Effects of Nano-MnO2 on Dopaminergic Neurons and the Spatial Learning Capability of Rats

Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents and Instruments
2.2. Experimental Designs
2.3. Animal Experiments
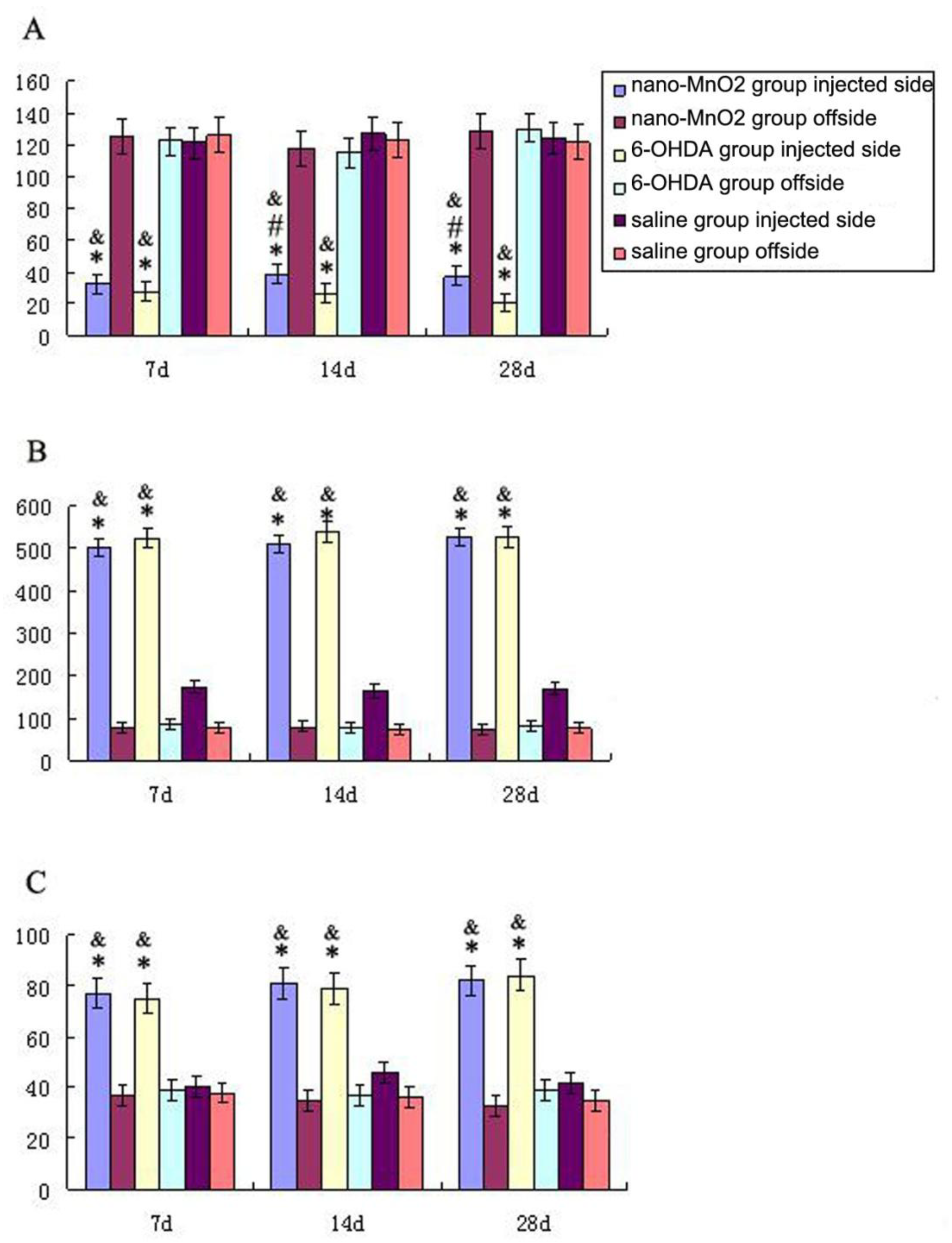
2.4. Rotation Behavior
2.5. Morris Water Maze Test
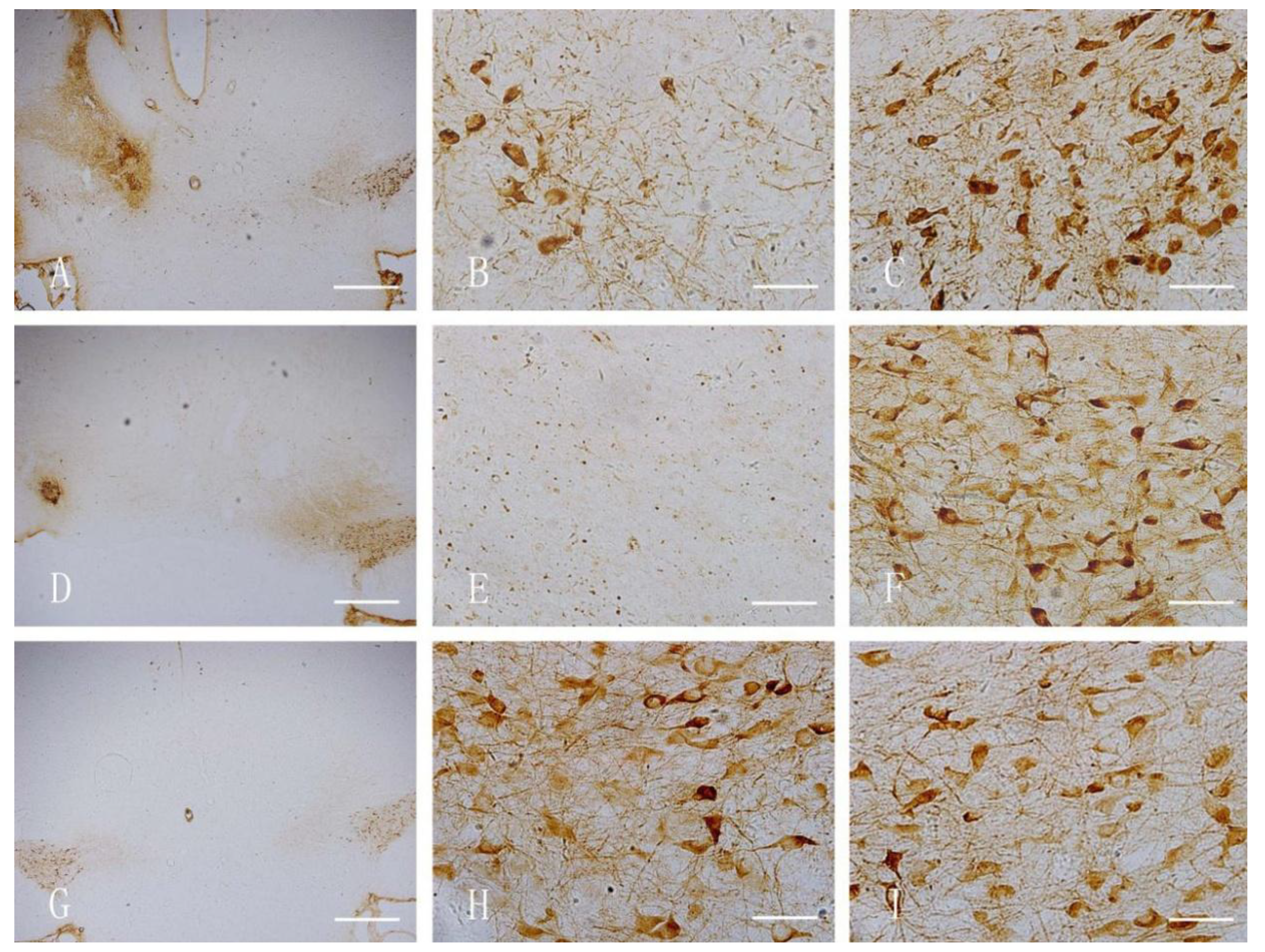
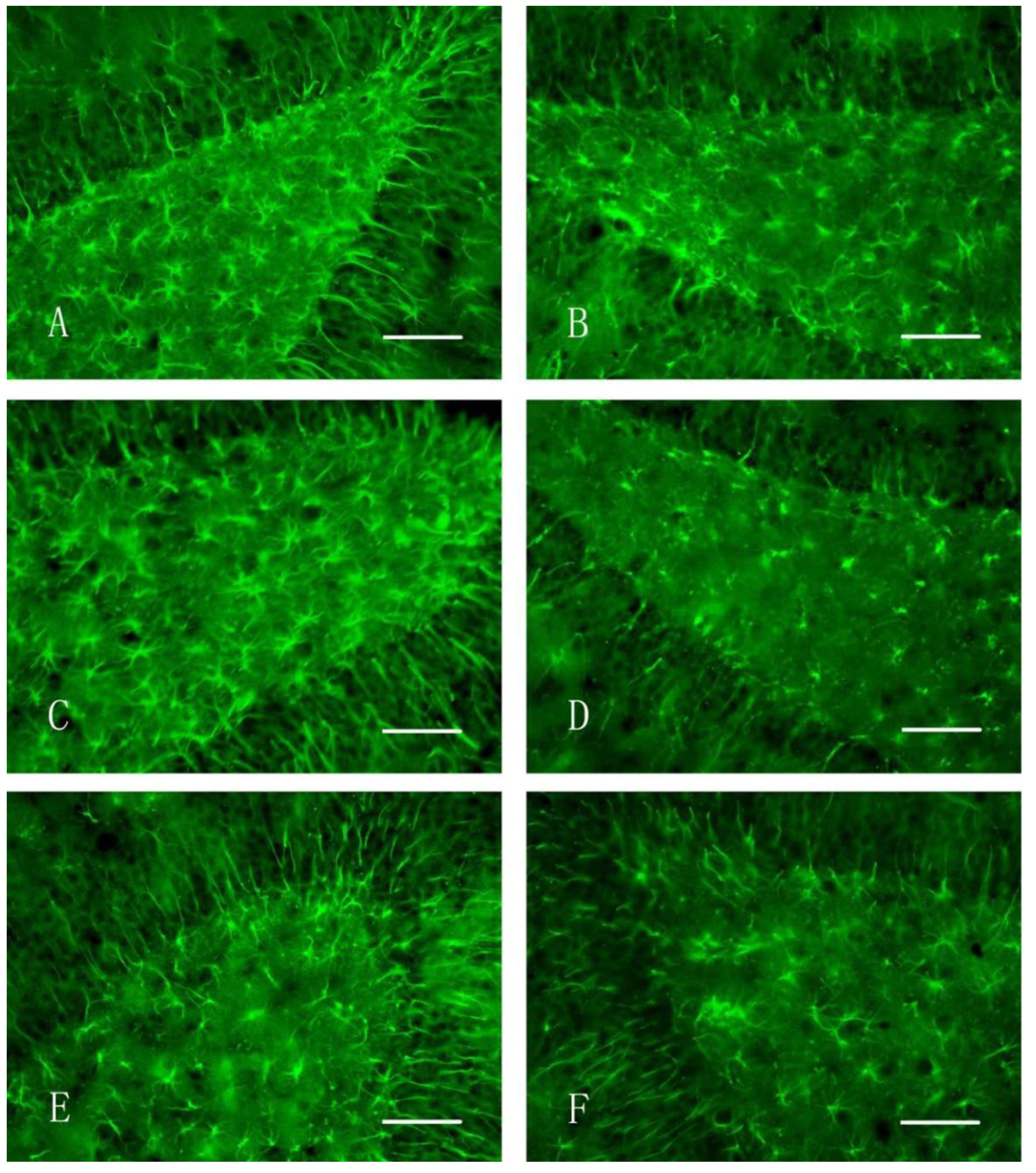
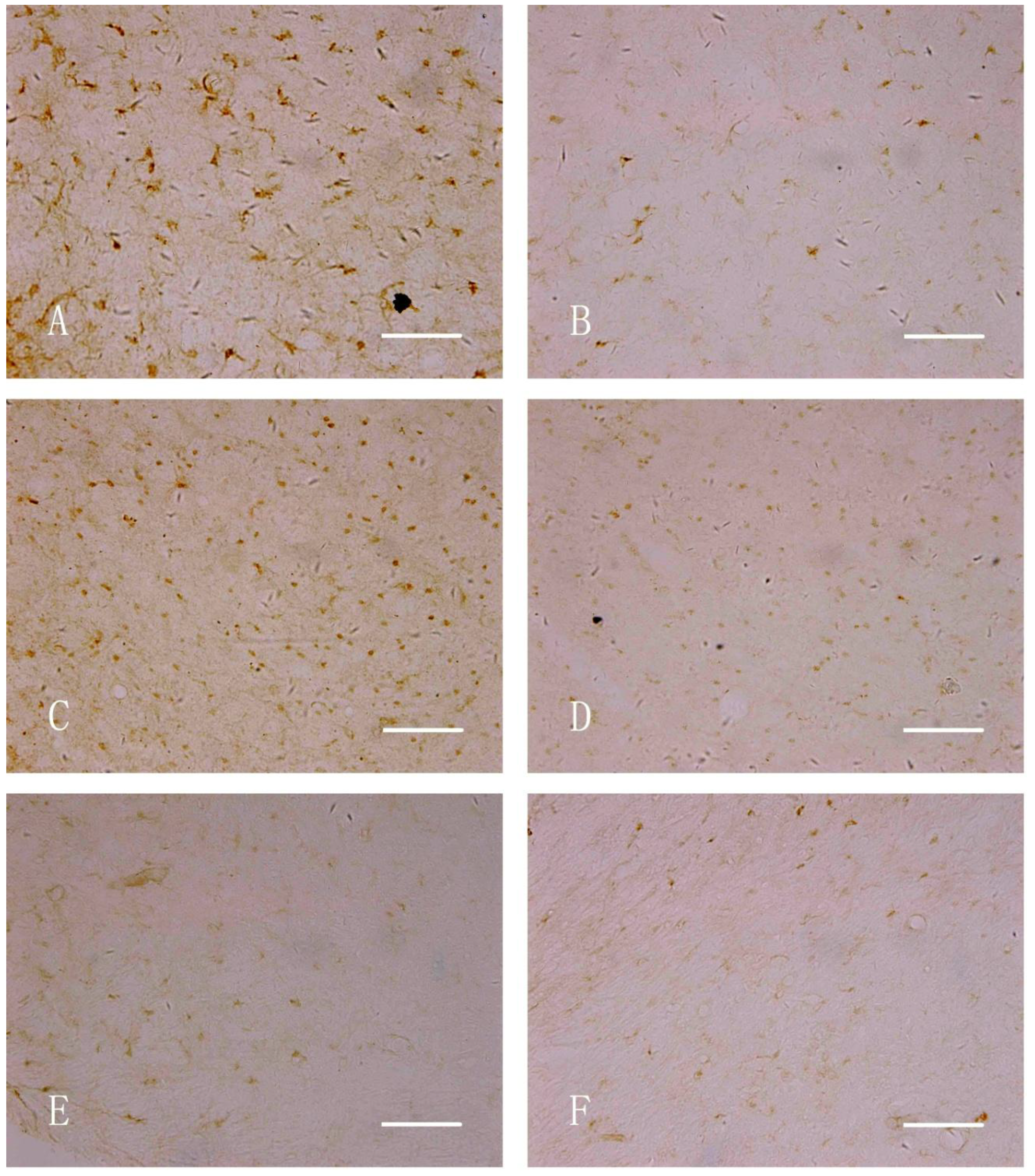
2.6. Immunohistochemical Staining
2.7. Statistical Analysis
3. Results and Discussion
3.1. Motor Behavioral Study
3.2. Morris Water Maze Test

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 7 Days after Injection | 14 Days after Injection | |||||
|---|---|---|---|---|---|---|
| Saline | 6-OHDA | Nano-MnO2 | Saline | 6-OHDA | Nano-MnO2 | |
| Escape latencies (s) | 27.62 ± 28.73 | 64.68 ± 43.62 ** | 63.89 ± 42.73 ** | 25.12 ± 29.03 | 67.69 ± 51.61 ** | 60.52 ± 41.24 ##,** |
| Velocity (cm/s) | 24.81 ± 4.35 | 19.28 ± 3.77 * | 21.59 ± 3.61 | 23.75±3.69 | 18.72 ± 4.65 * | 21.64 ± 3.32 |
3.3. Immunohistochemical Staining




4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Post, J.E. Manganese oxide minerals: Crystal structures and economic and environmental significance. Proc. Natl. Acad. Sci. 1999, 96, 3447–3454. [Google Scholar] [CrossRef]
- Addess, K.J.; Basilion, J.P. Structure and dynamics of the iron responsive element RNA: Implications for binding of the RNA by iron regulatory binding proteins. J. Mol. Biol. 1997, 274, 72–83. [Google Scholar] [CrossRef]
- Aschner, M.; Gannon, M. Manganese uptake and efflux in cultured rat astrocytes. J. Neurochem. 1992, 58, 730–735. [Google Scholar] [CrossRef]
- Fitsanakis, V.A.; Aschner, M. The importance of glutamate, glycine, and gamma-aminobutyric acid transport and regulation in manganese, mercury and lead neurotoxicity. Toxicol. Appl. Pharmacol. 2005, 204, 343–354. [Google Scholar] [CrossRef]
- Liao, S.L.; Chen, C.J. Manganese stimulates stellation of cultured rat cortical astrocytes. Neuroreport 2001, 12, 3877–3881. [Google Scholar] [CrossRef]
- Calne, D.B.; Chu, N.S. Manganism and idiopathic parkinsonism: Similarities and differences. Neurology 1994, 44, 1583–1586. [Google Scholar] [CrossRef]
- Cersosimo, M.G.; Koller, W.C. The diagnosis of manganese-induced parkinsonism. Neurotoxicology 2006, 27, 340–346. [Google Scholar] [CrossRef]
- Olanow, C.W. Manganese-induced parkinsonism and Parkinson’s disease. Ann. N. Y. Acad. Sci. 2004, 1012, 209–223. [Google Scholar] [CrossRef]
- Sousa, F.; Mandal, S. Functionalized gold nanoparticles: A detailed in vivo multimodal microscopic brain distribution study. Nanoscale 2010, 2, 2826–2834. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Li, M.Q. In vivo toxicity of nano-alumina on mice neurobehavioral profiles and the potential mechanisms. Int. J. Immunopathol. Pharmacol. 2011, 24, S23–S29. [Google Scholar]
- Ragupathy, P.; Vasan, H.N. Synthesis and characterization of Nano-MnO2 for electrochemical supercapacitor studies. J. Electrochem. Soc. 2008, 155, A34–A40. [Google Scholar] [CrossRef]
- Jothiramalingam, R.; Wang, M.K. Synthesis, characterization and photocatalytic activity of porous manganese oxide doped titania for toluene decomposition. J. Hazard. Mater. 2007, 147, 562–569. [Google Scholar] [CrossRef]
- Felton, C.; Karmakar, A. Magnetic nanoparticles as contrast agents in biomedical imaging: Recent advances in iron- and manganese-based magnetic nanoparticles. Drug Metab. Rev. 2014, 46, 142–154. [Google Scholar] [CrossRef]
- Kurokawa, A.; Yanoh, T. Preparation and magnetic properties of multiferroic CuMnO2 nanoparticles. J. Nanosci. Nanotechnol. 2014, 14, 2553–2556. [Google Scholar] [CrossRef]
- Antonini, J.M.; Afshari, A.A. Design, construction, and characterization of a novel robotic welding fume generator and inhalation exposure system for laboratory animals. J. Occup. Environ. Hyg. 2006, 3, 194–203. [Google Scholar] [CrossRef]
- Long, Z.; Jiang, Y.M. Vulnerability of welders to manganese exposur—A neuroimaging study. Neurotoxicology 2014, 3. [Google Scholar] [CrossRef]
- Hu, Y.L.; Gao, J.Q. Potential neurotoxicity of nanoparticles. Int. J. Pharm. 2010, 394, 115–121. [Google Scholar] [CrossRef]
- Lucchini, R.G.; Guazzetti, S. Neurofunctional dopaminergic impairment in elderly after lifetime exposure to manganese. Neurotoxicology 2014, 5. [Google Scholar] [CrossRef]
- Gerber, G.B.; Leonard, A. Carcinogenicity, mutagenicity and teratogenicity of manganese compounds. Crit. Rev. Oncol. Hematol. 2002, 42, 25–34. [Google Scholar] [CrossRef]
- Gonzalez-Cuyar, L.F.; Nelson, G. Quantitative neuropathology associated with chronic manganese exposure in South African mine workers. Neurotoxicology 2013, 12. [Google Scholar] [CrossRef]
- Benedetto, A.; Au, C. Manganese-induced dopaminergic neurodegeneration: Insights into mechanisms and genetics shared with Parkinson’s disease. Chem. Rev. 2009, 109, 4862–4884. [Google Scholar] [CrossRef]
- Riquelme, E.; Abarca, J. An NR2B-dependent decrease in the expression of trkB receptors precedes the disappearance of dopaminergic cells in Substantia Nigra in a rat model of Presymptomatic Parkinson’s Disease. Parkinson’s Dis. 2012, 2012. [Google Scholar] [CrossRef]
- Albani, S.H.; McHail, D.G. Developmental studies of the hippocampus and hippocampal-dependent behaviors: Insights from interdisciplinary studies and tips for new investigators. Neurosci. Biobehav. Rev. 2014, 43C, 183–190. [Google Scholar] [CrossRef]
- Frank, L.M.; Brown, E.N. Hippocampal and cortical place cell plasticity: Implications for episodic memory. Hippocampus 2006, 16, 775–784. [Google Scholar] [CrossRef]
- Holahan, M.R.; Routtenberg, A. Lidocaine injections targeting CA3 hippocampus impair long-term spatial memory and prevent learning-induced mossy fiber remodeling. Hippocampus 2011, 21, 532–540. [Google Scholar] [CrossRef]
- Blecharz-Klin, K.; Piechal, A. Effect of intranasal manganese administration on neurotransmission and spatial learning in rats. Toxicol. Appl. Pharmacol. 2012, 265, 1–9. [Google Scholar] [CrossRef]
- Fitsanakis, V.A.; Thompson, K.N. A chronic iron-deficient/high-manganese diet in rodents results in increased brain oxidative stress and behavioral deficits in the morris water maze. Neurotox. Res. 2009, 15, 167–178. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Y. Effects of nanoparticle zinc oxide on spatial cognition and synaptic plasticity in mice with depressive-like behaviors. J. Biomed. Sci. 2012, 19. [Google Scholar] [CrossRef] [Green Version]
- Mohammadipour, A.; Hosseini, M. The effects of exposure to titanium dioxide nanoparticles during lactation period on learning and memory of rat offspring. Toxicol. Ind. Health 2013, 9. [Google Scholar] [CrossRef]
- Afeseh Ngwa, H.; Kanthasamy, A. Manganese nanoparticle activates mitochondrial dependent apoptotic signaling and autophagy in dopaminergic neuronal cells. Toxicol. Appl. Pharmacol. 2011, 256, 227–240. [Google Scholar] [CrossRef]
- Wang, J.; Rahman, M.F. Expression changes of dopaminergic system-related genes in PC12 cells induced by manganese, silver, or copper nanoparticles. Neurotoxicology 2009, 30, 926–933. [Google Scholar] [CrossRef]
- Wu, J.; Wang, C. Neurotoxicity of silica nanoparticles: Brain localization and dopaminergic neurons damage pathways. ACS Nano 2011, 5, 4476–4489. [Google Scholar] [CrossRef]
- Lee, J.K.; Sayers, B.C. Multi-walled carbon nanotubes induce COX-2 and iNOS expression via MAP kinase-dependent and -independent mechanisms in mouse RAW264.7 macrophages. Part. Fibre Toxicol. 2012, 9. [Google Scholar] [CrossRef]
- Gojova, A.; Guo, B. Induction of inflammation in vascular endothelial cells by metal oxide nanoparticles: Effect of particle composition. Environ. Health Perspect. 2007, 115, 403–409. [Google Scholar] [CrossRef]
- Muller, L.; Riediker, M. Oxidative stress and inflammation response after nanoparticle exposure: Differences between human lung cell monocultures and an advanced three-dimensional model of the human epithelial airways. J. R. Soc. Interface 2009, 7, S27–S40. [Google Scholar] [CrossRef]
- Buskila, Y.; Abu-Ghanem, Y. Enhanced astrocytic nitric oxide production and neuronal modifications in the neocortex of a NOS2 mutant mouse. PLoS One 2007, 2. [Google Scholar] [CrossRef]
- Tieu, K.; Ischiropoulos, H. Nitric oxide and reactive oxygen species in Parkinson’s disease. IUBMB Life 2003, 55, 329–335. [Google Scholar] [CrossRef]
- Stevenson, L.; Matesanz, N. Reduced nitro-oxidative stress and neural cell death suggests a protective role for microglial cells in TNFalpha-/- mice in ischemic retinopathy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3291–3299. [Google Scholar] [CrossRef]
- Shiratsuch, H.; Basson, M.D. Differential regulation of monocyte/macrophage cytokine production by pressure. Am. J. Surg. 2005, 190, 757–762. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, T.; Shi, T.; Li, X.; Zeng, S.; Yin, L.; Pu, Y. Effects of Nano-MnO2 on Dopaminergic Neurons and the Spatial Learning Capability of Rats. Int. J. Environ. Res. Public Health 2014, 11, 7918-7930. https://doi.org/10.3390/ijerph110807918
Li T, Shi T, Li X, Zeng S, Yin L, Pu Y. Effects of Nano-MnO2 on Dopaminergic Neurons and the Spatial Learning Capability of Rats. International Journal of Environmental Research and Public Health. 2014; 11(8):7918-7930. https://doi.org/10.3390/ijerph110807918
Chicago/Turabian StyleLi, Tao, Tingting Shi, Xiaobo Li, Shuilin Zeng, Lihong Yin, and Yuepu Pu. 2014. "Effects of Nano-MnO2 on Dopaminergic Neurons and the Spatial Learning Capability of Rats" International Journal of Environmental Research and Public Health 11, no. 8: 7918-7930. https://doi.org/10.3390/ijerph110807918