Potential Changes in Rat Spermatogenesis and Sperm Parameters after Inhalation of Boswellia papyrifera and Boswellia carterii Incense



Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Incense
2.2. Exposure to B. papyrifera and B. carterii Smoke
2.3. Sperm Analysis
2.4. Histopathological and Ultrastructural Study
2.5. Statistical Analysis
3. Results
3.1. Sperm Analysis
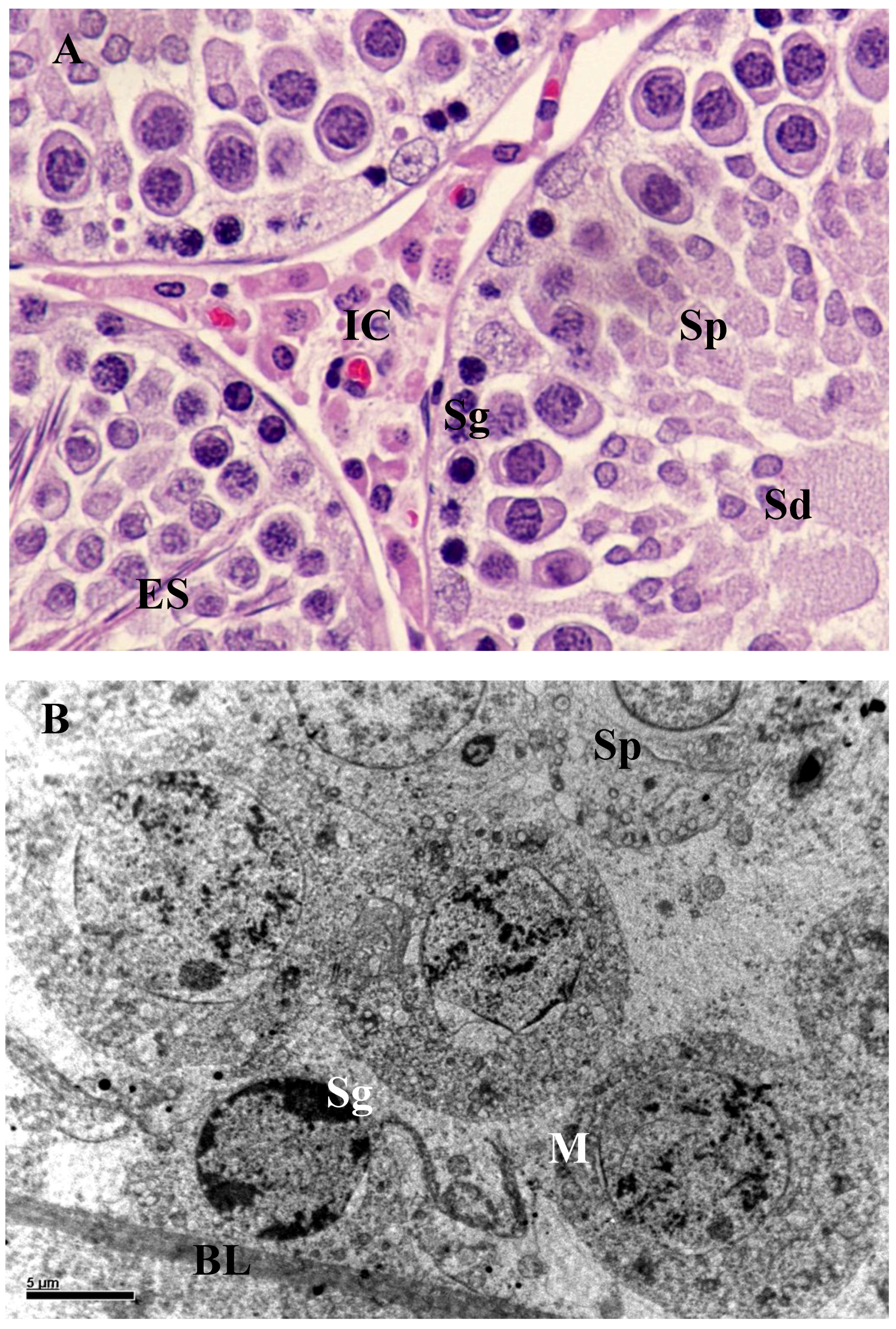
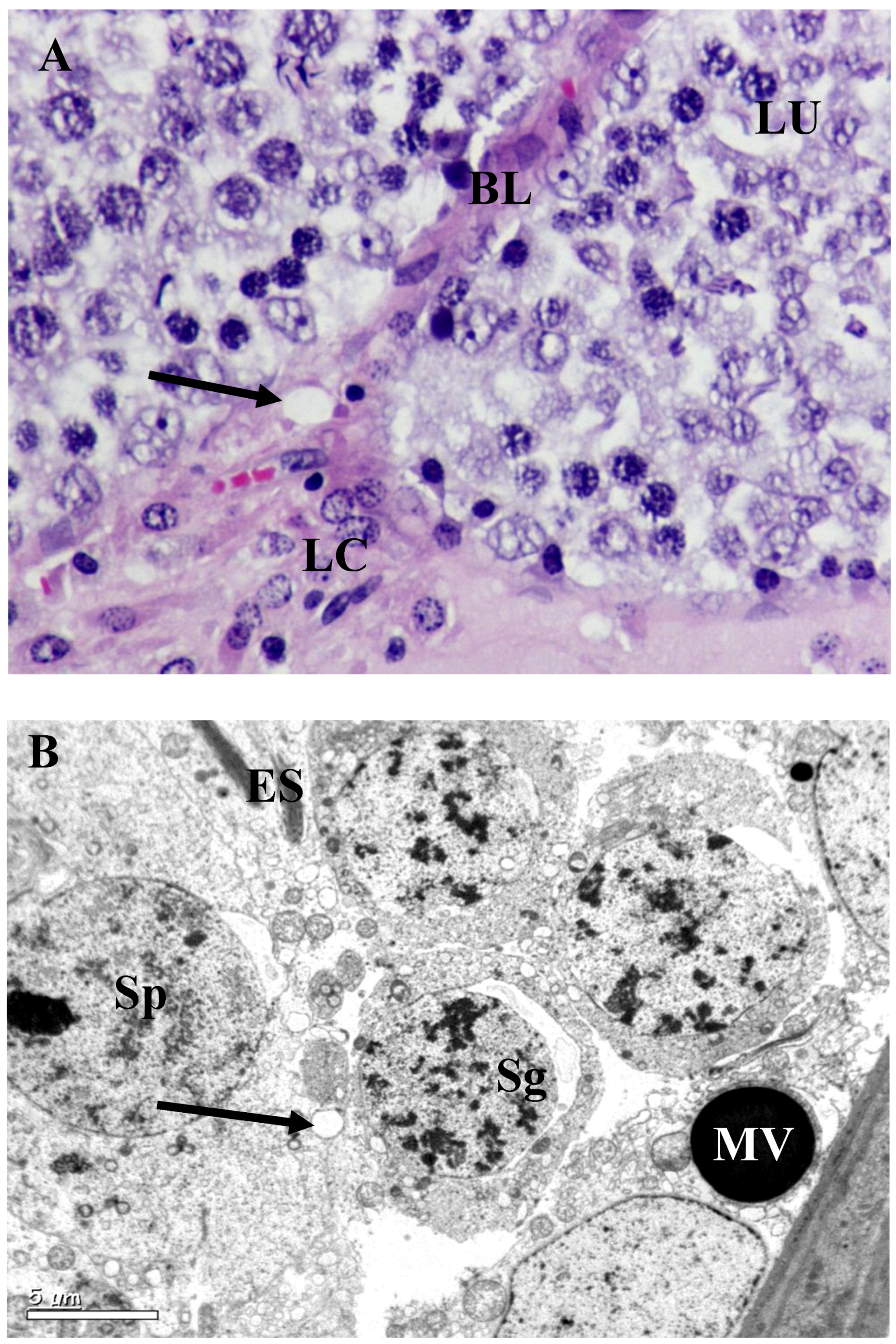
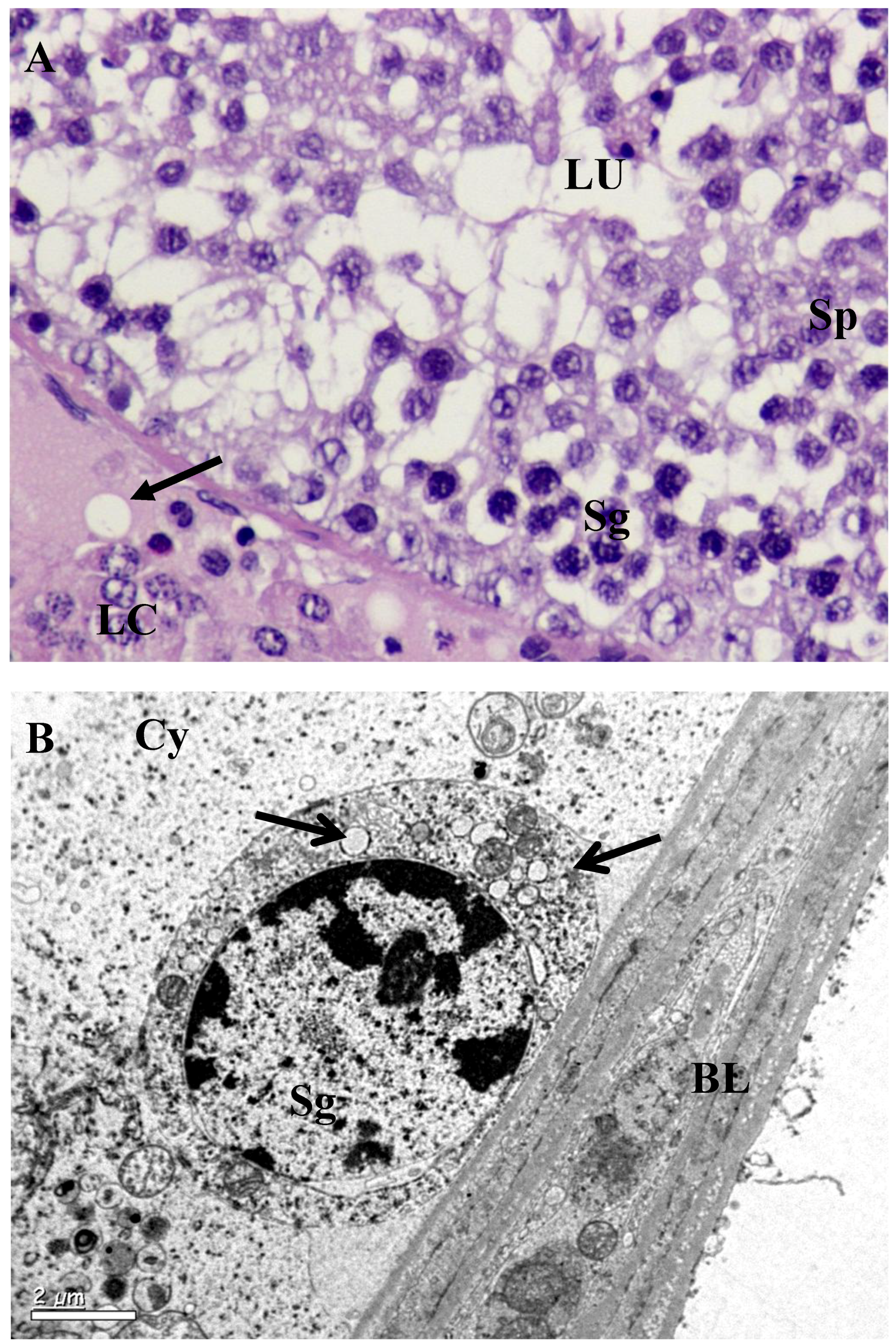
3.2. Histology and Morphometric Study
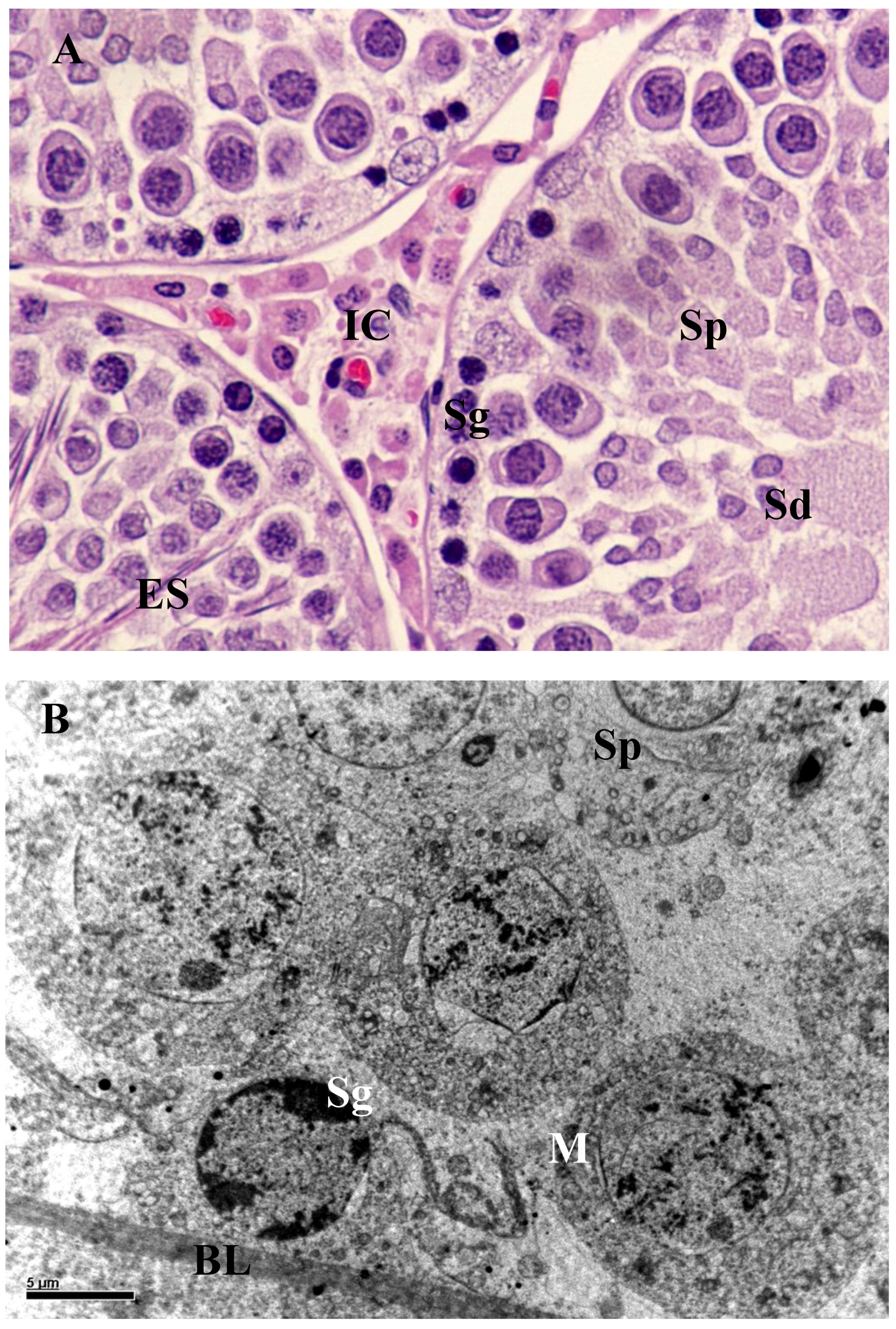
3.3. Ultrastructural Changes in Testis

{kind=link}
{kind=link}
{kind=link}
| Group | Sperm parameters | Plasma fructose (mg/100 mL) | ||||
|---|---|---|---|---|---|---|
| Total number of sperm (Total No. × 104/mL) | Total number of motile sperm (Total No. × 104/mL) | Velocity (µm/s) | Abnormal (%) | Cauda Epididymis | Seminal Vesicle | |
| I | 67.49 ± 1.29 | 55.39 ± 2.17 | 121.56 ± 1.70 | 11.80 ± 1.21 | 80.72 ± 2.13 | 110.58 ± 1.63 |
| Control | ||||||
| II | 37.23 ± 1.31 *** | 33.10 ± 1.16 *** | 65.48 ± 1.51 *** | 55.63 ± 1.47 *** | 39.15 ± 1.91 *** | 47.59 ± 1.37 *** |
| B. papyrifera | ||||||
| III | 31.47 ± 1.11 *** | 27.70 ± 1.31 *** | 61.86 ± 1.29 *** | 59.58 ± 0.34 *** | 29.64 ± 0.32 *** | 43.71 ± 1.67 *** |
| B. carterii | ||||||
| Group | Number of | |||||
|---|---|---|---|---|---|---|
| Seminiferous tubules in microscopic field (10×) | Spermatogonia | Spermatocytes | Spermatids | Leydig cells | Sertoli cells | |
| I | 15.50 ± 0.21 | 130.70 ± 2.39 | 549.55 ± 29.34 | 986.35 ± 7.28 | 57.30 ± 0.41 | 29.63 ± 1.56 |
| Control | ||||||
| II | 21.13 ± 0.41 *** | 90.50 ± 1.36 *** | 427.31 ± 1.41 *** | 762.30 ± 1.43 *** | 27.12 ± 1.35 *** | 20.81 ± 1.67 *** |
| B. papyrifera | ||||||
| III | 23.18 ± 0.40 *** | 70.09 ± 2.36 *** | 321.57 ± 4.21 *** | 593.17 ± 3.3 *** | 25.17 ± 1.77 *** | 17.91 ± 1.39 *** |
| B. carterii | ||||||
| Group | 10× | 100× | ||
|---|---|---|---|---|
| Seminiferous tubules (µm) | Spermatogonia (µm) | Spermatocytes (µm) | Spermatids (µm) | |
| I | 273.51 ± 1.27 | 11.18 ± 0.24 | 9.33 ± 0.31 | 9.00 ± 0.23 |
| Control | ||||
| II | 239.19 ± 5.43 *** | 7.21 ± 0.63 *** | 6.00 ± 0.27 *** | 5.90 ± 0.33 *** |
| B. papyrifera | ||||
| III | 261.16 ± 2.73 *** | 5.21 ± 0.39 *** | 5.39 ± 0.21 *** | 4.56 ± 0.39 *** |
| B. carterii | ||||
| Group | 100× (µm) | |||
|---|---|---|---|---|
| Spermatogonia | Spermatocytes | Spermatids | Leydig cells | |
| I | 10.38 ± 0.19 | 8.45 ± 0.22 | 7.08 ± 0.20 | 9.74 ± 0.27 |
| Control | ||||
| II | 5.09 ± 0.31 *** | 5.63 ± 0.35 *** | 4.26 ± 0.17 *** | 5.61 ± 0.21 *** |
| B. papyrifera | ||||
| III | 4.34 ± 0.27 *** | 4.28 ± 0.21 *** | 3.83 ± 0.11 *** | 4.00 ± 0.19 *** |
| B. carterii | ||||
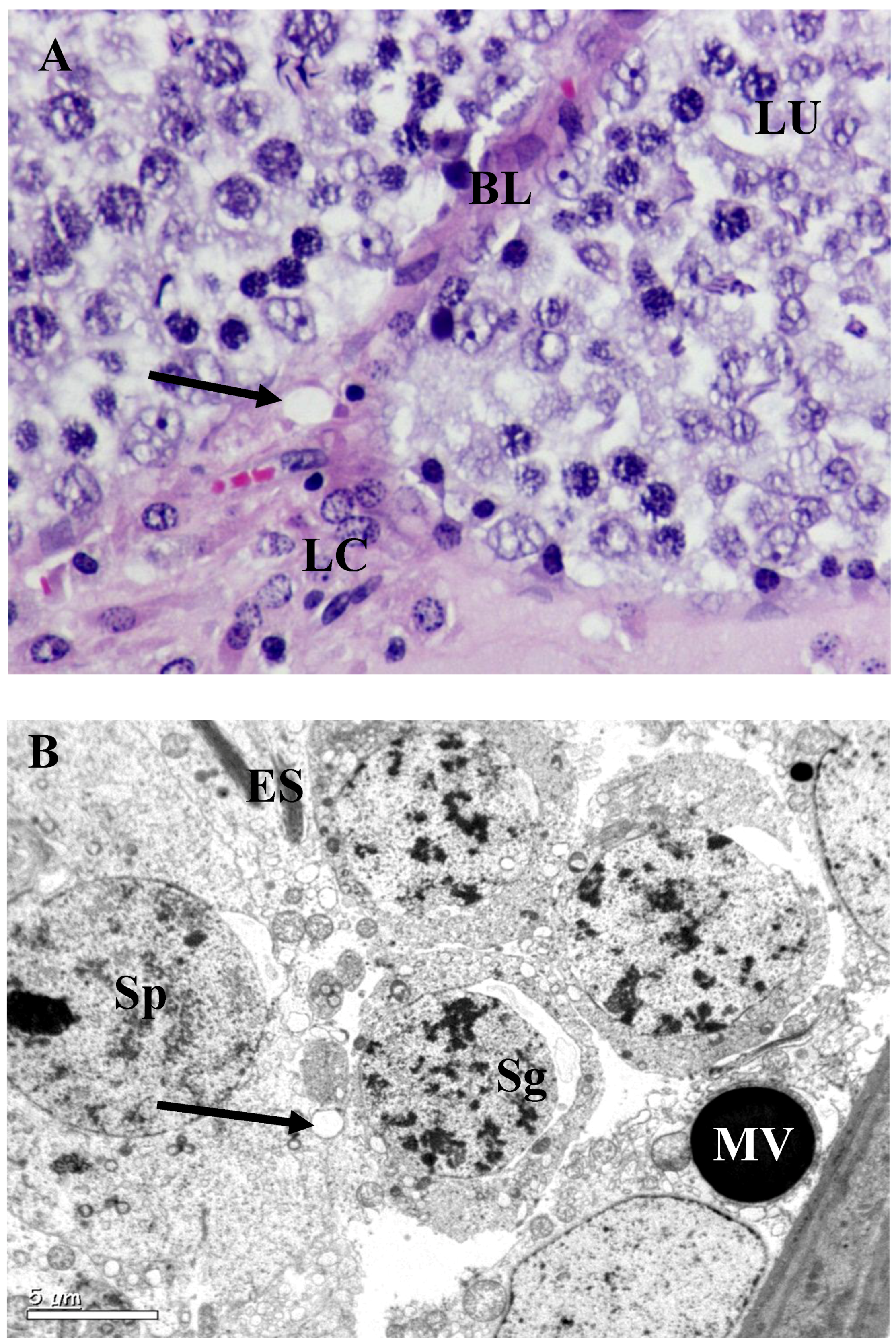
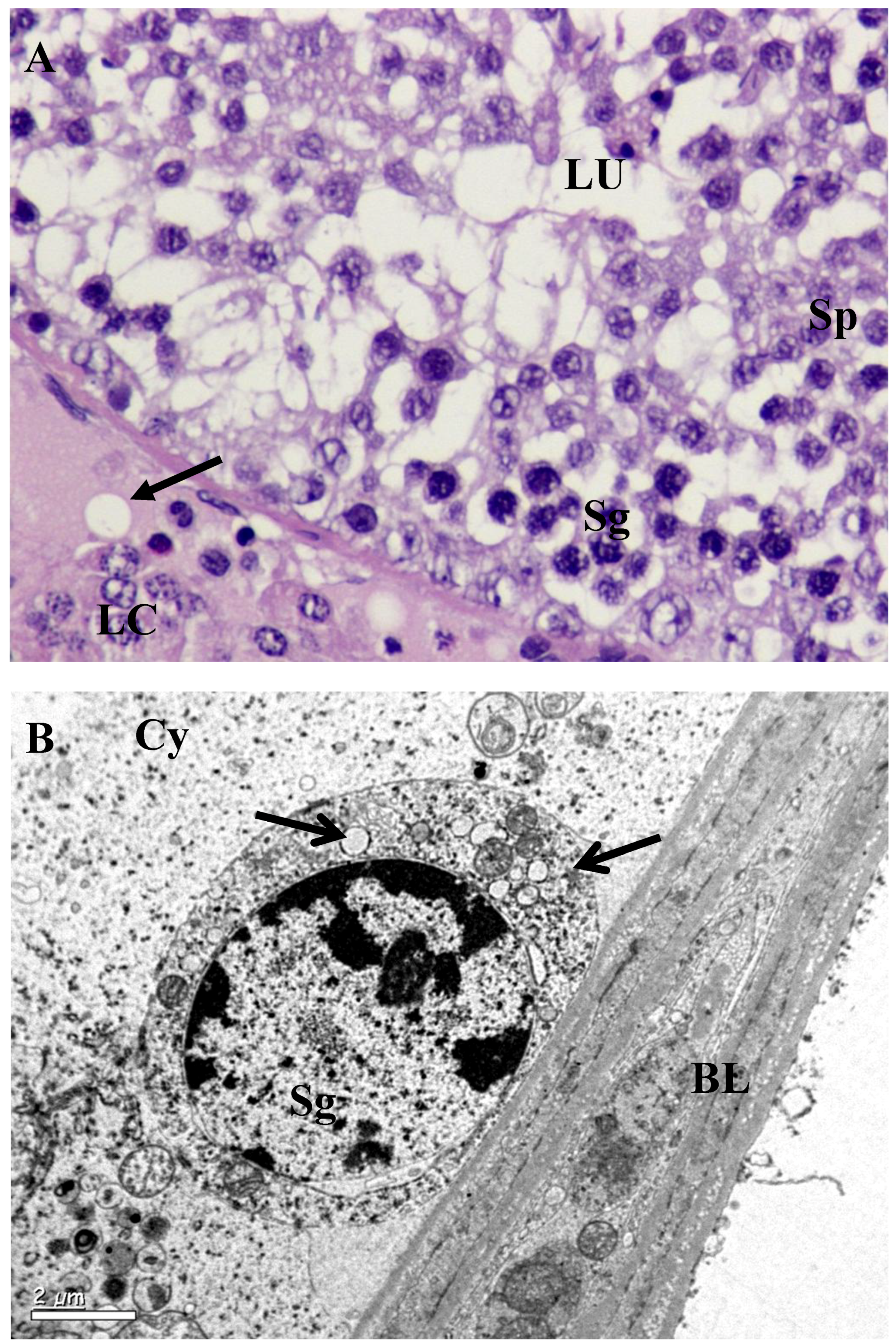
4. Discussion
4.1. Sperm Analysis
4.2. Histology of Testis
4.3. Ultrastructure of Testis
5. Conclusions
Acknowledgments
Conflict of interest
References
- Kulkani, R.R.; Patki, P.S.; Jog, V.P.; Gandage, S.G.; Patwardhan, B. Treatment of osteoarthritis with a herbomineral formulation. A double-blind, placebo-controlled, cross-over study. J. Ethnopharmacol. 1991, 33, 91–95. [Google Scholar] [CrossRef]
- Camarda, L.; Dayton, T.; Di Stefano, V.; Pitonzo, R.; Schillaci, D. Chemical composition and antimicrobial activity of some oleogum resin essential oils from Boswellia spp. (Burseraceae). Ann. Chim. 2007, 97, 837–844. [Google Scholar] [CrossRef]
- Huan, M.T.; Badmaev, V.; Ding, Y.; Liu, Y.; Xie, J.G.; Ho, C.T. Anti-tumor and anti-carcinogenic effects of triterpenoid, beta-boswellic-acid. Biofactors 2000, 13, 225–230. [Google Scholar] [CrossRef]
- Al-Arafi, S.A.; Mubarak, M.; Alokail, M.S. Ultrastructure of the pulmonary alveolar cells of rats exposed to Arabian mix incense (Ma’ amoul). J Biol Sci. 2004, 4, 694–699. [Google Scholar] [CrossRef]
- Alokail, M.S.; Alarifi, S.A. Histological changes in the lung of Wistar albino rats (Rattus norvegicus) after exposure to Arabian incense (Genus Boswellia). Ann. Saudi. med. 2004, 24, 293–295. [Google Scholar]
- Alokail, M.S.; Mohammad, A.I.; Al-Arafi, S.A. Antioxidant enzyme activity and lipid peroxidation in liver of wistar rats exposed to Arabian incense. Animal. Bio. J. 2011, 2, 1–9. [Google Scholar]
- Alokail, M.S.; Al-Daghri, N.M.; Al-Arafi, S.A.; Draz, H.M.; Tajamul, H.; Yakout, S.M. Long-term exposure to incense smoke alters metabolism in Wistar albino rats. Cell Bio. Fun. 2011, 28, 1–6. [Google Scholar]
- Wang, X.D.; Liu, C.; Bronson, R.T.; Smith, D.E.; Krinsky, N.I.; Russell, M.L. Retinoid signaling and activator protein-1 expression in ferrets given beta-carotene supplements and exposed to tobacco smoke. J. Nat. Can. Inst. 1999, 91, 60–66. [Google Scholar] [CrossRef]
- Besley, M.A.; Eliarson, R.; Gallegosm, A.J.; Moghissi, K.S.; Paulsen, C.A.; Prasad, M.R.N. Laboratory Manual for the Examination of Human Semen and Semen Cervical Mucus Interaction; WHO Press concern: Singapore, 1980. [Google Scholar]
- Mukhtar, A.; Nazeer, A.R.; Ravindranath, H.A.; Mukhtar, A.M.G. Effect of benzene extract of Ocimum sanctum leaves on cauda epididymal spermatozoa of rats. Iranian J. Repro. Med. 2011, 3, 177–186. [Google Scholar]
- Ratnasooriya, W.D. Effect of Atropine on fertility of female rat and sperm motility. Indian J. Exp. Boil. 1984, 22, 463–466. [Google Scholar]
- Bauer, J.D.; Ackermen, P.G.; Toro, G. Clinical Laboratory Methods; The C. V. Mosby Company: Saint Louis, MO, USA, 1974. [Google Scholar]
- Reyenolds, E.S. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef]
- Chauhan, A.; Agarwal, M. Reversible changes in the antifertility induced by Aegle marmelos in male albino rats. Sys. Biol. Repro. Med. 2008, 54, 240–246. [Google Scholar] [CrossRef]
- Mann, T. Fructose, Polyols, and Organic Acids. In The Biochemistry of Semen and of the Male Reproductive Tract; Nam, T., Ed.; Methuen: London, UK, 1964; pp. 237–264. [Google Scholar]
- Patel, S.M.; Skandhan, K.P.; Mehta, Y.B. Seminal plasma fructose and glucose in normal and pathological conditions. Acta Eur. Fertil. 1988 19, 329–332.
- Mylonas, C.; Kouretas, D. Lipid peroxidation and tissue damage. In Vivo 1999, 13, 295–309. [Google Scholar]
- Yamamoto, Y.; Isoyama, E.; Sofikitis, N.; Miyagawa, I. Effects of smoking on testicular function and fertilizing potential in rats. Urol. Res. 1998, 26, 45–48. [Google Scholar] [CrossRef]
- Kapawa, A.; Giannakis, D.; Tsoukanelis, K.; Kanakas, N.; Baltogiannis, D.; Agapitos, E.; Loutradis, D.; Miyagawa, I.; Sofikitis, N. Effects of paternal cigarette smoking on testicular function, sperm fertilizing capacity, embryonic development, and blastocyst capacity for implantation in rats. Andrologia 2004, 36, 57–68. [Google Scholar] [CrossRef]
- Beardsley, A.; O’Donnell, L. Characterization of normal spermiation and spermiation failure induced by hormone suppression in adult rats. Biol. Repro. 2003, 68, 1299–1307. [Google Scholar] [CrossRef]
- Aladakatti, R.H.; Nazeer, A.R. Azadirachta indica A. Juss induced changes in spermatogenic pattern in albino rats. J. Nat. Remd. 2006, 6, 62–72. [Google Scholar]
- Aladakatti, R.H.; Mukhtar, A.; Nazeer, A.R.; Ghodesawar, M.G. Effect of benzene leaf extract of Ocimum sanctum on testis and spermatogenic pattern in albino rats. Int. J. Curr. Res. 2010, 5, 22–29. [Google Scholar]
- Yang, Z.W.; Kong, L.S.; Guo, Y.; Yin, J.Q.; Mills, N. Histological changes of the testis and epididymis in adult rats as a result of Leydig cell destruction after ethane dimethane sulfonate treatment: A morphometric study. Asian J. Androl. 2006, 8, 289–299. [Google Scholar] [CrossRef]
- Aladakatti, R.H.; Nazeer, A.R. Changes in Sertoli cells induced by Azadirachta indica A. Juss leaves in albino rats. J. B. Clin. Physiol. Pharmacol. 2005, 16, 67–80. [Google Scholar]
- Aladakatti, R.H.; Nazeer, A.R. Ultrastructural changes in Leydig cell and cauda epididymal spermatozoa induced by Azadirachta indica leaves in albino rats. Phyto. Res. 2005, 19, 756–766. [Google Scholar] [CrossRef]
- Russell, L.D.; Griswold, M.D. The Sertoli Cell. Clearwater; Coche River Press: Saint Louis, MO, USA, 1993. [Google Scholar]
- Lohiya, N.K.; Mishra, P.K.; Pathak, N.; Manivannan, B.; Bhande, S.S.; Panneerdoss, S.; Sriram, S. Efficacy trial on the purified compounds of the seeds of Carica papaya for male contraception in albino rat. Reprod. Toxicol. 2005, 20, 135–148. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ahmed, M.; Al-Daghri, N.; Alokail, M.S.; Hussain, T. Potential Changes in Rat Spermatogenesis and Sperm Parameters after Inhalation of Boswellia papyrifera and Boswellia carterii Incense. Int. J. Environ. Res. Public Health 2013, 10, 830-844. https://doi.org/10.3390/ijerph10030830
Ahmed M, Al-Daghri N, Alokail MS, Hussain T. Potential Changes in Rat Spermatogenesis and Sperm Parameters after Inhalation of Boswellia papyrifera and Boswellia carterii Incense. International Journal of Environmental Research and Public Health. 2013; 10(3):830-844. https://doi.org/10.3390/ijerph10030830
Chicago/Turabian StyleAhmed, Mukhtar, Nasser Al-Daghri, Majed S. Alokail, and Tajamul Hussain. 2013. "Potential Changes in Rat Spermatogenesis and Sperm Parameters after Inhalation of Boswellia papyrifera and Boswellia carterii Incense" International Journal of Environmental Research and Public Health 10, no. 3: 830-844. https://doi.org/10.3390/ijerph10030830