Identification of Diarrheagenic Escherichia coli Strains from Avian Organic Fertilizers

 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples and Bacterial Strains
2.2. DNA Extraction
2.3. Detecting virulence genes using polymerase chain reaction (PCR)

{kind=link}
| Gene | PCR primer | Primer sequence (5’–3’) | Fragment (bp) | Concentration (pmol/µL) | ControlStrains | Reference |
|---|---|---|---|---|---|---|
| Primers used for multiplex PCR described by Aranda and collaborators [22] | ||||||
| eaeA | F | CTGAACGGCGATTACGCGAA | 917 | 10 | E2348/69 | [24] |
| R | CCAGACGATACGATCCAG | (EPEC) | ||||
| bfpA | F | AATGGTGCTTGCGCTTGCTGC | 326 | 1.25 | E2348/69 | [24] |
| R | GCCGCTTTATCCAACCTGGTA | (EPEC) | ||||
| aggR | F | GTATACACAAAAGAAGGAAGC | 254 | 2.5 | O42 | [25] |
| R | ACAGAATCGTCAGCATCAGC | (EAEC) | ||||
| elt | F | GGCGACAGATTATACCGTGC | 450 | 0.25 | H10407 | [24] |
| R | CGGTCTCTATATTCCCTGTT | (ETEC) | ||||
| est | F | ATTTTTMTTTCTGTATTRTCTT | 190 | 6.47 | B41 | [24] |
| R | CACCCGGTACARGCAGGATT | (ETEC) | ||||
| ipaH | F | GTTCCTTGACCGCCTTTCCGATACCGTC | 600 | 1 | EDL 1284 | [24] |
| R | GCCGGTCAGCCACCCTCTGAGAGTAC | (EIEC) | ||||
| stx | F | GAGCGAAATAATTTATATGTG | 518 | 6 | EDL 933 | [26] |
| R | TGATGATGGCAATTCAGTAT | (EHEC) | ||||
| Primers used for multiplex PCR described by Paton and Paton [23] | ||||||
| eaeA | F | GACCCGGCACAAGCATAAGC | 384 | 10 | E2348 | [23] |
| R | CCACCTGCAGCAACAAGAGG | (EPEC) | ||||
| stx1 | F | ATAAATCGCCATTCGTTGACTAC | 180 | 10 | EDL 933 | [23] |
| R | AGAACGCCCACTGAGATCATC | (EHEC) | ||||
| stx2 | F | GGCACTGTCTGAAACTGCTCC | 255 | 10 | EDL 933 | [23] |
| R | TCGCCAGTTATCTGACATTCTG | (EHEC) | ||||
| ehxA | F | GCATCATCAAGCGTAGCTTCC | 534 | 10 | EDL 933 | [23] |
| R | AATGAGCCAAGCTGGTTAAGCT | (EHEC) | ||||
| Primers used for multiplex PCR described by Clermont and collaborators [27] | ||||||
| chuA | F | GACGAACCAACGGTCAGGAT | 279 | 20 | E. coli | [27] |
| R | TGCCGCCAGTACAAAGACA | J96 | ||||
| yjaA | F | TGAAGTGTCAGAGACGCTG | 211 | 20 | E. coli | [27] |
| R | ATGGAGAATGCGTTCCTCAAC | J96 | ||||
| TspE4C2 | F | GAGTAATGTCGGGGCATTCA | 152 | 20 | E. coli | [27] |
| R | CGCGCCAACAAAGTATTACG | Strain 20 | ||||
2.4. Determination of Phylogenetic Group (PCR)
2.5. Adherence Assays
2.6. Antimicrobial Susceptibility Testing
2.7. Biofilm
2.8. Assay for Cytotoxicity Activity
2.9. Statistical Analysis
3. Results
3.1. E. coli Isolates
3.2. Virulence Factors of Diarrheagenic E. coli (DEC)
3.3. Phylogenetic Group
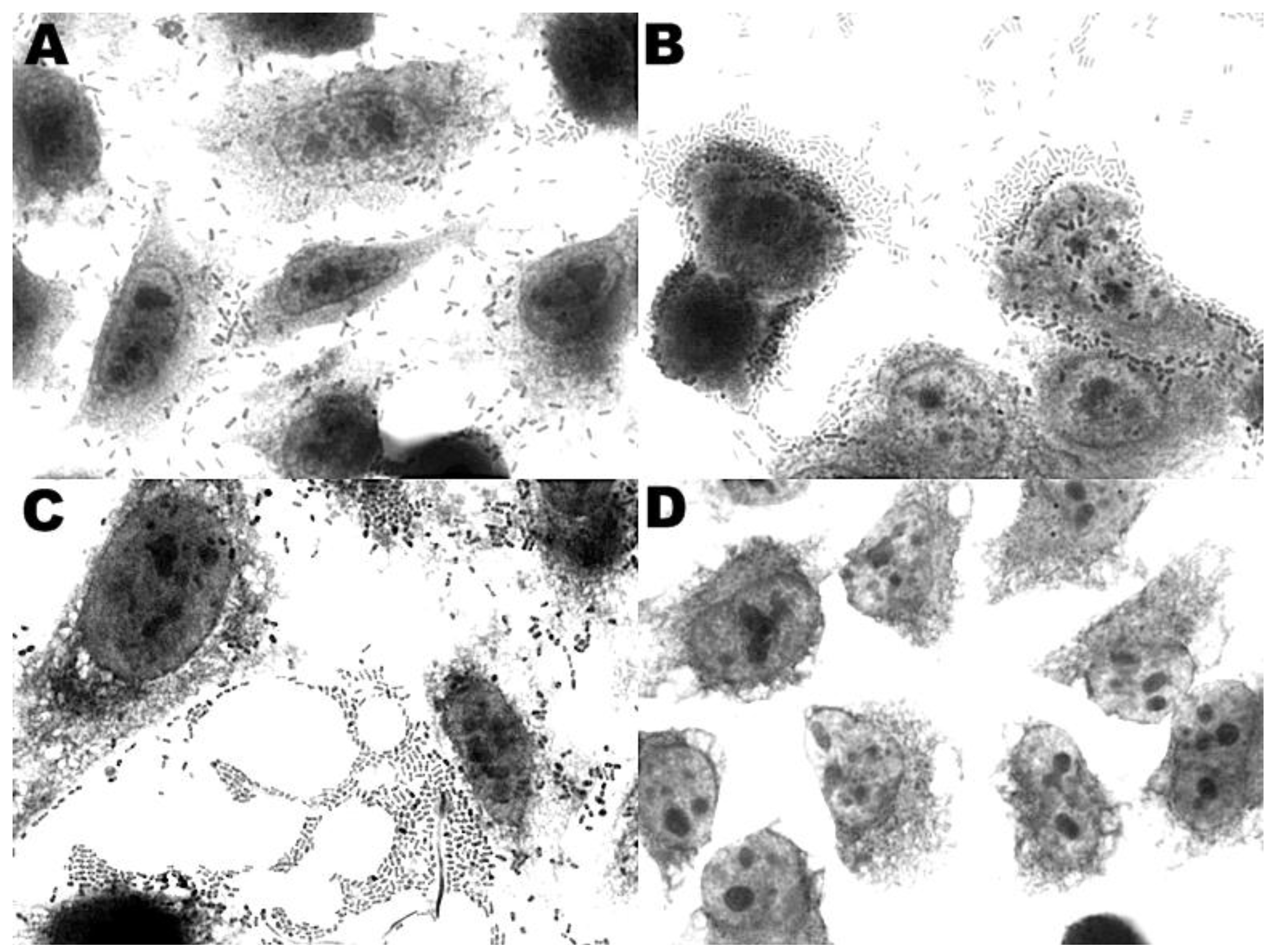
3.4. Adherence to HEp-2 and HeLa Cells
3.5. Antimicrobial Susceptibility

3.6. Biofilm Test
3.7. Evaluation of Cytotoxic Activity
| Isolate of E. coli | Genetic profile | Pattern of adherence in HEp-2 cell | Resistance profile | Biofilm formation (BF) | Phylogenetic group |
|---|---|---|---|---|---|
| OF-4 | eae+ | AA | NAL/AMO/SRT | Strong | A |
| OF-36 | eae+ | AA | Susceptible | Non-BF | B2 |
| OF-42 | eae+/aggR+ | AA | Susceptible | Non-BF | A |
| OF-51 | eae+/aggR+ | AA | AMO a/TET | Non-BF | B1 |
| OF-6 | aggR+ | AA | AMO a | Strong | A |
| OF-8 | aggR+ | AA | AMO a/ACL | Strong | B1 |
| OF-9 | aggR+ | AA | NAL a/AMO a/AMCa/IMPa/AMP | Non-BF | B1 |
| OF-22 | stx1+ | AA | AMO a/AMP a | Strong | A |
| OF-23 | stx1+ | AA | NAL a/AMO a/AMP a | Strong | B1 |
| OF-24 | stx1+ | AA | AMO a | Non-BF | A |
| OF-35 | aggR+ | AA | TET | Moderate | A |
| OF-39 | aggR+ | AA/DA | AMO a/AMP a | Non-BF | B1 |
| OF-44 | aggR+ | AA | STR/SXT/TET | Non-BF | A |
| OF-57 | aggR | AA | STR a | Non-BF | A |
| OF-61 | aggR | AA | NAL/AMO/AMC/AMP/STR/IMP/ TET | Non-BF | A |
4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Doyle, M.P.; Erickson, M.C. Summer meeting 2007—The problems with fresh produce: An overview. J. Appl. Microbiol. 2008, 105, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sivapalasingam, S.; Friedman, C.R.; Cohen, L.; Tauxe, R.V. Fresh produce: A growing cause of outbreaks of foodborne illness in the United States, 1973 through 1997. J. Food Prot. 2004, 67, 2342–2353. [Google Scholar] [PubMed]
- Composting Manure and Sludge. Available online: http://hdl.handle.net/1969.1/87650 (accessed on 29 July 2014).
- Hahn, L. Processamento da Cama de Aviário e Suas Implicações nos Agroecosistemas; Universidade Federal de Santa Catarina: Florianópolis, Brazil, 2004. [Google Scholar]
- Edwards, D.R.; Daniel, T.C. Environmental impacts of on-farm poultry waste disposal—A review. Bioresour. Technol. 1992, 41, 9–33. [Google Scholar] [CrossRef]
- Miller, C.; Heringa, S.; Kim, J.; Jiang, X. Analyzing indicator microorganisms, antibiotic resistant Escherichia coli, and regrowth potential of foodborne pathogens in various organic fertilizers. Foodborne Pathog. Dis. 2013, 10, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Pietronave, S.; Fracchia, L.; Rinaldi, M.; Martinotti, M.G. Influence of biotic and abiotic factors on human pathogens in a finished compost. Water Res. 2004, 38, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jiang, X. The growth potential of Escherichia coli O157:H7, Salmonella spp. and Listeria monocytogenes in dairy manure-based compost in a greenhouse setting under different seasons. J. Appl. Microbiol. 2010, 109, 2095–2104. [Google Scholar]
- Sidhu, J.; Gibbs, R.A.; Ho, G.E.; Unkovich, I. The role of indigenous microorganisms in suppression of Salmonella regrowth in composted biosolids. Water Res. 2001, 35, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Conway, P.L. Microbial Ecology of the Human Large Intestine. In Human colonic bacteria: Role in nutrition, physiology, and pathology; Gibson, G.R., Macfarlane, G.T., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 1–24. [Google Scholar]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [PubMed]
- McEwen, S.A.; Fedorka-Cray, P.J. Antimicrobial use and resistance in animals. Clin. Infect. Dis. 2002, 34, S93–S106. [Google Scholar] [CrossRef] [PubMed]
- van den Bogaard, A.E.; Stobberingh, E.E. Epidemiology of resistance to antibiotics. Links between animals and humans. Int. J. Antimicrob. Agents 2000, 14, 327–335. [Google Scholar]
- Guan, J.; Wasty, A.; Grenier, C.; Chan, M. Influence of temperature on survival and conjugative transfer of multiple antibiotic-resistant plasmids in chicken manure and compost microcosms. Poult. Sci. 2007, 86, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.P.; Hillman, K.; Fenlon, D.R.; Low, J.C. Transfer of antibiotic resistance between commensal and pathogenic members of the enterobacteriaceae under ileal conditions. J. Appl. Microbiol. 2003, 95, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Ewing, W.H. Edwards and Ewing’s Identification of Enterobacteriaceae; Elsevier Science Publishing Company: New York, NY, USA, 1986. [Google Scholar]
- Toledo, M.R.F.; Fontes, C.F.; Trabulsi, L.R. EPM—A modification of Rugai and Araujo medium for simultaneous test of gas production from glucose, H2S, urease and tryptophan deaminase. Rev. Microbiol. 1982, 13, 309–315. [Google Scholar]
- Toledo, M.R.F.; Fontes, C.F.; Trabulsi, L.R. Mili—A medium for detection of motilily, indole, and lysine decarboxylase. Rev. Microbiol. 1982, 13, 230–235. [Google Scholar]
- Koga, V.L.; Tomazetto, G.; Cyoia, P.S.; Neves, M.S.; Vidotto, M.C.; Nakazato, G.; Kobayashi, R.K.T. Molecular screening of virulence genes in extraintestinal pathogenic Escherichia coli isolated from human blood culture in Brazil. Biomed Res Int. 2014, 2014. [Google Scholar] [CrossRef]
- Aranda, K.R.; Fabbricotti, S.H.; Fagundes-Neto, U.; Scaletsky, I.C. Single multiplex assay to identify simultaneously enteropathogenic, enteroaggregative, enterotoxigenic, enteroinvasive and Shiga toxin-producing Escherichia coli strains in brazilian children. FEMS Microbiol. Lett. 2007, 267, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Paton, J.C. Detection and characterization of Shiga toxigenic Escherichia coli by using multiplex pcr assays for stx1, stx2, eaea, enterohemorrhagic E. coli hlya, rfbo111, and rfbo157. J. Clin. Microbiol. 1998, 36, 598–602. [Google Scholar]
- Aranda, K.R.; Fagundes-Neto, U.; Scaletsky, I.C. Evaluation of multiplex pcrs for diagnosis of infection with diarrheagenic Escherichia coli and Shigella spp. J. Clin. Microbiol. 2004, 42, 5849–5853. [Google Scholar] [CrossRef] [PubMed]
- Ratchtrachenchai, O.A.; Subpasu, S.; Ito, K. Investigation on enteroaggregative Escherichia coli infection by multiplex PCR. Bull. Dep. Med. Sci. 1997, 39, 211–220. [Google Scholar]
- Yamasaki, S.; Lin, Z.; Shirai, H.; Terai, A.; Oku, Y.; Ito, H.; Ohmura, M.; Karasawa, T.; Tsukamoto, T.; Kurazono, H.; Takeda, Y. Typing of verotoxins by DNA colony hybridization with poly- and oligonucleotide probes, a bead-enzyme-linked immunosorbent assay, and polymerase chain reaction. Microbiol. Immunol. 1996, 40, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Cravioto, A.; Gross, R.J.; Scotland, S.M.; Rowe, B. An adhesive factor found in strains of Escherichia coli belonging to the traditional infantile enteropathogenic serotypes. Curr. Microbiol. 1979, 3, 95–99. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B.; Robins-Browne, R.; Prado, V.; Vial, P.; Levine, M.M. Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr. Infect. Dis. J. 1987, 6, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Scaletsky, I.C.; Campos, L.C.; Gomes, T.A.; Whittam, T.S.; Trabulsi, L.R. Clonal structure and virulence factors in strains of Escherichia coli of the classic serogroup O55. Infect. Immun. 1996, 64, 2680–2686. [Google Scholar] [PubMed]
- Scaletsky, I.C.; Pedroso, M.Z.; Oliva, C.A.; Carvalho, R.L.; Morais, M.B.; Fagundes-Neto, U. A localized adherence-like pattern as a second pattern of adherence of classic enteropathogenic Escherichia coli to HEp-2 cells that is associated with infantile diarrhea. Infect. Immun. 1999, 67, 3410–3415. [Google Scholar] [PubMed]
- CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. Approved Standards 2012, Vol. M07-A9. [Google Scholar]
- Wakimoto, N.; Nishi, J.; Sheikh, J.; Nataro, J.P.; Sarantuya, J.; Iwashita, M.; Manago, K.; Tokuda, K.; Yoshinaga, M.; Kawano, Y. Quantitative biofilm assay using a microtiter plate to screen for enteroaggregative Escherichia coli. Am. J. Trop. Med. Hyg. 2004, 71, 687–690. [Google Scholar] [PubMed]
- Murakami, J.; Kishi, K.; Hirai, K.; Hiramatsu, K.; Yamasaki, T.; Nasu, M. Macrolides and clindamycin suppress the release of shiga-like toxins from Escherichia coli O157:H7 in vitro. Int. J. Antimicrob. Agents 2000, 15, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Piazza, R.M. Production of Shiga toxin by Shiga toxin-expressing Escherichia coli (STEC) in broth media: From divergence to definition. Lett. Appl. Microbiol. 2007, 45, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Bugarel, M.; Beutin, L.; Martin, A.; Gill, A.; Fach, P. Micro-array for the identification of Shiga toxin-producing Escherichia coli (STEC) seropathotypes associated with hemorrhagic colitis and Hemolytic Uremic Syndrome in humans. Int. J. Food Microbiol. 2010, 142, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic o island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar] [CrossRef] [PubMed]
- Besser, R.E.; Lett, S.M.; Weber, J.T.; Doyle, M.P.; Barrett, T.J.; Wells, J.G.; Griffin, P.M. An outbreak of diarrhea and Hemolytic Uremic Syndrome from Escherichia coli O157:H7 in fresh-pressed apple cider. JAMA 1993, 269, 2217–2220. [Google Scholar] [CrossRef] [PubMed]
- Nataro, J.P. Enteroaggregative Escherichia coli pathogenesis. Curr. Opin. Gastroenterol. 2005, 21, 4–8. [Google Scholar] [PubMed]
- Brzuszkiewicz, E.; Thurmer, A.; Schuldes, J.; Leimbach, A.; Liesegang, H.; Meyer, F.D.; Boelter, J.; Petersen, H.; Gottschalk, G.; Daniel, R. Genome sequence analyses of two isolates from the recent Escherichia coli outbreak in Germany reveal the emergence of a new pathotype: Entero-aggregative-haemorrhagic Escherichia coli (EAHEC). Arch. Microbiol. 2011, 193, 883–891. [Google Scholar] [CrossRef]
- CDC. Outbreak of Escherichia coli O157:H7 infections among children associated with farm visits-Pennsylvania and Washington, 2000. MMWR. 2001, 50, 293–297. [Google Scholar]
- Trabulsi, L.R.; Keller, R.; Tardelli Gomes, T.A. Typical and atypical enteropathogenic Escherichia coli. Emerg. Infect. Dis. 2002, 8, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Elliot, S.; Sperandio, V.; Perna, N.T.; Mayhew, F.; Blattner, F.R. Attaching-and-effacing intestinal histopathology and the locus of enterocyte effacement. In Escherichia coli O157:H7 and other Shiga toxin-producing E. coli strains; Kaper, J.B., O'Brien, A.D., Eds.; American Society for Microbiology: Washington, DC, USA, 1998; pp. 1–182. [Google Scholar]
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef]
- Castro, A.F.P. Virulence factors present in cultures of Escherichia coli isolated from pigs in the region of Concordia; Santa Catarina, Brazil. Pesq. Vet. Bras. 1984, 4, 109–114. [Google Scholar]
- Nakazato, G.; Gyles, C.; Ziebell, K.; Keller, R.; Trabulsi, L.R.; Gomes, T.A.; Irino, K.; Da Silveira, W.D.; Pestana De Castro, A.F. Attaching and effacing Escherichia coli isolated from dogs in Brazil: Characteristics and serotypic relationship to human enteropathogenic E. coli (EPEC). Vet. Microbiol. 2004, 101, 269–277. [Google Scholar]
- Puño-Sarmiento, J.; Medeiros, L.; Chiconi, C.; Martins, F.; Pelayo, J.; Rocha, S.; Blanco, J.; Blanco, M.; Zanutto, M.; Kobayashi, R.; et al. Detection of diarrheagenic Escherichia coli strains isolated from dogs and cats in Brazil. Vet. Microbiol. 2013, 166, 676–680. [Google Scholar]
- Lu, J.; Hofacre, C.L.; Lee, M.D. Emerging technologies in microbial ecology aid in understanding the effect of monoensin on necrotic enteritis. J. Appl. Poult. Res. 2006, 15, 145–153. [Google Scholar] [CrossRef]
- Mathew, A.G.; Cissell, R.; Liamthong, S. Antibiotic resistance in bacteria associated with food animals: A United States perspective of livestock production. Foodborne Pathog. Dis. 2007, 4, 115–133. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Dettman, B.; Beam, J.; Mix, C.; Jiang, X. Occurrence of ceftriaxone-resistant commensal bacteria on a dairy farm and a poultry farm. Can. J. Microbiol. 2006, 52, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Campagnolo, E.R.; Johnson, K.R.; Karpati, A.; Rubin, C.S.; Kolpin, D.W.; Meyer, M.T.; Esteban, J.E.; Currier, R.W.; Smith, K.; Thu, K.M.; et al. Antimicrobial residues in animal waste and water resources proximal to large-scale swine and poultry feeding operations. Sci. Total. Environ. 2002, 299, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Fairchild, A.S.; Smith, J.L.; Idris, U.; Lu, J.; Sanchez, S.; Purvis, L.B.; Hofacre, C.; Lee, M.D. Effects of orally administered tetracycline on the intestinal community structure of chickens and on tet determinant carriage by commensal bacteria and Campylobacter jejuni. Appl. Environ. Microbiol. 2005, 71, 5865–5872. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Drum, D.J.; Dai, Y.; Kim, J.M.; Sanchez, S.; Maurer, J.J.; Hofacre, C.L.; Lee, M.D. Impact of antimicrobial usage on antimicrobial resistance in commensal Escherichia coli strains colonizing broiler chickens. Appl. Environ. Microbiol. 2007, 73, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- FDA. Withdrawal of the new animal drug application for enrofloxacin in poultry; Document No. 2000N-1571; Department of Health and Human Services: Washington, DC, USA, 2005. [Google Scholar]
- Girardeau, J.P.; Dalmasso, A.; Bertin, Y.; Ducrot, C.; Bord, S.; Livrelli, V.; Vernozy-Rozand, C.; Martin, C. Association of virulence genotype with phylogenetic background in comparison to different seropathotypes of shiga toxin-producing Escherichia coli isolates. J. Clin. Microbiol. 2005, 43, 6098–6107. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Clermont, O.; Tolley, H.; Denamur, E. Assigning Escherichia coli strains to phylogenetic groups: Multi-locus sequence typing versus the pcr triplex method. Environ. Microbiol. 2008, 10, 2484–2496. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Paramo, P.; Clermont, O.; Blanc-Potard, A.B.; Bui, H.; Le Bouguenec, C.; Denamur, E. A specific genetic background is required for acquisition and expression of virulence factors in Escherichia coli. Mol. Biol. Evol. 2004, 21, 1085–1094. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Puño-Sarmiento, J.; Gazal, L.E.; Medeiros, L.P.; Nishio, E.K.; Kobayashi, R.K.T.; Nakazato, G. Identification of Diarrheagenic Escherichia coli Strains from Avian Organic Fertilizers. Int. J. Environ. Res. Public Health 2014, 11, 8924-8939. https://doi.org/10.3390/ijerph110908924
Puño-Sarmiento J, Gazal LE, Medeiros LP, Nishio EK, Kobayashi RKT, Nakazato G. Identification of Diarrheagenic Escherichia coli Strains from Avian Organic Fertilizers. International Journal of Environmental Research and Public Health. 2014; 11(9):8924-8939. https://doi.org/10.3390/ijerph110908924
Chicago/Turabian StylePuño-Sarmiento, Juan, Luis Eduardo Gazal, Leonardo P. Medeiros, Erick K. Nishio, Renata K. T. Kobayashi, and Gerson Nakazato. 2014. "Identification of Diarrheagenic Escherichia coli Strains from Avian Organic Fertilizers" International Journal of Environmental Research and Public Health 11, no. 9: 8924-8939. https://doi.org/10.3390/ijerph110908924