Acoustic Noise Alters Selective Attention Processes as Indicated by Direct Current (DC) Brain Potential Changes

Abstract
:
1. Introduction
2. Material and Methods
2.1. Participants
2.2. Design
2.3. DC Recordings
2.4. Experimental Conditions
2.4.1. Noise

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leq dB (A) | max dB (A) | |
|---|---|---|
| Irrelevant Speech | ||
| Fast | 61.5 | 71.6 |
| Slow | 61.6 | 66.8 |
| White Noise | ||
| Fast | 68.7 | 69.5 |
| Slow | 68.8 | 69.0 |
| Control | ||
| Fast | 38.2 | 48.1 |
| Slow | 37.6 | 41.8 |
2.4.2. Attention Tasks
Intake Verbal—“Listening Task”
Intake Figural—“Recognizing Photographs of Trees”
Reject Verbal—“Creating Words”
Reject Figural—“Picture Completion”
2.5. Procedure
2.6. Analysis
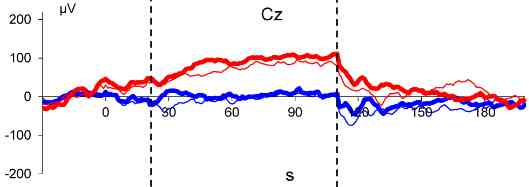
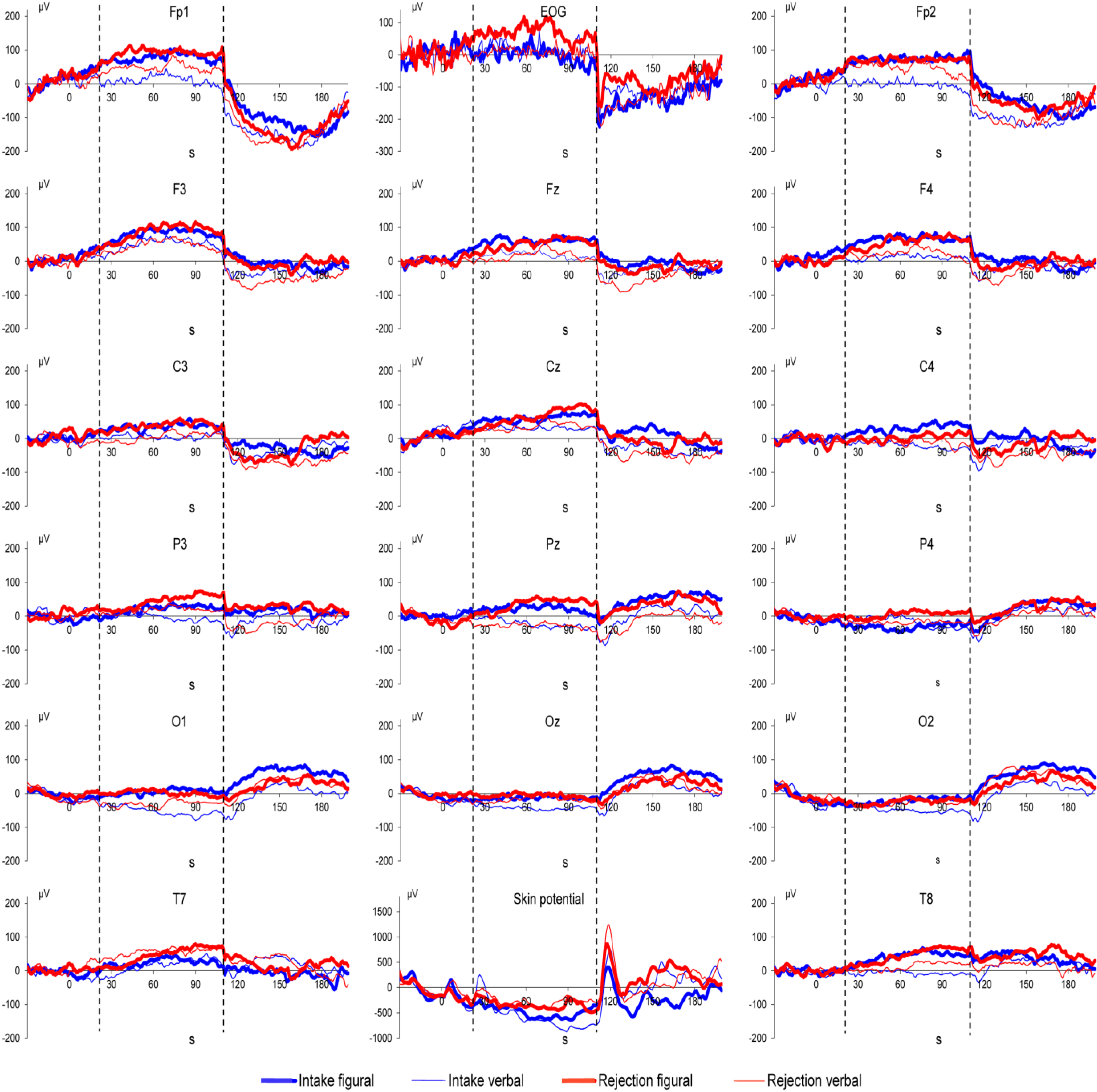
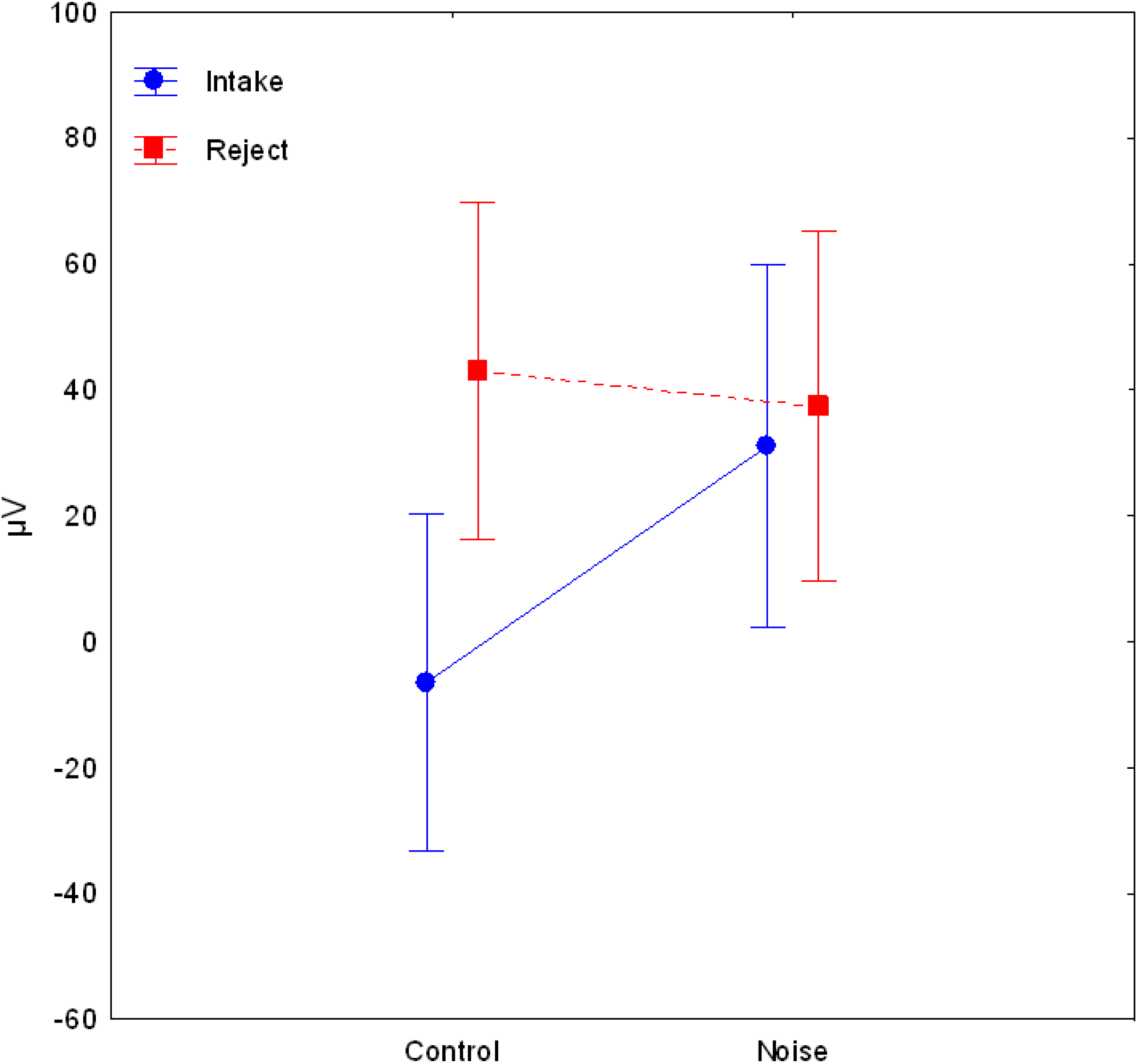
3. Results




4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Basner, M.; Babisch, W.; Davis, A.; Brink, M.; Clark, C.; Janssen, S.; Stansfeld, S. Auditory and non-auditory effects of noise on health. Lancet 2014, 383, 1325–1332. [Google Scholar]
- Babisch, W. The noise/stress concept, risk assessment and research needs. Noise Health 2002, 4, 1–11. [Google Scholar]
- Jahncke, A.; Hygge, S.; Halin, N.; Green, A.M.; Dimberg, K. Open-plan office noise: Cognitive performance and restoration. J. Environ. Psychol. 2011, 31, 373–382. [Google Scholar]
- Maschke, C.; Niemann, H. Health effects of annoyance induced by neighbour noise. Noise Control Eng. J. 2007, 55, 348–356. [Google Scholar]
- Lercher, P.; Botteldooren, D.; Widmann, U.; Uhrner, U.; Kammeringer, E. Cardiovascular effects of environmental noise: Research in Austria. Noise Health 2011, 13, 234–250. [Google Scholar]
- Shepherd, D.; Welch, D.; Dirks, K.N.; McBride, D. Do quiet areas afford greater health-related quality of life than noisy areas? Int. J. Environ. Res. Public Health 2013, 10, 1284–1303. [Google Scholar]
- Stansfeld, S.A.; Matheson, M.P. Noise pollution: Non-auditory effects on health. Br. Med. Bull. 2003, 68, 243–257. [Google Scholar]
- Woolner, P.; Hall, E. Noise in schools: A holistic approach to the issue. Int. J. Environ. Res. Public Health 2010, 7, 3255–3269. [Google Scholar]
- Trimmel, M.; Poelzl, G. Impact of background noise on reaction time and brain DC potential changes of VDT-based spatial attention. Ergonomics 2006, 49, 202–208. [Google Scholar]
- Trimmel, M.; Atzlsdorfer, J.; Tupy, N.; Trimmel, K. Effects of low intensity noise from aircraft or from neighbourhood on cognitive learning and electrophysiological stress responses. Int. J. Hyg. Environ. Health 2012, 215, 547–554. [Google Scholar]
- Hygge, S.; Boman, E.; Enmarker, I. The effects of road traffic noise and meaningful irrelevant speech on different memory systems. Scand. J. Psychol. 2003, 44, 13–21. [Google Scholar]
- James, W. The Principles of Psychology; Holt: New York, NY, USA, 1890. [Google Scholar]
- Milliken, B.; Tipper, S. Attention and inhibition. In Attention; Pashler, H., Ed.; Psychology Press: Hove, UK, 1998. [Google Scholar]
- Birbaumer, N.; Elbert, T.; Canavan, A.G.; Rockstroh, B. Slow potentials of the cerebral cortex and behavior. Physiol. Rev. 1990, 70, 1–41. [Google Scholar]
- Rösler, F.; Heil, M.; Röder, B. Slow negative brain potentials as reflections of specific modular resources of cognition. Biol. Psychol. 1997, 45, 109–141. [Google Scholar]
- Haider, M.; Groll-Knapp, E.; Ganglberger, J.A. Event-related slow (DC) potentials in the human brain. Rev. Physiol. Biochem. Pharmacol. 1981, 88, 125–197. [Google Scholar]
- Lacey, J.I. Psychophysiological approaches to the evaluation of psychotherapeutic process and outcome. In Research in Psychotherapy; Rubinstein, E.A., Parloff, M.B., Eds.; American Psychological Association: Washington, DC, USA, 1959. [Google Scholar]
- Ray, W.; Cole, H. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar]
- Cooper, N.; Croft, R.; Dominey, S.; Burgess, A.; Gruzelier, J. Paradox lost? Exploring the role of alpha oscillations during externally vs. internally directed attention and the implications for idling and inhibition hypotheses. Int. J. Psychophysiol. 2003, 47, 65–74. [Google Scholar]
- Cooper, N.; Burgess, A.; Croft, R.; Gruzelier, J. Investigating evoked and induced electroencephalogram activity in task-related alpha power increases during an internally directed attention task. Neuroreport 2006, 17, 205–208. [Google Scholar]
- Somjen, G.G. Contribution of neuroglia to extracellular sustained potential shifts. In Multidisciplinary Perspectives in Event-Related Brain Potential Research; Otto, D.A., Ed.; US Environmental Protection Agency: Washington, DC, USA, 1978; pp. 19–24. [Google Scholar]
- Caspers, H.; Speckmann, E.J.; Lehmenkueler, A. Electrogenesis of slow potentials. In Self-Regulation of Brain and Behavior; Elbert, T., Rockstroh, B., Lutzenberger, W., Birbaumer, N., Eds.; Springer: Berlin, Germany, 1984; pp. 26–41. [Google Scholar]
- Bauer, H.; Korunka, C.; Leodolter, M. Possible glial contribution in the electrogenesis of SPs. In Slow Potential Changes in the Human Brain; McCallum, W.C., Curry, S.H., Eds.; Plenum: New York, NY, USA, 1993; pp. 23–24. [Google Scholar]
- Caspers, H.; Speckmann, E.J.; Lehmenkuehler, A. Effects of CO2 on cortical field potentials in relation to neuronal activity. In Origin of Cerebral Field Potentials; Speckmann, E.J., Caspers, H., Eds.; Georg Thieme: Stuttgart, Germany, 1979; pp. 151–163. [Google Scholar]
- Vuopio, J.; Tallgren, P.; Heinonen, E.; Vanhatalo, S.; Kaila, K. Millivolt-scale DC shifts in the human scalp EEG: Evidence for a nonneuronal generator. J. Neurophysiol. 2003, 89, 2208–2214. [Google Scholar]
- Nita, D.A.; Vanhatalo, S.; Lafortune, F.D.; Voipio, J.; Kaila, K.; Amzica, F. Nonneuronal origin of CO2-related DC EEG shifts: An in vivo study in the cat. J. Neurophysiol. 2004, 92, 1011–1022. [Google Scholar]
- Hiltunen, T.; Kantola, J.; Abou Elseoud, A.; Lepola, P.; Suominen, K.; Starck, T.; Nikkinen, J.; Remes, J.; Tervonen, O.; Palva, S.; et al. Infra-slow EEG fluctuations are correlated with resting-state network dynamics in fMRI. J. Neurosci. 2014, 34, 356–362. [Google Scholar]
- Monto, S.; Palva, S.; Voipio, J.; Palva, J.M. Very slow EEG fluctuations predict the dynamics of stimulus detection and oscillation amplitudes in humans. J. Neurosci. 2008, 28, 8268–8272. [Google Scholar]
- Trimmel, M.; Mikowitsch, A.; Groll-Knapp, E.; Haider, M. Occurrence of infraslow potential oscillations in relation to task, ability to concentrate and intelligence. Int. J. Psychophysiol. 1990, 9, 167–170. [Google Scholar]
- Stamm, J.; Rosen, S. Cortical steady potential shifts and anodal polarization during delayed response performance. Acta Neurobiol. Exp. 1972, 32, 193–209. [Google Scholar]
- Bauer, H.; Nirnberger, G. Concept identification as a function of preceding negative or positive spontaneous shifts in slow brain potentials. Psychophysiology 1981, 18, 466–469. [Google Scholar]
- Pirch, J.H.; Peterson, S.L. Event-related slow potentials and activity of singly neurons in rat frontal cortex. Int. J. Neurosci. 1981, 15, 141–146. [Google Scholar]
- Trimmel, M.; Meixner-Pendleton, M. Relationship of brain DC potentials to vigilance-based ERPs and operator functional state. In Operator Functional State. The Assessment and Prediction of Human Performance Degradation in Complex Tasks; Hockey, G.R., Gaillard, A.W., Burow, O., Eds.; IOS Press: Amsterdam, The Netherland, 2003; pp. 131–139. [Google Scholar]
- Marshall, L.; Mölle, M.; Fehm, H.; Born, J. Scalp recorded direct current brain potentials during human sleep. Eur. J. Neurosci. 1998, 10, 1167–1178. [Google Scholar]
- Rowland, V. Dynamic patterns of brain cell assemblies. IV. Mixed systems. Steady states and their shifts. Behavioral studies. Neurosci. Res. Prog. Bull. 1974, 12, 109–121. [Google Scholar]
- Marczynski, T.J.; Karmos, G. Reward contingent positive variation and patterns of neuronal activity in the visual cortex of the cat. In Multidisciplinary Perspectives in Event-Related Brain Potential Research; Otto, D.A., Ed.; Environmental Protection Agency: Washington, DC, USA, 1978; pp. 36–38. [Google Scholar]
- Posner, M.I.; Snyder, C.R.; Davidson, B.J. Attention and the detection of signals. J. Exp. Psychol. 1980, 109, 160–174. [Google Scholar]
- Moore, W.H.; Haynes, W.O. A study of alpha hemispheric asymmetries for verbal and nonverbal stimuli in males and females. Brain Lang. 1980, 9, 338–349. [Google Scholar]
- Rugg, M.; Dickens, A. Dissociation of alpha and theta activity as a function of verbal and visuospatial tasks. Electr. Clin. Neuro 1982, 53, 201–207. [Google Scholar]
- Trimmel, M.; Groll-Knapp, E.; Haider, M. Different storage methods for biopotential skin electrodes (sintermetallic Ag/AgCl) and their influence on the bias potential. Eur. J. Appl. Physiol. 1982, 50, 105–116. [Google Scholar]
- Seyfried, H. Lern- und Merkfähigkeitstest (LMT); Ketterl: Wien, Austria, 1990. [Google Scholar]
- Schoppe, K.J. Verbaler Kreativitätstest-Ein Verfahren zur Erfassung verbal-produktiver Kreativitätsmerkmale; Verlag für Psychologie: Göttingen, Germany, 1975. [Google Scholar]
- Torrance, E.P. Torrance Test of Creative Thinking; Scholastic Testing Service, Inc.: Bensenville, IL, USA, 1974. [Google Scholar]
- Hennighausen, E.; Heil, M.; Rösler, F. A correction method for DC drift artifacts. Electr. Clin. Neuro 1993, 86, 199–204. [Google Scholar]
- Haider, M.; Groll-Knapp, E.; Trimmel, M. Cortical DC-shifts related to sustained sensory stimulation and motor activity. In From Neuron to Action; Deecke, L., Eccles, J.C., Mountcastle, V.B., Eds.; Springer-Verlag: Berlin, Germany, 1990; pp. 59–64. [Google Scholar]
- Ghatan, P.H.; Hsieh, J.C.; Petersson, K.M.; Stone-Elander, S.; Ingvar, M. Coexistence of attention-based facilitation and inhibition in the human cortex. Neuroimage 1998, 7, 23–29. [Google Scholar]
- Purser, J.; Radford, A.N. Acoustic noise induces attention shifts and reduces foraging performance in three-spined sticklebacks (Gasterosteus aculeatus). PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Clark, C.; Sörqvist, P. A 3 year update on the influence of noise on performance and behavior. Noise Health 2012, 14, 292–296. [Google Scholar]
- Sörqvist, P.; Nöstl, A.; Halin, N. Disruption of writing processes by the semanticity of background speech. Scand. J. Psychol. 2012, 53, 97–102. [Google Scholar]
- Valente, D.L.; Plevinsky, H.M.; Franco, J.M.; Heinrichs-Graham, E.C.; Lewis, D.E. Experimental investigation of the effects of the acoustical conditions in a simulated classroom on speech recognition and learning in children. J. Acoust. Soc. Am. 2012, 131, 232–246. [Google Scholar]
- Haines, M.M.; Stansfeld, S.A.; Job, R.F.S.; Berglund, B.; Head, J. Chronic aircraft noise exposure, stress responses, mental health and cognitive performance in school children. Psychol. Med. 2001, 31, 265–277. [Google Scholar]
- Lercher, P.; Evans, G.W.; Meis, M. Ambient noise and cognitive processes among primary schoolchildren. Environ. Behav. 2003, 35, 725–735. [Google Scholar]
- Hygge, S. Classroom experiments on the effects of different noise sources and sound levels on long-term recall and recognition in children. Appl. Cogn. Psychol. 2003, 17, 895–914. [Google Scholar]
- Rösler, F.; Heil, M.; Glowalla, U. Monitoring retrieval from long-term memory by slow event-related brain potentials. Psychophysiology 1993, 30, 170–182. [Google Scholar]
- Brunia, C.H.; Damen, E.J. Distribution of slow brain potentials related to motor preparation and stimulus anticipation in a time estimation task. Electr. Clin. Neuro 1988, 69, 234–243. [Google Scholar]
- Trimmel, M.; Kundi, M.; Binder, E.; Groll-Knapp, E.; Haider, M. Combined effects of mental load and background noise on CNS activity indicated by brain DC potentials. Environ. Int. 1996, 22, 83–92. [Google Scholar]
- Trimmel, M.; Kundi, M.; Binder, G. Effects of background noise and mental load on CNS activity indicated by brain DC potentials. Zbl. Hyg. Umweltmed. 1997, 200, 415–416. [Google Scholar]
- Andringa, T.C.; Lanser, J.J. How pleasant sounds promote and annoying sounds impede health: A cognitive approach. Int. J. Environ. Res. Public Health 2013, 10, 1439–1461. [Google Scholar]
- Wickens, C.D. Multiple resources and mental workload. Hum. Fact. 2008, 50, 449–455. [Google Scholar]
- Hommel, B.; Fischer, R.; Lorenza, S.C.; van den Wildenberg, W.P.; Cellini, C. The effect of fMRI (noise) on cognitive control. J. Exp. Psychol. 2012, 38, 290–301. [Google Scholar]
- Addison, J.; Dancer, J.; Montague, J.; Davis, P. Ambient noise levels in university classrooms: detrimental to teaching and learning. Percept. Mot. Skills 1999, 89, 649–650. [Google Scholar]
- Michalek, A.M.P.; Watson, S.M.; Ash, I.; Ringleb, S.; Raymer, A. Effects of noise and audiovisual cues on speech processing in adults with and without ADHD. Int. J. Audiol. 2014, 53, 145–152. [Google Scholar]
- Beattie, R.L.; Lu, Z.-L.; Manis, F.R. Dyslexic adults can learn from repeated stimulus presentation but have difficulties in excluding external noise. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Helfer, K.S.; Freyman, R.L. Stimulus and listening factors affecting age-related changes in competing speech perception. J. Acoust. Soc. Am. 2014, 136, 748–759. [Google Scholar]
- Wong, P.C.M.; Jin, J.X.; Gunasekera, G.M.; Abel, R.; Lee, E.R.; Dhar, S. Aging and cortical mechanisms of speech perception in noise. Neuropsychologia 2009, 47, 693–703. [Google Scholar]
- Mattys, S.L.; Scharenborg, O. Phoneme categorization and discrimination in younger and older adults: A comparative analysis of perceptual, lexical, and attentional factors. Psychol. Aging 2014, 29, 150–162. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trimmel, K.; Schätzer, J.; Trimmel, M. Acoustic Noise Alters Selective Attention Processes as Indicated by Direct Current (DC) Brain Potential Changes. Int. J. Environ. Res. Public Health 2014, 11, 9938-9953. https://doi.org/10.3390/ijerph111009938
Trimmel K, Schätzer J, Trimmel M. Acoustic Noise Alters Selective Attention Processes as Indicated by Direct Current (DC) Brain Potential Changes. International Journal of Environmental Research and Public Health. 2014; 11(10):9938-9953. https://doi.org/10.3390/ijerph111009938
Chicago/Turabian StyleTrimmel, Karin, Julia Schätzer, and Michael Trimmel. 2014. "Acoustic Noise Alters Selective Attention Processes as Indicated by Direct Current (DC) Brain Potential Changes" International Journal of Environmental Research and Public Health 11, no. 10: 9938-9953. https://doi.org/10.3390/ijerph111009938
APA StyleTrimmel, K., Schätzer, J., & Trimmel, M. (2014). Acoustic Noise Alters Selective Attention Processes as Indicated by Direct Current (DC) Brain Potential Changes. International Journal of Environmental Research and Public Health, 11(10), 9938-9953. https://doi.org/10.3390/ijerph111009938