
Validation of Type 2 Diabetes Risk Variants Identified by Genome-Wide Association Studies in Northern Han Chinese



Abstract
:1. Introduction
2. Experimental Section
2.1. Study Participants
2.2. Selection of SNPs and Genotyping
2.3. Data Collection
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Participants and SNP Information
3.2. Association Analysis of the Candidate SNPs for T2DM
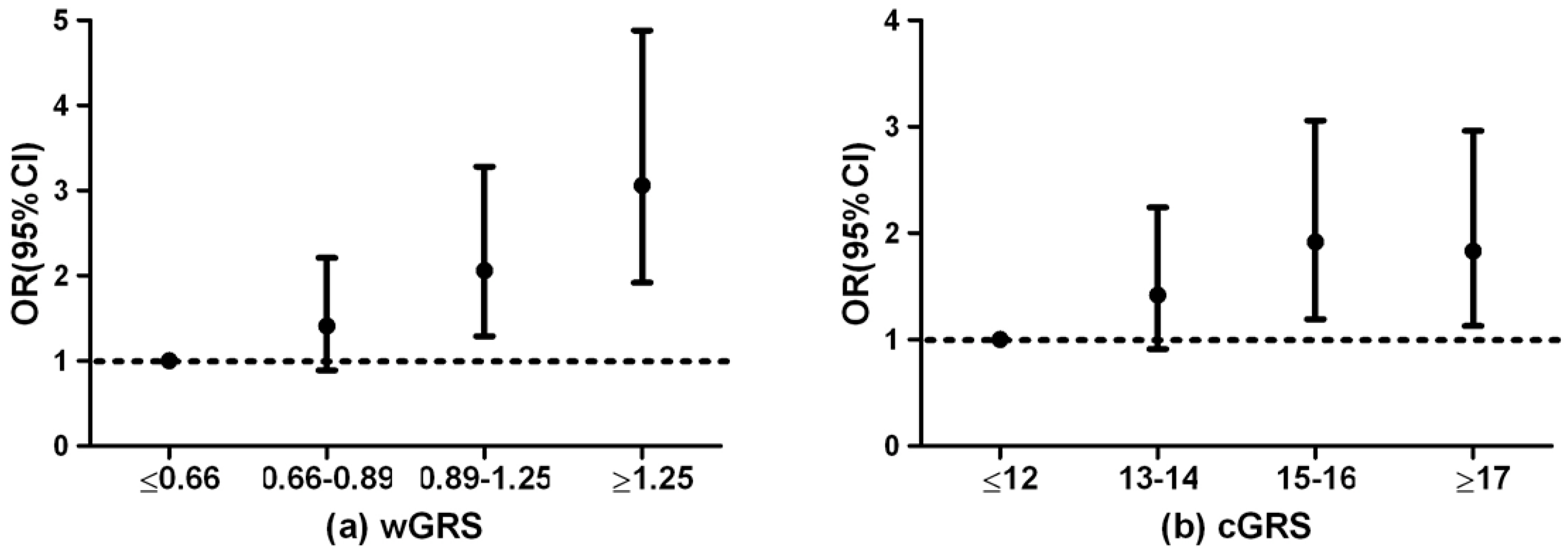
3.3. Genetic Risk Score and Diabetes Risk
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ginter, E.; Simko, V. Type 2 diabetes mellitus, pandemic in 21st century. Adv. Exp. Med. Biol. 2012, 771, 42–50. [Google Scholar] [PubMed]
- Xu, Y.; Wang, L.; He, J.; Bi, Y.; Li, M.; Wang, T.; Wang, L.; Jiang, Y.; Dai, M.; Lu, J.; et al. Prevalence and control of diabetes in Chinese adults. JAMA 2013, 310, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Almgren, P.; Lehtovirta, M.; Isomaa, B.; Sarelin, L.; Taskinen, M.R.; Lyssenko, V.; Tuomi, T.; Groop, L. Heritability and familiality of type 2 diabetes and related quantitative traits in the Botnia Study. Diabetologia 2011, 54, 2811–2819. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Chong, S.; Jalaludin, B.; Comino, E.; Bauman, A.E. Risk factors of incident type 2-diabetes mellitus over a 3-year follow-up, results from a large Australian sample. Diabetes Res. Clin. Pract. 2015, 108, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Voight, B.F.; Lyssenko, V.; Burtt, N.P.; de Bakker, P.I.; Chen, H.; Roix, J.J.; Kathiresan, S.; Hirschhorn, J.N.; Daly, M.J.; et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 2007, 316, 1331–1336. [Google Scholar] [PubMed]
- Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Shen, L.; Serre, D.; Boutin, P.; Vincent, D.; Belisle, A.; Hadjadj, S.; et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature 2007, 445, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Zeggini, E.; Weedon, M.N.; Lindgren, C.M.; Frayling, T.M.; Elliott, K.S.; Lango, H.; Timpson, N.J.; Perry, J.R.; Rayner, N.W.; Freathy, R.M.; et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science 2007, 316, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Omori, S.; Tanaka, Y.; Horikoshi, M.; Takahashi, A.; Hara, K.; Hirose, H.; Kashiwagi, A.; Kaku, K.; Kawamori, R.; Kadowaki, T.; et al. Replication study for the association of new meta-analysis-derived risk loci with susceptibility to type 2 diabetes in 6244 Japanese individuals. Diabetologia 2009, 52, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.J.; Yang, C.F.; Chen, C.C.; Chuang, L.M.; Lu, C.H.; Chang, C.T.; Wang, T.Y.; Chen, R.H.; Shiu, C.F.; Liu, Y.M.; et al. A genome-wide association study identifies susceptibility variants for type 2 diabetes in Han Chinese. PLoS Genet. 2010, 6, e1000847. [Google Scholar] [CrossRef] [PubMed]
- Onuma, H.; Tabara, Y.; Kawamoto, R.; Shimizu, I.; Kawamura, R.; Takata, Y.; Nishida, W.; Ohashi, J.; Miki, T.; Kohara, K.; et al. The GCKR rs780094 polymorphism is associated with susceptibility of type 2 diabetes, reduced fasting plasma glucose levels, increased triglycerides levels and lower HOMA-IR in Japanese population. J. Hum. Genet. 2010, 55, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Zhu, X.; Xu, M.; Guo, T.; Zhu, D.; Chen, G.; Li, X.; Xu, L.; Bi, Y.; Chen, Y.; et al. A genome-wide association study confirms previously reported loci for type 2 diabetes in Han Chinese. PLoS ONE 2011, 6, e22353. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Go, M.J.; Zhang, W.; Below, J.E.; Gaulton, K.J.; Ferreira, T.; Horikoshi, M.; Johnson, A.D.; Ng, M.C.; Prokopenko, I.; et al. Genome-wide trans-ancestry meta-analysis provides insight into the genetic architecture of type 2 diabetes susceptibility. Nat. Genet. 2014, 46, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Harder, M.N.; Appel, E.V.; Grarup, N.; Gjesing, A.P.; Ahluwalia, T.S.; Jørgensen, T.; Christensen, C.; Brandslund, I.; Linneberg, A.; Sørensen, T.I.; et al. The type 2 diabetes risk allele of TMEM154-rs6813195 associates with decreased beta cell function in a study of 6486 Danes. PLoS ONE 2015, 10, e120890. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.L.; Jacobs, S.B.; Moreno-Macías, H.; Huerta-Chagoya, A.; Churchhouse, C.; Márquez-Luna, C.; García-Ortíz, H.; Gómez-Vázquez, M.J.; Burtt, N.P.; Aguilar-Salinas, C.A.; et al. Sequence variants in SLC16A11 are a common risk factor for type 2 diabetes in Mexico. Nature 2013, 506, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Almawi, W.Y.; Nemr, R.; Keleshian, S.H.; Echtay, A.; Saldanha, F.L.; AlDoseri, F.A.; Racoubian, E. A replication study of 19 GWAS-validated type 2 diabetes at-risk variants in the Lebanese population. Diabetes Res. Clin. Pract. 2013, 102, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Liu, P.H.; Yu, Y.H.; Kuo, S.S.; Chang, T.J.; Jiang, Y.D.; Nong, J.Y.; Hwang, J.J.; Chuang, L.M. Validation of type 2 diabetes risk variants identified by genome-wide association studies in Han Chinese population, a replication study and meta-analysis. PLoS ONE 2014, 9, e95045. [Google Scholar] [CrossRef] [PubMed]
- Narayan, K.M.; Weber, M.B. Clinical risk factors, DNA variants, and the development of type 2 diabetes. N. Engl. J. Med. 2009, 360, 1360–1361. [Google Scholar] [PubMed]
- Meigs, J.B.; Shrader, P.; Sullivan, L.M.; McAteer, J.B.; Fox, C.S.; Dupuis, J.; Manning, A.K.; Florez, J.C.; Wilson, P.W.; D’Agostino, R.B., Sr.; et al. Genotype score in addition to common risk factors for prediction of type 2 diabetes. N. Engl. J. Med. 2008, 359, 2208–2219. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Delahanty, R.; Gao, Y.T.; Long, J.; Williams, S.M.; Xiang, Y.B.; Cai, H.; Li, H.L.; Hu, F.; Cai, Q.; et al. Joint effect of genetic and lifestyle risk factors on type 2 diabetes risk among Chinese men and women. PLoS ONE 2012, 7, e49464. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Wang, H.; Fang, H.H.; Gao, Q.; Zhang, J.; Song, M.S.; Zhou, Y.; Wang, Y.X.; Wang, W. Association between IGF2BP2 polymorphisms and type 2 diabetes mellitus: A case-control study and meta-analysis. Int. J. Environ. Res. Public Health 2016, 13, 574. [Google Scholar] [CrossRef] [PubMed]
- Genuth, S.; Alberti, K.G.; Bennett, P.; Buse, J.; Defronzo, R.; Kahn, R.; Kitzmiller, J.; Knowler, W.C.; Lebovitz, H.; Lernmark, A.; et al. Follow-up report on the diagnosis of diabetes mellitus. Diabetes Care 2003, 26, 3160–3167. [Google Scholar] [PubMed]
- The HapMap-CHB Data. Available online: http://hapmap.ncbi.nlm.nih.gov/ (accessed on 25 Augut 2016).
- Wei, W.; Xin, X.; Shao, B.; Zeng, F.; Love, E.J.; Wang, B. The relationship between anthropometric indices and type 2 diabetes mellitus among adults in North-East China. Public Health Nutr. 2015, 18, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes, results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Matharoo, K.; Sharma, R.; Bhanwer, A.J. C-reactive protein + 1059 G>C polymorphism in type 2 diabetes and coronary artery disease patients. Meta Gene 2013, 1, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Sikka, R.; Raina, P.; Matharoo, K.; Bandesh, K.; Bhatia, R.; Chakrabarti, S.; Bhanwer, A.J. TNF-alpha (g. − 308 G > A) and ADIPOQ (g. + 45 T > G) gene polymorphisms in type 2 diabetes and microvascular complications in the region of Punjab (North-West India). Curr. Eye Res. 2014, 39, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.W.; Kim, H.S.; Yoon, S.K.; Murty, V.V.; Kim, E. Human Fas associated factor 1, hFAF1, gene maps to chromosome band 1p32. Mol. Cells 2000, 10, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Park, M.Y.; Moon, J.H.; Lee, K.S.; Choi, H.I.; Chung, J.; Hong, H.J.; Kim, E. FAF1 suppresses IkappaB kinase (IKK) activation by disrupting the IKK complex assembly. J. Biol. Chem. 2007, 282, 27572–27577. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Yuan, M.; Frantz, D.F.; Melendez, P.A.; Hansen, L.; Lee, J.; Shoelson, S.E. Local and systemic insulin resistance resulting from hepatic activation of IKK-beta and NF-kappaB. Nat. Med. 2005, 11, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Harte, A.L.; Tripathi, G.; Piya, M.K.; Barber, T.M.; Clapham, J.C.; Al-Daghri, N.; Al-Disi, D.; Kumsaiyai, W.; Saravanan, P.; Fowler, A.E.; et al. NF kappaB as a potent regulator of inflammation in human adipose tissue, influenced by depot, adiposity, T2DM status, and TNF alpha. Obesity (Silver Spring) 2013, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Park, C.S.; Hwang, M.; Nam, H.Y.; Chang, H.S.; Park, S.G.; Han, B.G.; Kimm, K.; Kim, H.L.; Oh, B.; et al. A common intronic variant of CXCR3 is functionally associated with gene expression levels and the polymorphic immune cell responses to stimuli. J. Allergy Clin. Immunol. 2008, 122, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.A.; Boucher, G.; Lees, C.W.; Franke, A.; D’Amato, M.; Taylor, K.D.; Lee, J.C.; Goyette, P.; Imielinski, M.; Latiano, A.; et al. Meta-analysis identifies 29 additional ulcerative colitis risk loci, increasing the number of confirmed associations to 47. Nat. Genet. 2011, 43, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Corfe, M.; Majumdar, D.; Assadsangabi, A.; Marsh, A.M.; Cross, S.S.; Connolly, J.B.; Evans, C.A.; Lobo, A.J. Inflammation decreases keratin level in ulcerative colitis, inadequate restoration associates with increased risk of colitis-associated cancer. BMJ Open Gastroenterol. 2015, 2, e000024. [Google Scholar] [CrossRef] [PubMed]
- Fumery, M.; Duricova, D.; Gower-Rousseau, C.; Annese, V.; Peyrin-Biroulet, L.; Lakatos, P.L. Review article, the natural history of paediatric-onset ulcerative colitis in population-based studies. Aliment. Pharmacol. Ther. 2016, 43, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa-Kobayashi, K.; Natsume, M.; Aoki, S.; Nakano, S.; Inamori, T.; Kasezawa, N.; Goda, T. The combined effect of the T2DM susceptibility genes is an important risk factor for T2DM in non-obese Japanese, a population based case-control study. BMC Med. Genet. 2012, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Shigemizu, D.; Tsunoda, T.; Iwata, M.; Maegawa, H.; Watada, H.; Hirose, H.; Tanaka, Y.; Tobe, K.; Kaku, K.; et al. Assessing the clinical utility of a genetic risk score constructed using 49 susceptibility alleles for type 2 diabetes in a Japanese population. J. Clin. Endocrinol. Metab. 2013, 98, e1667–e1673. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Qi, L.; Zhang, C.; Kraft, P.; Manson, J.; Cai, T.; Hunter, D.J.; Hu, F.B. Joint effects of common genetic variants on the risk for type 2 diabetes in U.S. men and women of European ancestry. Ann. Intern. Med. 2009, 150, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, G.; Spurgeon, C.J.; Tabassum, R.; Bhaskar, S.; Kulkarni, S.R.; Mahajan, A.; Chavali, S.; Kumar, M.V.; Prakash, S.; Dwivedi, O.P.; et al. Impact of common variants of PPARG, KCNJ11, TCF7L2, SLC30A8, HHEX, CDKN2A, IGF2BP2, and CDKAL1 on the risk of type 2 diabetes in 5164 Indians. Diabetes 2010, 59, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Tabara, Y.; Osawa, H.; Kawamoto, R.; Onuma, H.; Shimizu, I.; Makino, H.; Kohara, K.; Miki, T. Genotype risk score of common susceptible variants for prediction of type 2 diabetes mellitus in Japanese, the Shimanami Health Promoting Program (J-SHIPP study). Development of type 2 diabetes mellitus and genotype risk score. Metabolism 2011, 60, 1634–1640. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| No. | SNP rs# | Gene | Chr. | Chr. Position | Risk/Non-RiskAllele | HW-P | Call Rate | |
|---|---|---|---|---|---|---|---|---|
| Case | Control | |||||||
| 1 | rs17106184 | FAF1 | 1 | 50444313 | A/G | 1.00 | 0.24 | 98.2 |
| 2 | rs780094 | GCKR | 2 | 27518370 | A/G | 0.78 | 1.00 | 98.9 |
| 3 | rs3773159 | MGLL | 3 | 127720095 | T/C | 1.00 | 0.79 | 98.9 |
| 4 | rs4402960 | IGF2BP2 | 3 | 185793899 | T/G | 0.21 | 0.09 | 98.1 |
| 5 | rs1470579 | IGF2BP2 | 3 | 185811292 | C/A | 0.19 | 0.05 | 98.1 |
| 6 | rs702634 | ARL15 | 5 | 53975590 | A/G | 1.00 | 0.81 | 99.1 |
| 7 | rs4712523 | CDKAL1 | 6 | 20657333 | A/G | 0.50 | 0.2 | 98.5 |
| 8 | rs4712524 | CDKAL1 | 6 | 20657634 | A/G | 0.85 | 1.00 | 98.2 |
| 9 | rs10946398 | CDKAL1 | 6 | 20660803 | A/C | 0.70 | 0.38 | 98.6 |
| 10 | rs7756992 | CDKAL1 | 6 | 20679478 | A/G | 0.09 | 1.00 | 98.1 |
| 11 | rs3130501 | POU5F1-TCF19 | 6 | 31168676 | G/A | 0.50 | 0.18 | 99.4 |
| 12 | rs9472138 | VEGFA | 6 | 43844025 | C/T | 0.02 | 0.04 | 99.6 |
| 13 | rs864745 | JAZF1 | 7 | 28140937 | G/A | 0.48 | 0.89 | 99.0 |
| 14 | rs13266634 | SLC30A8 | 8 | 117172544 | T/C | 0.05 | 0.49 | 98.0 |
| 15 | rs10811661 | CDKN2B | 9 | 22134095 | T/C | 0.57 | 0.29 | 98.8 |
| 16 | rs12779790 | CDC123/CAMKID | 10 | 12286011 | A/G | 0.71 | 0.47 | 99.3 |
| 17 | rs1111875 | HHEX | 10 | 92703125 | A/G | 0.57 | 0.008 | 99.0 |
| 18 | rs7923837 | HHEX | 10 | 92722160 | G/A | 0.17 | 0.58 | 98.6 |
| 19 | rs7903146 | TCF7L2 | 10 | 112998590 | T/C | 1.00 | 1.00 | 98.6 |
| 20 | rs1153188 | DCD | 12 | 54705212 | A/T | 1.00 | 1.00 | 99.0 |
| 21 | rs1370176 | C2CD4A/B | 15 | 62105035 | T/C | 1.00 | 0.91 | 98.8 |
| 22 | rs1436953 | C2CD4A/B | 15 | 62121815 | A/G | 0.84 | 0.83 | 98.8 |
| 23 | rs8050136 | FTO | 16 | 53782363 | A/C | 0.83 | 0.07 | 97.8 |
| 24 | rs7192960 | MAF/WWOX | 16 | 79382666 | T/C | 1.00 | 1.00 | 99.8 |
| 25 | rs75493593 | SLC16A11 | 17 | 7041768 | T/G | 1.00 | 0.82 | 98.5 |
| 26 | rs75418188 | SLC16A11 | 17 | 7042164 | T/C | 1.00 | 0.66 | 98.5 |
| 27 | rs13342232 | SLC16A11 | 17 | 7042621 | G/A | 1.00 | 0.66 | 98.0 |
| 28 | rs13342692 | SLC16A11 | 17 | 7042968 | C/T | 1.00 | 1.00 | 99.0 |
| 29 | rs117767867 | SLC16A11 | 17 | 7043011 | T/C | 1.00 | 1.00 | 99.0 |
| No. | SNP rs# | Gene | Frequency of Risk Allele | Crude Model | Adjusted Model * | |||
|---|---|---|---|---|---|---|---|---|
| Case | Control | OR (95% CI) | p | OR (95% CI) | p | |||
| 1 | rs17106184 | FAF1 | 0.12 | 0.06 | 1.91 (1.36–2.69) | <0.0001 | 2.22 (1.53–3.24) | <0.0001 |
| 2 | rs780094 | GCKR | 0.53 | 0.52 | 0.96 (0.80–1.17) | 0.71 | 0.95 (0.77–1.17) | 0.60 |
| 3 | rs3773159 | MGLL | 0.12 | 0.10 | 1.16 (0.86–1.56) | 0.33 | 1.04 (0.75–1.45) | 0.82 |
| 4 | rs4402960 | IGF2BP2 | 0.25 | 0.25 | 1.02 (0.82–1.26) | 0.89 | 1.05 (0.83–1.34) | 0.67 |
| 5 | rs1470579 | IGF2BP2 | 0.27 | 0.26 | 1.04 (0.84–1.28) | 0.75 | 1.07 (0.85–1.36) | 0.55 |
| 6 | rs702634 | ARL15 | 0.90 | 0.89 | 0.89 (0.66–1.20) | 0.45 | 0.77 (0.55–1.08) | 0.13 |
| 7 | rs4712523 | CDKAL1 | 0.57 | 0.55 | 0.92 (0.77–1.11) | 0.41 | 0.84 (0.69–1.04) | 0.11 |
| 8 | rs4712524 | CDKAL1 | 0.57 | 0.55 | 0.91 (0.75–1.11) | 0.33 | 0.84 (0.68–1.03) | 0.10 |
| 9 | rs10946398 | CDKAL1 | 0.57 | 0.56 | 0.94 (0.78–1.14) | 0.55 | 0.87 (0.71–1.07) | 0.18 |
| 10 | rs7756992 | CDKAL1 | 0.49 | 0.48 | 1.06 (0.88–1.28) | 0.55 | 1.13 (0.92–1.39) | 0.26 |
| 11 | rs3130501 | POU5F1-TCF19 | 0.70 | 0.68 | 0.91 (0.74–1.11) | 0.35 | 0.81 (0.64–1.01) | 0.06 |
| 12 | rs9472138 | VEGFA | 0.91 | 0.90 | 0.88 (0.64–1.20) | 0.41 | 0.75 (0.53–1.07) | 0.11 |
| 13 | rs864745 | JAZF1 | 0.27 | 0.24 | 1.17 (0.94–1.45) | 0.16 | 1.13 (0.89–1.43) | 0.32 |
| 14 | rs13266634 | SLC30A8 | 0.42 | 0.42 | 1.02 (0.85–1.24) | 0.81 | 0.96 (0.78–1.18) | 0.68 |
| 15 | rs10811661 | CDKN2B | 0.54 | 0.52 | 0.91 (0.75–1.09) | 0.31 | 0.88 (0.71–1.08) | 0.21 |
| 16 | rs12779790 | CDC123/CAMKID | 0.85 | 0.84 | 0.89 (0.69–1.16) | 0.39 | 0.83 (0.62–1.11) | 0.20 |
| 17 | rs7923837 | HHEX | 0.78 | 0.77 | 0.96 (0.76–1.20) | 0.69 | 1.01 (0.79–1.30) | 0.91 |
| 18 | rs7903146 | TCF7L2 | 0.09 | 0.09 | 1.01 (0.63–1.62) | 0.97 | 1.05 (0.63–1.74) | 0.86 |
| 19 | rs1153188 | DCD | 0.09 | 0.09 | 1.21 (0.60–2.45) | 0.60 | 1.32 (0.58–2.96) | 0.51 |
| 20 | rs1370176 | C2CD4A/B | 0.30 | 0.29 | 1.02 (0.83–1.25) | 0.88 | 0.98 (0.78–1.23) | 0.87 |
| 21 | rs1436953 | C2CD4A/B | 0.36 | 0.35 | 1.06 (0.87–1.28) | 0.59 | 1.02 (0.82–1.27) | 0.85 |
| 22 | rs8050136 | FTO | 0.13 | 0.11 | 1.26 (0.94–1.69) | 0.12 | 1.16 (0.84–1.60) | 0.37 |
| 23 | rs7192960 | MAF/WWOX | 0.29 | 0.29 | 1.06 (0.82–1.24) | 0.97 | 1.07 (0.85–1.34) | 0.59 |
| 24 | rs75493593 | SLC16A11 | 0.13 | 0.12 | 1.13 (0.85–1.49) | 0.41 | 1.14 (0.84–1.56) | 0.39 |
| 25 | rs75418188 | SLC16A11 | 0.13 | 0.12 | 1.11 (0.84–1.47) | 0.46 | 1.23 (0.83–1.53) | 0.45 |
| 26 | rs13342232 | SLC16A11 | 0.14 | 0.12 | 1.11 (0.83–1.45) | 0.51 | 1.11 (0.82–1.52) | 0.49 |
| 27 | rs13342692 | SLC16A11 | 0.13 | 0.13 | 1.08 (0.82–1.43) | 0.58 | 1.09 (0.80–1.48) | 0.58 |
| 28 | rs117767867 | SLC16A11 | 0.13 | 0.13 | 1.06 (0.81–1.40) | 0.67 | 1.06 (0.78–1.44) | 0.71 |
| rs17106184 | Healthy Controls (N = 419) | T2DM Patients (N = 460) | Crude Model | Adjusted Model * | ||
|---|---|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) * | p | |||
| Allele (%) | ||||||
| G | 784 (93.6) | 813 (88.3) | Ref. | Ref. | ||
| A | 54 (6.4) | 107 (11.7) | 1.91 (1.36–2.69) | <0.0001 | 2.22 (1.53–3.24) | <0.0001 |
| Genotype (%) | ||||||
| GG | 368 (87.8) | 359 (78) | Ref. | Ref. | - | |
| AG | 48 (11.5) | 95 (20.7) | 2.03 (1.39–2.96) | <0.0001 | 2.30 (1.53–3.52) | <0.0001 |
| AA | 3 (0.7) | 6 (1.3) | 2.05 (0.51–7.36) | 0.310 | 2.54 (0.70–11.03) | 0.130 |
| Dominant model (%) | ||||||
| GG | 368 (87.8) | 359 (78) | Ref. | Ref. | ||
| AA/AG | 51 (12.2) | 101 (22) | 2.03 (1.41–2.93) | 1.54 × 10−4 | 2.32 (1.57–3.47) | 4.61 × 10−5 |
| Recessive model (%) | ||||||
| GG/AG | 416 (99.3) | 454 (98.7) | Ref. | Ref. | ||
| AA | 3 (0.7) | 6 (1.3) | 1.85 (0.46–7.46) | 0.390 | 2.41 (0.63–10.12) | 0.170 |
| Additive model (%) | ||||||
| GG:GA:AA | - | - | 2.07 (1.34–3.02) | <0.0001 | 2.14 (1.47–3.12) | 7.96 × 10−5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, P.; Zhou, Y.; Ge, S.-Q.; Wang, A.-X.; Yu, X.-W.; Alzain, M.A.; Veronica, A.K.; Qiu, J.; Song, M.-S.; Zhang, J.; et al. Validation of Type 2 Diabetes Risk Variants Identified by Genome-Wide Association Studies in Northern Han Chinese. Int. J. Environ. Res. Public Health 2016, 13, 863. https://doi.org/10.3390/ijerph13090863
Rao P, Zhou Y, Ge S-Q, Wang A-X, Yu X-W, Alzain MA, Veronica AK, Qiu J, Song M-S, Zhang J, et al. Validation of Type 2 Diabetes Risk Variants Identified by Genome-Wide Association Studies in Northern Han Chinese. International Journal of Environmental Research and Public Health. 2016; 13(9):863. https://doi.org/10.3390/ijerph13090863
Chicago/Turabian StyleRao, Ping, Yong Zhou, Si-Qi Ge, An-Xin Wang, Xin-Wei Yu, Mohamed Ali Alzain, Andrea Katherine Veronica, Jing Qiu, Man-Shu Song, Jie Zhang, and et al. 2016. "Validation of Type 2 Diabetes Risk Variants Identified by Genome-Wide Association Studies in Northern Han Chinese" International Journal of Environmental Research and Public Health 13, no. 9: 863. https://doi.org/10.3390/ijerph13090863
APA StyleRao, P., Zhou, Y., Ge, S.-Q., Wang, A.-X., Yu, X.-W., Alzain, M. A., Veronica, A. K., Qiu, J., Song, M.-S., Zhang, J., Wang, H., Fang, H.-H., Gao, Q., Wang, Y.-X., & Wang, W. (2016). Validation of Type 2 Diabetes Risk Variants Identified by Genome-Wide Association Studies in Northern Han Chinese. International Journal of Environmental Research and Public Health, 13(9), 863. https://doi.org/10.3390/ijerph13090863