Stirling, DJ; Quilliam, MA. First report of the cyanobacterial toxin cylindrospermopsin in New Zealand. Toxicon 2001, 39, 1219–1222. [Google Scholar]
- Richey, LJ; Carbonneau, DA; Schoeb, TR; Taylor, SK; Woodward, AR; Clemmons, R. Potential Toxicity of Cyanobacteria to American Alligators (Alligator mississippiensis) 19.
- Chapman, AD; Schelske, CL. Recent appearance of Cylindrospermopsis (cyanobacteria) in five hypereutrophic Florida lakes. J Phycol 1997, 33, 191–195. [Google Scholar]
- Hamilton, PB; M, LL; Dean, S; Pick, FR. The occurrence of the cyanobaterium Cylindrospermopsis raciborskii in Constance Lake: an exotic cyanoprokaryote new to Canada. Phycologia 2005, 44, 17–25. [Google Scholar]
- Alster, A; Kaplan-Levy, RN; Sukenik, A; Zohary, T. Morphology and phylogeny of a nontoxic invasive Cylindrospermopsis raciborskii from a Mediterranean Lake. Hydrobiol 2009, 1–14. [Google Scholar]
- Li, R; Carmichael, WW; Brittain, JE; Eaglesham, GK; Shaw, GR; Mahakhant, A; Noparatnaraporn, N; Yongmanitchai, W; Kaya, K; Watanabe, MM. Isolation and identification of the cyanotoxin cylindrospermopsin and deoxycylindrospermopsin from a Thailand strain of Cylindrospermopsis raciborskii (Cyanobacteria). Toxicon 2001, 39, 973–980. [Google Scholar]
- Bernard, C; Harvey, M; Biré, R; Krys, S; Fontaine, JJ. Toxicological comparison of diverse Cylindrospermopsis raciborskii strains: Evidence of liver damage caused by a French C. raciborskii strain. Environ Toxicol 2003, 18, 176–186. [Google Scholar]
- Chonudomkul, D; Yongmanitchai, W; Theeragool, G; Kawachi, M; Kasai, F; Kaya, K; Watanabe, MM. Morphology, genetic diversity, temperature tolerance, and toxicity of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) strains from Thailand and Japan. FEMS Microbiol Ecol 2004, 48, 345–355. [Google Scholar]
- Barone, R; Castelli, G; Naselli-Flores, L. Red sky at night cyanobacteria delight: the role of climate in structuring phytoplankton assemblage in a shallow, Mediterranean lake (Biviere di Gela, southeastern Sicily). Hydrobiologia 2009, 1–11. [Google Scholar]
- Molica, R; Onodera, H; Garcia, C; Rivas, M; Andrinolo, D; Nascimento, SM; Meguro, H; Oshima, Y; Azevedo, S; Lagos, N. Toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii (Cyanophyceae) isolated from Tabocas Reservoir in Caruaru, Brazil, including demonstration of a new saxitoxin analogue. Phycologia 2002, 41, 606–611. [Google Scholar]
- Bouvy, M; Falcão, D; Marinho, M; Pagano, M; Moura, A. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquat Microb Ecol 2000, 23, 13–27. [Google Scholar]
- Komárková, J; Laudares-Silva, R; Senna, PA. Extreme morphology of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) in the Lagoa do Peri, a freshwater coastal lagoon, Santa Catarina, Brazil. Algol Stud 1999, 94, 207–222. [Google Scholar]
- Briand, JF; Robillot, C; Quiblier-Llobéras, C; Humbert, JF; Couté, A; Bernard, C. Environmental context of Cylindrospermopsis raciborskii (Cyanobacteria) blooms in a shallow pond in France. Water Res 2002, 36, 3183–3192. [Google Scholar]
- Saker, ML; Nogueira, ICG; Vasconcelos, VM; Neilan, BA; Eaglesham, GK; Pereira, P. First report and toxicological assessment of the cyanobacterium Cylindrospermopsis raciborskii from Portuguese freshwaters. Ecotox Environ Safety 2003, 55, 243–250. [Google Scholar]
- Valério, E; Pereira, P; Saker, ML; Franca, S; Tenreiro, R. Molecular characterization of Cylindrospermopsis raciborskii strains isolated from Portuguese freshwaters. Harmful Algae 2005, 4, 1044–1052. [Google Scholar]
- Fastner, J; Heinze, R; Humpage, AR; Mischke, U; Eaglesham, GK; Chorus, I. Cylindrospermopsin occurrence in two German lakes and preliminary assessment of toxicity and toxin production of Cylindrospermopsis raciborskii (Cyanobacteria) isolates. Toxicon 2003, 42, 313–321. [Google Scholar]
- Fastner, J; Rücker, J; Stüken, A; Preußel, K; Nixdorf, B; Chorus, I; Köhler, A; Wiedner, C. Occurrence of the cyanobacterial toxin cylindrospermopsin in northeast Germany. Environ Toxicol 2007, 22, 26–32. [Google Scholar]
- Briand, JF; Leboulanger, C; Humbert, J.-F; Bernard, C; Dufour, P. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: selection; wide physiological tolerance, or global warming? J Phycol 2004, 40, 231–238. [Google Scholar]
- Dvořák, P; Haŝler, P. Occurrence and morphological variability of Cylindrospermopsis raciborskii (Wolosz.) Seenayya et Subba Raju (Cyanophyta, Nostocales) near Olomouc in 2006. Fottea 2007, 7, 39–42. [Google Scholar]
- Stüken, A; Rücker, J; Endrulat, T; Preussel, K; Hemm, M; Nixdorf, B; Karsten, U; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar]
- St Amand, A. Map of selected sires in the United States where Cylindrosermopsis raciborskii has been encountered. Lakeline 2002, 22. [Google Scholar]
- Berry, JP; Lind, O. First evidence of “paralytic shellfish toxins” and cylindrospermopsin in a Mexican freshwater system, Lago Catemaco, and apparent bioaccumulation of the toxins in “tegogolo” snails (Pomacea patula catemacensis). Toxicon 2010, in press. [Google Scholar]
- Li, R; Carmichael, WW; Brittain, S; Eaglesham, G; Shaw, G; Liu, Y; Watanabe, M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (Cyanobacteria). J Phycol 2001, 37, 1121–1126. [Google Scholar]
- Shaw, G; Sufenik, A; Livne, A; Chiswell, RK; Smith, MJ; Seawright, AA; Norris, KR; Eaglesham, G; Moore, MR. Blooms of the cylindrospermopsin containing cyanobacterium, Aphanizomenon ovalisporum (Forti) in newly constructed lakes, Queensland, Australia. Environ Toxicol 1999, 14, 167–177. [Google Scholar]
- Banker, R; Carmeli, S; Werman, M; Teltsch, B; Porat, R; Sukenik, A. Uracil moiety is required for toxicity of the cyanobacterial hepatotoxin cylindrospermopsin. J Toxicol Environ Health: Part A 2001, 62, 281–288. [Google Scholar]
- Preußel, K; Stüken, A; Wiedner, C; Chorus, I; Fastner, J. First report on cylindrospermopsin producing Aphanizomenon flos-aquae (Cyanobacteria) isolated from two German lakes. Toxicon 2006, 47, 156–162. [Google Scholar]
- Falconer, IR; Humpage, AR. Cyanobacterial (blue-green algal) toxins in water supplies: cylindrospermopsins. Environ Toxicol 2006, 21, 299–304. [Google Scholar]
- Fergusson, KM; Saint, CP. Multiplex PCR assay for Cylindrospermopsis raciborskii and cylindrospermopsin-producing cyanobacteria. Environ Toxicol 2003, 18, 120–125. [Google Scholar]
- Spoof, L; Berg, KA; Rapala, J; Lahti, K; Lepistö, L; Metcalf, JS; Codd, GA; Meriluoto, J. First observation of cylindrspermopsin in Anabaena lapponica isolated from the Boreal Environment (Finland). Environ Toxicol 2006, 21, 552–560. [Google Scholar]
- Terao, K; Ohmori, S; Igarshi, K; Ohtani, I; Watanabe, MF; Harada, KI; Ito, E; Watanabe, M. Electron microscope studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon 1994, 32, 833–843. [Google Scholar]
- Harada, KI. Chemistry and detection of microcystins. In Toxic Microcystis; Watanabe, M, Harada, KI, Carmichael, WW, Fujiki, H, Eds.; CRC Press: Boca Raton, FL USA, 1996; pp. 103–148. [Google Scholar]
- Brient, L; Lengronne, M; Bormans, M; Fastner, J. First occurrence of cylindrospermopsin in freshwater in France. Environ Toxicol 2009, 24, 415–420. [Google Scholar]
- Bouaïcha, N; Nasri, A.-B. First report of Cyanobacterium Cylindrospermopsis raciborskii from Algerian Freshwaters. Environ Toxicol 2004, 19, 541–543. [Google Scholar]
- Garnett, C; Shaw, G; Moore, D; Florian, P; Moore, M. Impact of climate change on toxic cyanobacterial (blue-green algal) blooms and algal toxin production in Queensland 111.
- Neilan, BA; Saker, ML; Fastner, J; Törökné, AK; Burns, BP. Phylogeography of the invasive cyanobacterium Cylindrospermopsis raciborskii. Mol Ecol 2003, 12, 133–140. [Google Scholar]
- Wiedner, C; Rücker, J; Brüggemann, R; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar]
- Harada, KI; Ohtani, I; Iwamoto, K; Suzuki, M; Watanabe, MF; Watanabe, M; Terao, K. Isolation of cylindrospermopsin from a cyanobacterium Umezakia natans and its screening method. Toxicon 1994, 32, 73–84. [Google Scholar]
- Hawkins, PR; Chandrasena, NR; Jones, GJ; Humpage, AR; Falconer, IR. Isolation and toxicity of Cylindrospermopsis raciborskii from an ornamental lake. Toxicon 1997, 35, 341–346. [Google Scholar]
- Carson, B. Cylindrospermopsin-A Review of Toxicological Literature; National Institute of Environmental Health Services: Research Triangle Park, NC, USA, 29 April 2003. [Google Scholar]
- Eaglesham, G; Norris, KR; Shaw, GR; Smith, MJ; Chiswell, RK; Davis, BC; Neville, GR; Seawright, AA; Moore, BS. Use of HPLC-MS/MS to monitor cylindrospermopsin, a blue-green algal toxin, for public health purposes. Environ Toxicol 1999, 14, 151–154. [Google Scholar]
- Nicholson, BC; Burch, MD. Evaluation of analytical methods for detection and quantification of cyanotoxins in relation to australian drinking water guidelines 2001.
- Welker, M; Bickel, H; Fastner, J. HPLC-PDA detection of cylindrospermopsin - opportunities and limits. Water Res 2002, 36, 4659–4663. [Google Scholar]
- Bláhová, L; Oravec, M; Marsálek, B; Sejnohová, L; Simek, Z; Bláha, L. The first occurrence of the cyanobacterial alkaloid toxin cylindrospermopsin in the Czech Republic as determined by immunochemical and LC/MS methods. Toxicon 2009, 53, 519–524. [Google Scholar]
- Humpage, AR; Falconer, IR. Oral toxicity of the Cyanobacterial Toxin Cylindrospermopsin in Male Swiss Albino Mice: Determination of No Observed Adverse Effect Level for Deriving a Drinking Water Guideline. Environ Toxicol 2003, 18, 94–103. [Google Scholar]
- Seawright, AA; Nolan, CC; Shaw, GR; Chiswell, RK; Norris, RL; Moore, MR; Smith, MJ. The oral toxicity for mice of the tropical cyanobacterium Cylindrospermopsis raciborskii (Woloszynska). Environ Toxicol 1999, 14, 135–142. [Google Scholar]
- Froscio, SM; Humpage, AR; Burcham, PC; Falconer, IR. Cylindrospermopsin-induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ Toxicol 2003, 18, 243–251. [Google Scholar]
- Kinnear, SHW; Fabbro, LD; Duivenvoorden, LJ; Hibberd, EMA. Multiple-organ toxicity resulting from cylindrospermopsin exposure in tadpoles of the cane toad (Bufo marinus). Environ Toxicol 2007, 22, 550–558. [Google Scholar]
- Rogers, EH; Zehr, RD; Gage, MI; Humpage, AR; Falconer, IR; Marr, M; Chernoff, N. The cyanobacterial toxin, cylindrospermopsin, induces fetal toxicity in the mouse after exposure late in gestation. Toxicon 2007, 49, 855–864. [Google Scholar]
- Hawkins, PR; Runnegar, MTC; Jackson, ARB; Falconer, IR. Severe hepatotoxicity caused by the tropical cyanobacterium (blue-green alga) Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Subba Raju isolated from a domestic water supply reservoir. Appl Env Microbiol 1985, 50, 1292–1295. [Google Scholar]
- Kinnear, SHW; Duivenvoorden, LJ; Fabbro, LD. Sublethal responses in Melanoides tuberculata following exposure to Cylindrospermopsis raciborskii containing cylindrospermopsin. Harmful Algae 2007, 6, 642–650. [Google Scholar]
- Nogueira, ICG; Lobo-da-Cunha, A; Vasconcelos, VM. Effects of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum (cyanobacteria) ingestion on Daphnia magna midgut and associated diverticula epithelium. Aquat Toxicol 2006, 80, 194–203. [Google Scholar]
- Seifert, M. The ecological effects of the cyanobacterial toxin cylindrospermopsin; The University of Queensland: Brisbane, Australia, 2007. [Google Scholar]
- Rasmussen, JP; Cursaro, M; Froscio, SM; Saint, CP. An examination of the antibiotic effects of cylindrospermopsin on common gram-positive and gram-negative bacteria and the protozoan Naegleria lovaniensis. Environ Toxicol 2008, 23, 36–43. [Google Scholar]
- Saker, M; Thomas, AD; Norton, JH. Cattle mortality attributed to the toxic cyanobacterium Cylindrospermopsis raciborskii in an outback region of North Queensland. Environ Toxicol 1999, 14, 179–182. [Google Scholar]
- Metcalf, JS; Barakate, A; Codd, GA. Inhibition of plant protein synthesis by the cyanobacterial hepatotoxin, cylindrospermopsin. FEMS Microbiol Lett 2004, 235, 125–129. [Google Scholar]
- Kinnear, S; Duivenvoorden, L; Fabbro, L. Growth and bioconcentration in Spirodela oligorrhiza following exposure to Cylindrospermopsis raciborskii whole cell extracts. Australas J Ecotoxicol 2007, 12, 19–31. [Google Scholar]
- Kinnear, S; Fabbro, L; Duivenvoorden, L. Variable growth responses of water thyme (hydrilla verticillata) to whole-cell extracts of cylindrospermopsis raciborskii. Arch Environ Contam Toxicol 2008, 54, 187–194. [Google Scholar]
- Metcalf, JS; Lindsay, J; Beattie, KA; Birmingham, S; Saker, ML; Törökné, AK; Codd, GA. Toxicity of cylindrospermopsin to the brine shrimp Artemia salina: comparisons with protein synthesis inhibitors and microcystins. Toxicon 2002, 40, 1115–1120. [Google Scholar]
- Humpage, A. Chapter 17: Toxin types, toxicokinetics and toxicodynamics. Cyanobacterial Harmful Algal Blooms State of the Science and Research Needs 2008, 619, 383–416. [Google Scholar]
- Bazin, E; Mourot, A; Humpage, AR; Fessard, V. Genotoxicity of a freshwater cyanotoxin, cylindrospermopsin, in two human cell lines: Caco-2 and HepaRG. Environ Mol Mutagen 2009. [Google Scholar] [CrossRef]
- Pegram, RA; Nichols, T; Etheridge, S; Humpage, A; LeBlanc, S; Love, A; Neilan, B; Pflugmacher, S; Runnegar, M; Thacker, R. Cyanotoxins workgroup report. In Cyanobacterial Harmful Algal Blooms State of the Science and Research Needs; Hudnel, HK, Ed.; Springer Press: New York, NY, USA, 2008; Volume 619, Chapter 15; pp. 317–381. [Google Scholar]
- Young, FM; Micklem, J; Humpage, AR. Effects of blue-green algal toxin cylindrospermopsin (CYN) on human granulosa cells in vitro. Reprod Toxicol 2008, 25, 374–380. [Google Scholar]
- Hudnell, HK. Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer Press: New York, NY, USA, 2008; Volume 619. [Google Scholar]
- Kinnear, SHW; Duivenvoorden, LJ; Fabbro, LD. Ecotoxicity and bioaccumulation of toxin from Cylindrospermopsis raciborskii: towards the development of environmental protection guidelines for contaminated water bodies. In Lake Pollution Research Progress; Miranda, FR, Bernards, LM, Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 81–105. [Google Scholar]
- Saker, ML; Eaglesham, GK. The accumulation of cylindrospermopsin from the cyanobacterium Cylindrospermopsis raciborskii in tissues of the redclaw crayfish Cherax quadricarinatus. Toxicon 1999, 37, 1065–1077. [Google Scholar]
- White, SH; Duivenvoorden, LJ; Fabbro, LD; Eaglesham, GK. Influence of intracellular toxin concentration on cylindrospermopsin bioaccumulation in a freshwater gastropod (Melanoides tuberculata). Toxicon 2006, 47, 497–509. [Google Scholar]
- White, SH; Duivenvoorden, LJ; Fabbro, LD; Eaglesham, GK. Mortality and toxin bioaccumulation in Bufo marinus following exposure to Cylindrospermopsis raciborskii cell extracts and live cultures. Environ Pollut 2007, 147, 158–167. [Google Scholar]
- Saker, ML; Metcalf, JS; Codd, GA; Vasconcelos, VM. Accumulation and depuration of the cyanobacterial toxin cylindrospermopsin in the freshwater mussel Anodonta cygnea. Toxicon 2004, 43, 185–194. [Google Scholar]
- Anderson, L; Fabbro, LD; Cowden, K. Assessment of Blue-Green Algal Toxins in Barramundi. In Red Clay and Mussels from Awoonga Dam; Central Queensland University: Gladstone, Australia, 2003. [Google Scholar]
- White, SH; Duivenvoorden, LJ; Fabbro, LD. Absence of Free-Cylindrospermopsin Bioconcentration in Water Thyme (Hydrilla verticillata). Bull Environ Cont Toxicol 2005, 75, 574–583. [Google Scholar]
- Norris, RL; Seawright, AA; Shaw, GR; Smith, MJ; Chiswell, RK; Moore, MR. Distribution of 14C cylindrospermopsin in vivo in the mouse. Environ Toxicol 2001, 16, 498–505. [Google Scholar]
- Nogueira, ICG; Saker, ML; Pflugmacher, S; Wiegand, C; Vasconcelos, VM. Toxicity of the Cyanobacterium Cylindrospermopsis raciborskii to Daphnia magna. Environ Toxicol 2004, 19, 453–459. [Google Scholar]
- Runnegar, MT; Xie, C; Snider, BB; Wallace, GA; Weinreb, SM; Kuhlenkamp, J. In vitro hepatotoxicity of the cyanobacterial alkaloid cylindrospermopsin and related synthetic analogues. Toxicol Sci 2002, 67, 81–87. [Google Scholar]
- Chong, MWK; Wong, BSF; Lam, PKS; Shaw, GR; Seawright, AA. Toxicity and uptake mechanism of cylindrospermopsin and lophyrotomin in primary rat hepatocytes. Toxicon 2002, 40, 205–211. [Google Scholar]
- Wiegand, C; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol Appl Pharmacol 2005, 203, 201–218. [Google Scholar]
- Meriluoto, J. Cyanotoxins: sampling, sample processing and toxin uptake. In Cyanobacterial harmful Algal Blooms State of the Science and Research Needs; Hudnel, HK, Ed.; Springer Press: New York, NY, USA, 2008 619; Chapter 21; pp. 317–381.
- Runnegar, MT; Kong, SM; Zhong, YZ; Ge, JL; Lu, SC. The role of glutathione in the toxicity of a novel cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem Biophys Res Commun 1994, 201, 235–241. [Google Scholar]
- Beyer, D; Suranyi, G; Vasas, G; Roszik, J; Erdodi, F; M-Hamvas, M; Bacsi, I; Batori, R; Serfozo, Z; Szigeti, ZM; Vereb, G; Demeter, Z; Gonda, S; Mathe, C. Cylindrospermopsin induces alterations of root histology and microtubule organization in common reed (Phragmites australis) plantlets cultured in vitro. Toxicon 2009, 54, 440–449. [Google Scholar]
- Chiswell, RK; Shaw, GR; Eaglesham, G; Smith, MJ; Norris, RI; Seawright, AA; Moore, MR. Stability of cylindrospermopsin, the toxin produced from the cyanobacterium, Cylindrospermopsis raciborskii. Effect of pH, temperature, and sunlight on decomposition. Environ Toxicol 1999, 14, 155–161. [Google Scholar]
- Everson, S; Fabbro, L; Kinnear, S; Eaglesham, G; Wright, P. Distribution of the cyanobacterial toxins cylindrospermopsin and deoxycylindrospermopsin in a stratified lake in north-eastern New South Wales, Australia. Mar Fresh Res 2009, 60, 25–33. [Google Scholar]
- Saker, ML; Griffiths, DJ. Occurrence of blooms of the cyanobacterium Cylindrospermopsis raciborskii (Woloszynska) Seenayya and Subba Raju in a north Queensland domestic water supply. Mar Freshwater Res 2001, 52, 907–915. [Google Scholar]
- Klitzke, S; Apelt, S; Weiler, C; Fastner, J; Chorus, I. Retention and degradation of the cyanobacterial toxin cylindrospermopsin in sediments - The role of sediment preconditioning and DOM composition. Toxicon 2010, in press. [Google Scholar]
- Codd, GA; Metcalf, JS; Beattie, KA. Retention of Microcystis aeruginosa and microcystin by salad lettuce (Lactuca sativa) after spray irrigation with water containing cyanobacteria. Toxicon 1999, 37, 1181–1185. [Google Scholar]
- Peuthert, A; Lawton, L; Pflugmacher, S. In vivo influence of cyanobacterial toxins on enzyme activity and gene expression of protein phosphatases in Alfalfa (Medicago sativa). Toxicon 2008, 52, 84–90. [Google Scholar]
- Saqrane, S; Ouahid, Y; El Ghazali, I; Oudra, B; Bouarab, L; del Campo, FF. Physiological changes in Triticum durum, Zea mays, Pisum sativum and Lens esculenta cultivars, caused by irrigation with water contaminated with microcystins: A laboratory experimental approach. Toxicon 2009, 53, 786–796. [Google Scholar]
- Orr, PT; Rasmussen, JP; Burford, MA; Eaglesham, GK; Lennox, SM. Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 2010, in press. [Google Scholar]
- Fabbro, LD; Baker, M; Duivenvoorden, LJ; Pegg, G; Shiel, R. The Effects of the Ciliate Paramecium cf. caudatum Ehrenberg on Toxin Producing Cylindrospermopsis Isolated from the Fitzroy River, Australia. Environ Toxicol 2001, 16, 489–497. [Google Scholar]
- Gérard, C; Poullain, V; Lance, E; Acou, A; Brient, L; Carpentier, A. Influence of toxic cyanobacteria on community structure and microcystin accumulation of freshwater molluscs. Environ Pollut 2009, 157, 609–617. [Google Scholar]
- Lance, E; Brient, L; Bormans, M; Gerard, C. Interactions between cyanobacteria and Gastropods: I. Ingestion of toxic Planktothrix agardhii by Lymnaea stagnalis and the kinetics of microcystin bioaccumulation and detoxification. Aquat Toxicol 2006, 79, 140–148. [Google Scholar]
- Wiegand, C; Pflugmacher, S. Uptake of Microcystin-LR in Aquatic Organisms. In Cyanotoxins Occurence, Causes, Consequences; Chorus, I, Ed.; Springer-Verlag: Berlin, Germany, 2001; pp. 249–252. [Google Scholar]
- Kotak, BG; Zurawell, R; Prepas, E; Holmes, CF. Microcystin-LR concentration in aquatic food web compartments from lakes of varying trophic status. Can J Fish Aquat Sci 1996, 53, 1974–1985. [Google Scholar]
- Ferrão-Filho, A; Kozlowsky-Suzuki, B; Azevedo, SMFO. Accumulation of microcystins by a tropical zooplankton community. Aquat Toxicol 2002, 59, 201–208. [Google Scholar]
- Saker, ML; Vale, M; Kramer, D; Vasconcelos, VM. Molecular techniques for the early warning of toxic cyanobacteria blooms in freshwater lakes and rivers. App Microbiol Biotech 2007, 75, 441–449. [Google Scholar]
- Froscio, SM; Fanok, S; Humpage, AR. Cytotoxicity screening for the cyanobacterial toxin cylindrospermopsin. J Toxicol Environ Health Part A 2009, 72, 345–349. [Google Scholar]
- Oehrle, SA; Southwell, B; Westrick, J. Detection of various freshwater cyanobacterial toxins using ultra-performance liquid chromatography tandem mass spectrometry. Toxicon 2010, in press. [Google Scholar]
- Wormer, L; Carrasco, D; Cirés, S; Quesada, A. Advances in solid phase extraction of the cyanobacterial toxin cylindrospermopsin. Limnol Oceanogr 2009, 7, 568–575. [Google Scholar]
- Gallo, P; Fabbrocino, S; Cerulo, MG; Ferranti, P; Bruno, M; Serpe, L. Determination of cylindrospermopsin in freshwaters and fish tissue by liquid chromatography coupled to electrospray ion trap mass spectrometry. Rapid Commun Mass Spectrom 2009, 23, 3279–3284. [Google Scholar]
- Kankaanpää, H; Vuorinen, PJ; Sipiä, V; Keinänen, M. Acute effects and bioaccumulation of nodularin in sea trout (Salmo trutta m. trutta L.) exposed orally to Nodularia spumigena under laboratory conditions. Aquat Toxicol 2002, 61, 155–168. [Google Scholar]
- Zhang, D; Xie, P; Chen, J. Effects of Temperature on the Stability of Microcystins in Muscle of Fish and Its Consequences for Food Safety. Environ Cont Toxicol 2010, 84, 202–207. [Google Scholar]
- Neumann, C; Bain, P; Shaw, G. The in vitro toxicology of deoxycylindrospermopsin. Australian Research Network for Algal Toxins Workshop, Moreton Bay Research Station, Australia, , 9–11th July, 2005.
- Smith, JL; Boyer, GL; Zimba, PV. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar]
- Baker, ML. Development of immunoassays for the detection of cylindrospermopsin, produced by Cylindrospermopsis raciborskii. In Master of Applied Science Thesis; Central Queensland University: Rockhampton, Australia, 1998. [Google Scholar]
- Hawkins, PR; Putt, E; Falconer, IR; Humpage, A. Phenotypical variation in a toxic strain of the phytoplankter, Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) during batch culture. Environ Toxicol 2001, 16, 460–467. [Google Scholar]

{kind=link}
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kinnear, S. Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research. Mar. Drugs 2010, 8, 542-564. https://doi.org/10.3390/md8030542
Kinnear S. Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research. Marine Drugs. 2010; 8(3):542-564. https://doi.org/10.3390/md8030542
Chicago/Turabian StyleKinnear, Susan. 2010. "Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research" Marine Drugs 8, no. 3: 542-564. https://doi.org/10.3390/md8030542