Marine Biocatalysts: Enzymatic Features and Applications


Abstract
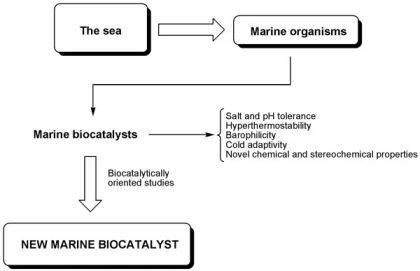
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Marine Biocatalysts
3. Habitat-Related Properties of Marine Biocatalysts
3.1. Salt and pH Tolerance
3.2. Hyperthermostability
3.3. Barophilicity
3.4. Cold Adaptivity
3.5. Novel Chemical and Stereochemical Properties
4. Conclusions
Acknowledgments
References
- Leary, D; Vierros, M; Hamon, G; Arico, S; Monagle, C. Marine genetic resources: a review of scientific and commercial interest. Mar Policy 2009, 33, 183–194. [Google Scholar]
- Trincone, A. Potential biocatalysts originating from sea environments. J Mol Catal B Enzym 2010, 66, 241–256. [Google Scholar]
- Debashish, G; Malay, S; Barindra, S; Joydeep, M. Marine enzymes. Adv Biochem Eng Biotechnol 2005, 96, 189–218. [Google Scholar]
- Antranikian, G; Vorgias, CE; Bertoldo, C. Extreme environments as a resource for microorganisms and novel biocatalysts. Adv Biochem Eng Biotechnol 2005, 96, 219–262. [Google Scholar]
- Demirjian, DC; Morís-Varas, F; Cassidy, CS. Enzymes from extremophiles. Curr Opin Chem Biol 2001, 5, 144–151. [Google Scholar]
- Ferrer, M; Golyshina, O; Beloqui, A; Golyshin, PN. Mining enzymes from extreme environments. Curr Opin Microbiol 2007, 10, 207–214. [Google Scholar]
- Wang, X; Müller, WEG. Marine biominerals: perspectives and challenges for polymetallic nodules and crusts. Trends Biotechnol 2009, 27, 375–383. [Google Scholar]
- Takahashi, K; Fukunaga, K. Conversion of fisheries by-products and waste into value-added products—Attempts undergoing in Hokkaido, Japan. In Biocatalysis and Bioenergy; Hou, CT, Shaw, J-F, Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2008; pp. 417–430. [Google Scholar]
- van der Oost, R; Beyer, J; Vermeulen, NPE. Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ Toxicol Pharmacol 2003, 13, 57–149. [Google Scholar]
- Jorgensen, A; Giessing, AMB; Rasmussen, LJ; Andersen, O. Biotransformation of polycyclic aromatic hydrocarbons in marine polychaetes. Mar Environ Res 2008, 65, 171–186. [Google Scholar]
- Torres, MA; Barros, MP; Campos, SCG; Pinto, E; Rajamani, S; Sayre, RT; Colepicolo, P. Biochemical biomarkers in algae and marine pollution: a review. Ecotoxicol Environ Saf 2008, 71, 1–15. [Google Scholar]
- Sarkar, A; Ray, D; Shrivastava, AN; Sarker, S. Molecular biomarkers: their significance and application in marine pollution monitoring. Ecotoxicology 2006, 15, 333–340. [Google Scholar]
- Apraiz, I; Cristobal, S. Identification of proteomic signatures of exposure to marine pollutants in mussels (Mytilus edulis). Mol Cell Proteomics 2006, 5, 1274–1285. [Google Scholar]
- Donovan, CJ; Garduno, RA; Kalmokoff, M; Ku, JC; Quilliam, MA; Gill, TA. Pseudoalteromonas bacteria are capable of degrading paralytic shellfish toxins. Appl Environ Microbiol 2009, 75, 6919–6923. [Google Scholar]
- Uria, AR; Fawzya, YN; Chasanah, E. Novel molecular methods for discovery and enginnering of biocatalysts from uncultured marine microorganisms. J Coast Dev 2005, 8, 53–71. [Google Scholar]
- Kennedy, J; Marchesi, JR; Dobson, ADW. Marine metagenomics: strategies for the discovery of novel enzymes with biotechnological applications from marine environments. Microb Cell Fact 2008, 7, 27–34. [Google Scholar]
- Hochmuth, T; Piel, J. Polyketide synthases of bacterial symbionts in sponges—Evolution-based applications in natural products research. Phytochemistry 2009, 70, 1841–1849. [Google Scholar]
- Lee, HS; Kwon, KK; Kang, SG; Cha, S-S; Kim, S-J; Lee, J-H. Approaches for novel enzyme discovery from marine environments. Curr Opin Biotechnol 2010, 21, 353–357. [Google Scholar]
- Kennedy, J; Flemer, B; Jackson, SA; Lejon, DPH; Morrissey, JP; O’Gara, F; Dobson, ADW. Marine metagenomics: new tools for the study and exploitation of marine microbial metabolism. Mar Drugs 2010, 8, 608–628. [Google Scholar]
- Sanchez-Amat, A; Solano, F; Lucas-Elío, P. Finding new enzymes from bacterial physiology: a successful approach illustrated by the detection of novel oxidases in Marinomonas mediterranea. Mar Drugs 2010, 8, 519–541. [Google Scholar]
- Sarkar, S; Pramanik, A; Mitra, A; Mukherjee, J. Bioprocessing data for the production of marine enzymes. Mar Drugs 2010, 8, 1323–1372. [Google Scholar]
- Natsumi, S; Martin, R; Kitamura, S; Baran, R; Soga, T; Hirotada, M; Takkaaki, N; Masaru, T. Metabolomics approach for enzyme discovery. J Proteome Res 2006, 5, 1979–1987. [Google Scholar]
- Quillaguamán, J; Guzmán, H; Van-Thuoc, D; Hatti-Kaul, R. Synthesis and production of polyhydroxyalkanoates by halophiles: current potential and future prospects. Appl Microbiol Biotechnol 2010, 85, 1687–1696. [Google Scholar]
- Marhuenda-Egea, FC; Bonete, MJ. Extreme halophilic enzymes in organic solvents. Curr Opin Biotechnol 2002, 13, 385–389. [Google Scholar]
- Lee, M-K; González, JM; Robb, FT. Extremely thermostable glutamate dehydrogenase (GDH) from the freshwater archaeon Thermococcus waiotapuensis: cloning and comparison with two marine hyperthermophilic GDHs. Extremophiles 2002, 6, 151–159. [Google Scholar]
- Matsumoto, M; Yokouchi, H; Suzuki, N; Ohata, H; Matsunaga, T. Saccharification of marine microalgae using marine bacteria for ethanol production. Appl Biochem Biotechnol 2003, (105–108), 247–254. [Google Scholar]
- Chakraborty, S; Khopade, A; Kokare, C; Mahadika, K; Chopade, B. Isolation and characterization of novel α-amylase from marine Streptomyces sp. D1. J Mol Catal B Enzym 2009, 58, 17–23. [Google Scholar]
- Bougatef, A; Balti, R; Nasri, R; Jellouli, K; Soussi, N; Nasri, M. Biochemical properties of anionic trypsin acting at high concentration of NaCl purified from the intestine of a carnivorous fish: smooth hound (Mustelus mustelus). J Agric Food Chem 2010, 58, 5763–5769. [Google Scholar]
- Uo, T; Ueda, M; Nishiyama, T; Yoshimura, T; Esaki, N. Purification and characterization of alanine racemase from hepatopancreas of black-tiger prawn, Penaeus monodon. J Mol Catal B Enzym 2001, 12, 137–144. [Google Scholar]
- Egorova, K; Antranikian, G. Industrial relevance of thermophilic Archaea. Curr Opin Microbiol 2005, 8, 649–655. [Google Scholar]
- Matsuda, T; Nakamura, K. Recent progress in biocatalysis for asymmetric oxidation and reduction. Tetrahedron Asymmetry 2009, 20, 513–557. [Google Scholar]
- Zhu, D; Malik, HT; Hua, L. Asymmetric ketone reduction by a hyperthermophilic alcohol dehydrogenase. The substrate specificity, enantioselectivity and tolerance of organic solvents. Tetrahedron Asymmetry 2006, 17, 3010–3014. [Google Scholar]
- Goyal, K; Selvakumar, P; Hayashi, K. Characterization of a thermostable β-glucosidase (BglB) from Thermotoga maritima showing transglycosylation activity. J Mol Catal B Enzym 2001, 15, 45–53. [Google Scholar]
- Jiang, ZQ; Deng, W; Zhu, YP; Li, LT; Sheng, YJ; Hayashi, K. The recombinant xylanase B of Thermotoga maritima is highly xylan specific and produces exclusively xylobiose from xylans, a unique character for industrial applications. J Mol Catal B Enzym 2004, 27, 207–213. [Google Scholar]
- Tramice, A; Pagnotta, E; Romano, I; Gambacorta, A; Trincone, A. Transglycosylation reactions using glycosyl hydrolases from Thermotoga neapolitana, a marine hydrogen-producing bacterium. J Mol Catal B Enzym 2007, 47, 21–27. [Google Scholar]
- Verenium-Fuelzyme®. Fuelzyme® enzyme is a next generation alpha amylase for starch liquefaction. Available online: http://www.verenium.com/products_fuelzyme.html (accessed on 23 March 2011).
- Bartlett, DH. Microbial life at high pressures. Sci Prog (Oxf) 1992, 76, 479–496. [Google Scholar]
- Abe, F; Horikoshi, K. The biotechnological potential of piezophiles. Trends Biotechnol 2001, 19, 102–108. [Google Scholar]
- Horikoshi, K. Barophiles: deep-sea microorganisms adapted to an extreme environment. Curr Opin Microbiol 1998, 1, 291–295. [Google Scholar]
- Hay, S; Evans, RM; Levy, C; Loveridge, EJ; Wang, X; Leys, D; Allemann, RK; Scrutton, NS. Are the catalytic properties of enzymes from piezophilic organisms pressure adapted. ChemBioChem 2009, 10, 2348–2353. [Google Scholar]
- Canganella, F; Gonzalez, JM; Yanagibayashi, M; Kato, C; Horikoshi, K. Pressure and temperature effects on growth and viability of the hyperthermophilic archaeon Thermococcus peptonophilus. Arch Microbiol 1997, 168, 1–7. [Google Scholar]
- Nakayama, A; Yano, Y; Yoshida, K. New method for the isolation of barophiles from intestinal contents of deep-sea fishes retrieved from the abyssal zone. Appl Environ Microbiol 1994, 60, 4210–4212. [Google Scholar]
- Fang, J; Kato, C; Sato, T; Chan, O; McKay, D. Biosynthesis and dietary uptake of polyunsaturated fatty acids by piezophilic bacteria. Comp Biochem Physiol B 2004. [Google Scholar]
- Daniel, I; Oger, P; Winter, R. Origins of life and biochemistry under high-pressure conditions. Chem Soc Rev 2006, 35, 858–875. [Google Scholar]
- Lo Giudice, A; Michaud, L; de Pascale, D; De Domenico, M; di Prisco, G; Fani, R; Bruni, V. Lipolytic activity of Antarctic cold-adapted marine bacteria (Terra Nova Bay, Ross Sea). J Appl Microbiol 2006, 101, 1039–1048. [Google Scholar]
- Gerday, C; Aittaleb, M; Bentahir, M; Chessa, JP; Claverie, P; Collins, T; D’Amico, S; Dumont, J; Garsoux, G; Georlette, D; Hoyoux, A; Lonhienne, T; Meuwis, MA; Feller, G. Cold-adapted enzymes: from fundamentals to biotechnology. Trends Biotechnol 2000, 18, 103–107. [Google Scholar]
- Cristóbal, HA; Schmidt, A; Kothe, E; Breccia, J; Abate, CM. Characterization of inducible cold-active β-glucosidases from the psychrotolerant bacterium Shewanella sp. G5 isolated from a sub-Antarctic ecosystem. Enzym Microb Technol 2009, 45, 498–506. [Google Scholar]
- Khudary, RA; Venkatachalam, R; Katzer, M; Elleuche, S; Antranikian, G. A cold-adapted esterase of a novel marine isolate, Pseudoalteromonas arctica: gene cloning, enzyme purification and characterization. Extremophiles 2010, 14, 273–285. [Google Scholar]
- Kim, HS; Lee, OK; Hwang, S; Kim, BJ; Lee, EY. Biosynthesis of (R)-phenyl-1,2-ethanediol from racemic styrene oxide by using bacterial and marine fish epoxide hydrolases. Biotechnol Lett 2008, 30, 127–133. [Google Scholar]
- Kim, HS; Lee, OK; Lee, SJ; Hwang, S; Kim, SJ; Yang, SH; Park, S; Lee, EY. Enantioselective epoxide hydrolase activity of a newly isolated microorganism, Sphingomonas echinoides EH-983, from seawater. J Mol Catal B Enzym 2006, 41, 130–135. [Google Scholar]
- Kim, HS; Lee, SJ; Lee, EJ; Hwang, JW; Park, S; Kim, SJ; Lee, EY. Cloning and characterization of a fish microsomal epoxide hydrolase of Danio rerio and application to kinetic resolution of racemic styrene oxide. J Mol Catal B Enzym 2005, 37, 30–35. [Google Scholar]
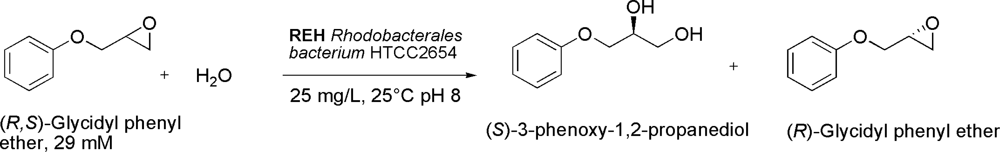
- Woo, J-H; Kang, J-H; Hwang, Y-O; Cho, J-C; Kim, S-J; Kang, SG. Biocatalytic resolution of glycidyl phenyl ether using a novel epoxide hydrolase from a marine bacterium, Rhodobacterales bacterium HTCC2654. J. Biosci. Bioeng 2010, 109, 539–544. [Google Scholar]
- Jorgensen, A; Giessing, AMB; Rasmussen, LJ; Andersen, O. Biotransformation of polycyclic aromatic hydrocarbons in marine polychaetes. Mar Environ Res 2008, 65, 171–186. [Google Scholar]
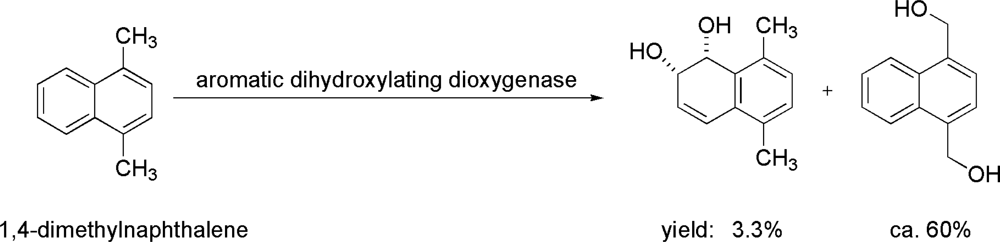
- Shindo, K; Osawa, A; Kasai, Y; Iba, N; Saotome, A; Misawa, N. Hydroxylations of substituted naphthalenes by Escherichia coli expressing aromatic dihydroxylating dioxygenase genes from polycyclic aromatic hydrocarbon-utilizing marine bacteria. J Mol Catal B Enzym 2007, 48, 77–83. [Google Scholar]
- Wasmund, K; Burns, KA; Kurtboke, DI; Boume, DG. Novel alkane hydroxylase (alkB) gene diversity in sediments associated with hydrocarbon seeps in the Timor Sea, Australia. Appl Environ Microbiol 2009, 75, 7391–7398. [Google Scholar]
- Miri, M; Bambai, B; Tabandeh, F; Sadeghizadeh, M; Kamali, N. Production of a recombinant alkane hydroxylase (AlkB2) from Alcanivorax borkumensis. Biotechnol Lett 2010, 32, 497–502. [Google Scholar]
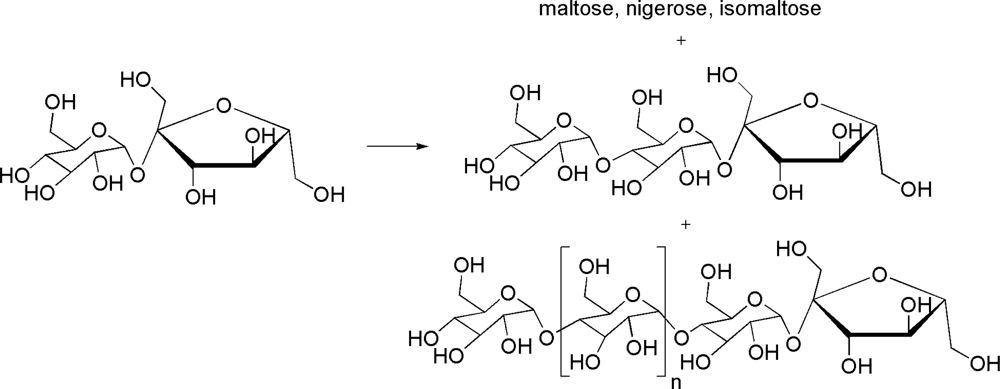
- Giordano, A; Andreotti, G; Mollo, E; Trincone, A. Transglycosylation reactions performed by glycosyl hydrolases from the marine anaspidean mollusc Aplysia fasciata. J Mol Catal B Enzym 2004, 30, 51–59. [Google Scholar]
- Andreotti, G; Giordano, A; Tramice, A; Mollo, E; Trincone, A. Purification and characterization of a β-d-mannosidase from the marine anaspidean Aplysia fasciata. J Biotechnol 2005, 119, 26–35. [Google Scholar]
- Andreotti, G; Giordano, A; Tramice, A; Mollo, E; Trincone, A. Hydrolyses and transglycosylations performed by purified α-d-glucosidase of the marine mollusc Aplysia fasciata. J Biotechnol 2006, 122, 274–284. [Google Scholar]
- Giordano, A; Tramice, A; Andreotti, G; Mollo, E; Trincone, A. Enzymatic syntheses and selective hydrolysis of O-β-d-galactopyranosides using a marine mollusc β-galactosidase. Bioorg Med Chem Lett 2005, 15, 139–143. [Google Scholar]
- Andreotti, G; Trincone, A; Giordano, A. Convenient synthesis of β-galactosyl nucleosides using the marine β-galactosidase from Aplysia fasciata. J Mol Catal B Enzym 2007, 47, 28–32. [Google Scholar]
- Tramice, A; Andreotti, G; Trincone, A. High-yielding enzymatic α-glucosylation of pyridoxine by marine α-glucosidase from Aplysia fasciata. Mar Biotechnol 2006, 8, 448–452. [Google Scholar]
- Tramice, A; Andreotti, G; Trincone, A. Direct enzymatic glucosylation of naringin in grapefruit juice by α-d-glucosidase from the marine mollusc Aplysia fasciata. Biotechnol J 2008, 3, 545–554. [Google Scholar]
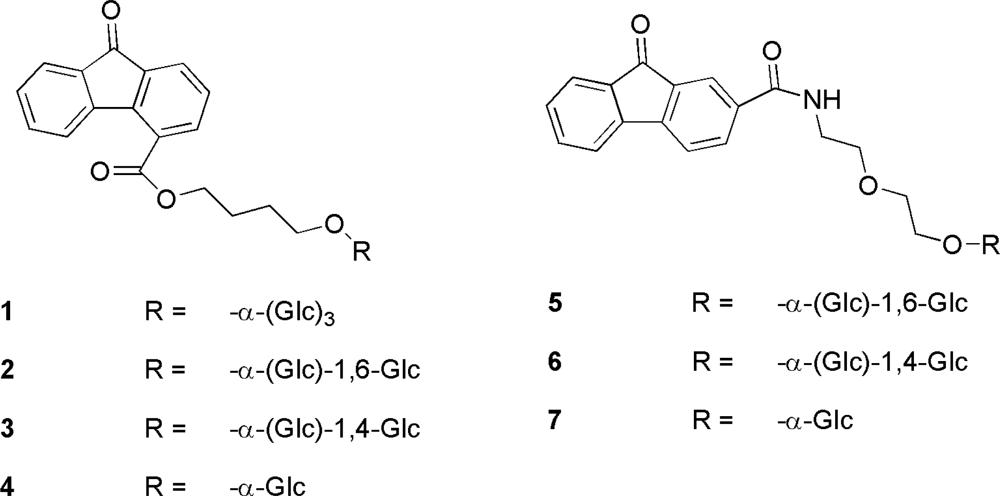
- Arena, A; Arena, N; de Gregorio, A; Maccari, R; Ottanà, R; Pavone, B; Tramice, A; Trincone, A; Vigorita, MG. 2/4-Substituted-9-fluorenones and their O-glucosides as potential immunomodulators and anti-herpes simplex virus-2 agents. Part 5. Eur J Med Chem 2008, 43, 2656–2664. [Google Scholar]
- Hung, VS; Hatada, Y; Goda, S; Yuko Hidaka, JL; Li, Z; Akita, M; Ohta, Y; Watanabe, K; Matsui, H; Ito, S; Horikoshi, K. α-Glucosidase from a strain of deep-sea Geobacillus: a potential enzyme for the biosynthesis of complex carbohydrates. Appl Microbiol Biotechnol 2005, 68, 757–765. [Google Scholar]
- Berteau, O; McCort, I; Goasdonuè, N; Tissot, B; Daniel, R. Characterization of a new α-l-fucosidase isolated from the marine mollusk Pecten maximus that catalyzes the hydrolysis of α-l-fucose from algal fucoidan (Ascophyllum nodosum). Glycobiology 2002, 4, 273–282. [Google Scholar]
- Berteau, O; Bielicki, J; Kilonda, A; Machy, D; Anson, DS; Kenne, L. α-l-Fucosidases: exoglycosidases with unusual transglycosylation properties. Biochemistry 2004, 22, 7881–7891. [Google Scholar]
- Soro, RY; Diopoh, JK; Willemot, R-M; Combes, D. Enzymatic synthesis of polyglucosylfructosides from sucrose alone by a novel α-glucosidase isolated from the digestive juice of Archachatina ventricosa (Achatinideae). Enzym Microb Technol 2007, 42, 44–51. [Google Scholar]
- Long, M; Yu, Z; Xu, X. A novel β-Agarase with high pH stability from marine Agarivorans sp. LQ48. Mar Biotechnol 2010, 12, 62–69. [Google Scholar]
- Zahura, UA; Rahman, MM; Inoue, A; Tanaka, H; Ojima, T. An endo-β-1,4-mannanase, AkMan, from the common sea hare Aplysia kurodai. Comp Biochem Physiol B 2010, 157, 137–143. [Google Scholar]
- Rodrigues, MV; Borges, N; Almeida, CP; Lamosa, P; Santos, H. A unique β-1,2-mannosyltransferase of Thermotoga maritima that uses di-myo-inositol phosphate as the mannosyl acceptor. J Bacteriol 2009, 191, 6105–6115. [Google Scholar]
- Kim, J-T; Kang, SG; Woo, J-H; Lee, J-H; Jeong, BC; Kim, S-J. Screening and its potential application of lipolytic activity from a marine environment: characterization of a novel esterase from Yarrowia lipolytica CL180. Appl Microbiol Biotechnol 2007, 74, 820–828. [Google Scholar]
- Lee, J; McIntosh, J; Hathaway, BJ; Schmidt, EW. Using marine natural products to discover a protease that catalyzes peptide macrocyclization of diverse substrates. J Am Chem Soc 2009, 131, 2122–2124. [Google Scholar]
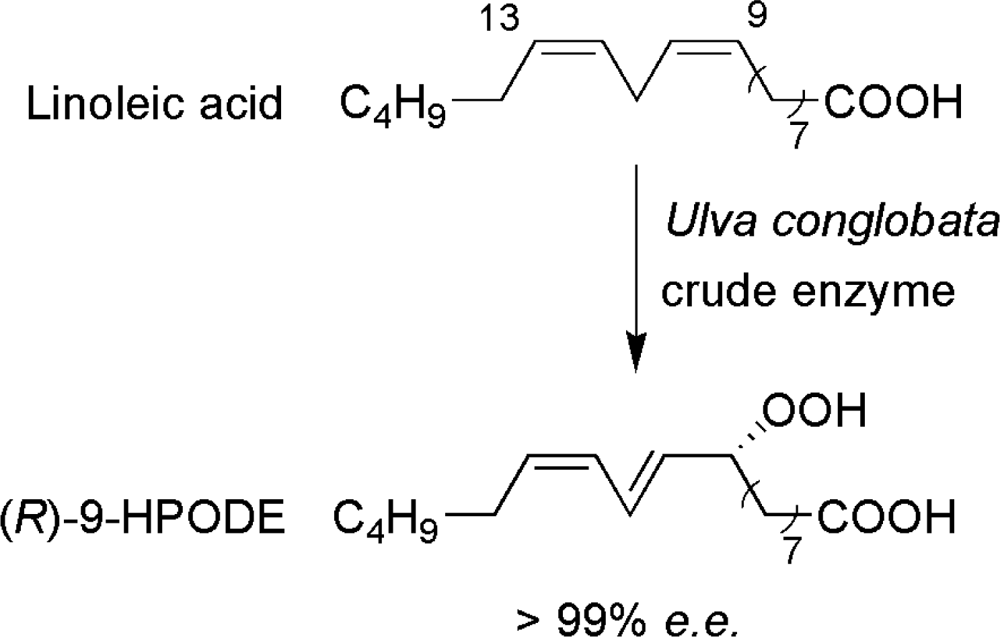
- Akakabe, Y; Matsui, K; Kajiwara, T. Enantioselective formation of (R)-9-HPODE and (R)-9-HPOTrE in marine green alga Ulva conglobata. Bioorg Med Chem Lett 2002, 10, 3171–3173. [Google Scholar]
- Brandao, PFB; Bull, AT. Nitrile hydrolysing activities of deep-sea and terrestrial mycolate actinomycetes. Antonie Van Leeuwenhoek 2003, 84, 89–98. [Google Scholar]
- James, MO. Steroid catabolism in marine and freshwater fish. J Steroid Biochem Mol Biol 2010. [Google Scholar] [CrossRef]






© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Trincone, A. Marine Biocatalysts: Enzymatic Features and Applications. Mar. Drugs 2011, 9, 478-499. https://doi.org/10.3390/md9040478
Trincone A. Marine Biocatalysts: Enzymatic Features and Applications. Marine Drugs. 2011; 9(4):478-499. https://doi.org/10.3390/md9040478
Chicago/Turabian StyleTrincone, Antonio. 2011. "Marine Biocatalysts: Enzymatic Features and Applications" Marine Drugs 9, no. 4: 478-499. https://doi.org/10.3390/md9040478