
Predicting Avian Influenza Co-Infection with H5N1 and H9N2 in Northern Egypt

 , ,
, , 
Abstract
:
1. Introduction
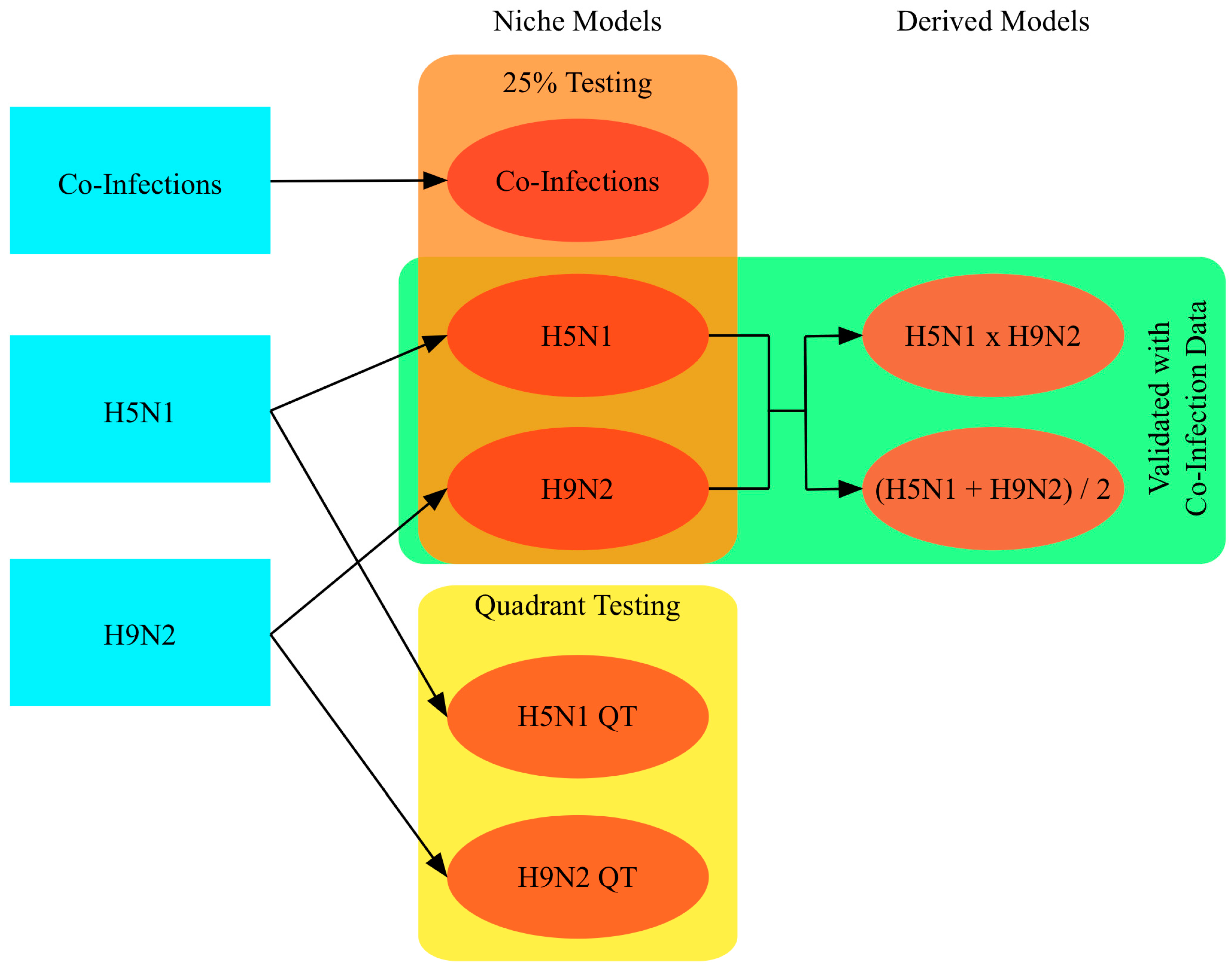
2. Materials and Methods
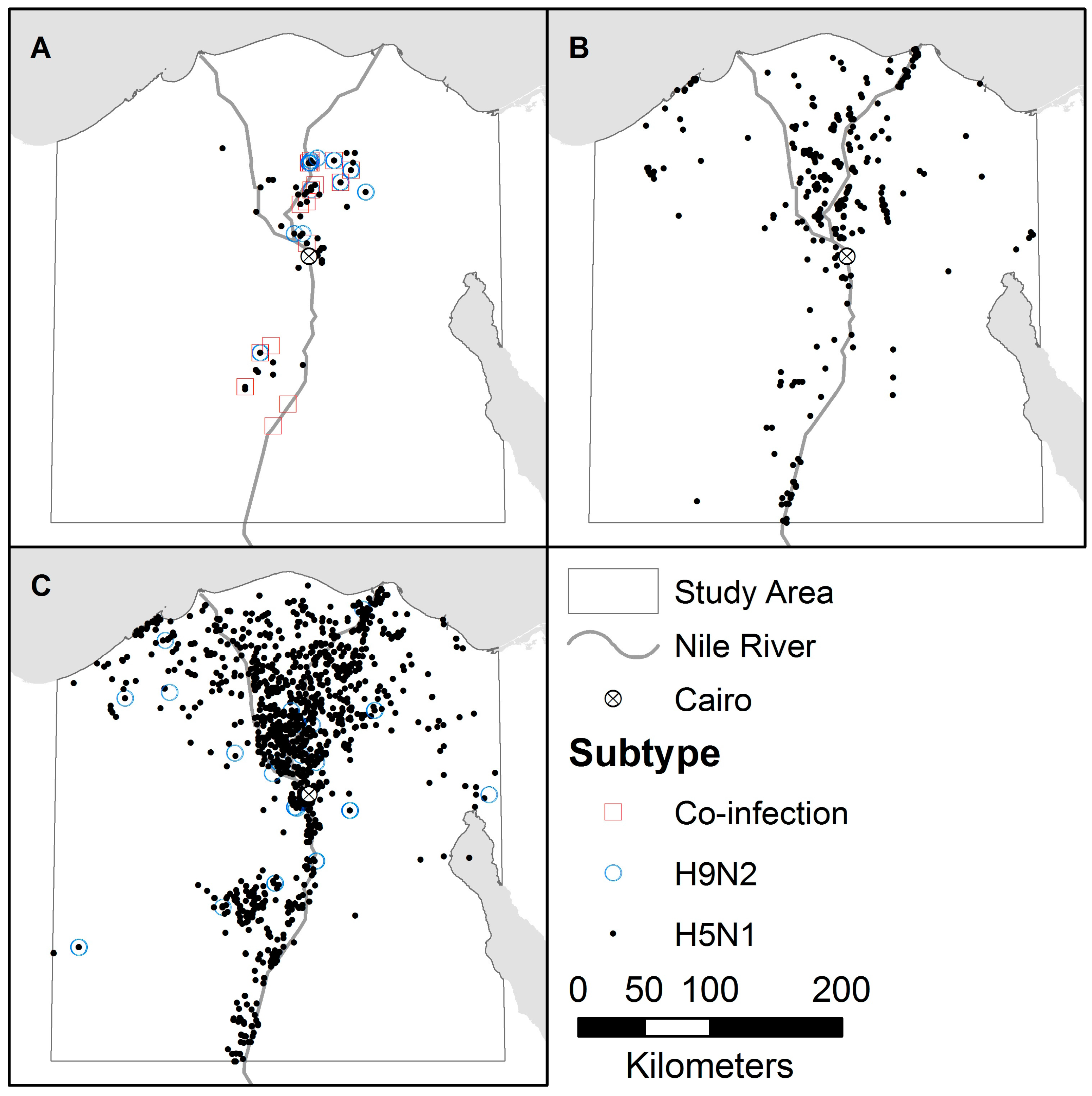
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peiris, J.S.M.; de Jong, M.D.; Guan, Y. Avian influenza virus (H5N1): A threat to human health. Clin. Microbiol. Rev. 2007, 20, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.J.D.; Naipospos, T.S.P.; Nguyen, T.D.; de Jong, M.D.; Vijaykrishna, D.; Usman, T.B.; Hassan, S.S.; Nguyen, T.V.; Dao, T.V.; Bui, N.A.; et al. Evolution and adaptation of H5N1 influenza virus in avian and human hosts in Indonesia and Vietnam. Virology 2006, 350, 258–268. [Google Scholar] [CrossRef] [PubMed]
- El-Zoghby, E.F.; Aly, M.M.; Nasef, S.A.; Hassan, M.K.; Arafa, A.-S.; Selim, A.A.; Kholousy, S.G.; Kilany, W.H.; Safwat, M.; Abdelwhab, E.M.; et al. Surveillance on A/H5N1 virus in domestic poultry and wild birds in Egypt. Virol. J. 2013, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Skeik, N.; Jabr, F.I. Influenza viruses and the evolution of avian influenza virus H5N1. Int. J. Infect. Dis. 2008, 12, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Kawaoka, Y. Host range restriction and pathogenicity in the context of influenza pandemic. Emerg. Infect. Dis. 2006, 12, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Bai, T.; Li, X.; Xiong, Y.; Huang, Y.; Pan, M.; Zhang, Y.; Bo, H.; Zou, S.; Shu, Y. The comparison of pathology in ferrets infected by H9N2 avian influenza viruses with different genomic features. Virology 2016, 488, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2014, 112, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Negovetich, N.J.; Forrest, H.L.; Webster, R.G. Ducks: The “Trojan horses” of H5N1 influenza. Influenza Other Respir. Viruses 2009, 3, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.; Croisier, A. Transmission of avian influenza viruses to and between humans. J. Infect. Dis. 2005, 192, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Taha, F.A. Poultry Sector in Middle-Income Countries and Its Feed Requirements: The Case of Egypt; United States Department of Agriculture, Economic Research Service: Washington, DC, USA, 2003; p. 47.
- Gomaa, M.R.; Kayed, A.S.; Elabd, M.A.; Zeid, D.A.; Zaki, S.A.; El Rifay, A.S.; Sherif, L.S.; McKenzie, P.P.; Webster, R.G.; Webby, R.J.; et al. Avian influenza A(H5N1) and A(H9N2) seroprevalence and risk factors for infection among Egyptians: A prospective, controlled seroepidemiological study. J. Infect. Dis. 2015, 211, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Central Intelligence Agency. Egypt. In The World Factbook 2014–2015; Central Intelligence Agency: Washington, DC, USA, 2014. [Google Scholar]
- El-Zanaty, F.; Way, A. Egypt Demographic and Health Survey 2008; MEASURE DHS; Ministry of Health, El-Zanaty and Associates, and Macro International: Cairo, Egypt, 2009; p. 463.
- Aly, M.M.; Arafa, A.; Hassan, M.K. Epidemiological findings of outbreaks of disease caused by highly pathogenic H5N1 avian influenza virus in poultry in Egypt during 2006. Avian Dis. 2008, 52, 269–277. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cumulative Number of Confirmed Human Cases for Avian Influenza A(H5N1) Reported to WHO, 2003–2015. Available online: http://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/ (accessed on 18 July 2016).
- World Organisation for Animal Health (OIE). Outbreaks of Highly Pathogenic Avian Influenza (Subtype H5N1) in Poultry Notified to the OIE from the End of 2003 to 22 October 2015. Available online: http://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/graph_avian_influenza/graphs_HPAI_22_10_2015.pdf (accessed on 18 July 2016).
- Kandeel, A.; Manoncourt, S.; El Kareem, E.A.; Ahmed, A.-N.M.; El-Refaie, S.; Essmat, H.; Tjaden, J.; de Mattos, C.C.; Earhart, K.C.; Marfin, A.A.; et al. Zoonotic transmission of avian influenza virus (H5N1), Egypt, 2006–2009. Emerg. Infect. Dis. 2010, 16, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Arafa, A.-S.; Hagag, N.; Erfan, A.; Mady, W.; El-Husseiny, M.; Adel, A.; Nasef, S. Complete genome characterization of avian influenza virus subtype H9N2 from a commercial quail flock in Egypt. Virus Genes 2012, 45, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Kayali, G.; Kandeil, A.; El-Shesheny, R.; Kayed, A.S.; Gomaa, M.M.; Maatouq, A.M.; Shehata, M.M.; Moatasim, Y.; Bagato, O.; Cai, Z.; et al. Active surveillance for avian influenza virus, Egypt, 2010–2012. Emerg. Infect. Dis. 2014, 20, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J. Predictive vegetation mapping: Geographic modelling of biospatial patterns in relation to environmental gradients. Prog. Phys. Geogr. 1995, 19, 474–499. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Getting the concepts right. Nat. Conserv. 2012, 10, 102–107. [Google Scholar] [CrossRef]
- Warren, D.L. In defense of “niche modeling”. Trends Ecol. Evol. 2012, 27, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- McInerny, G.J.; Etienne, R.S. “Niche” or “distribution” modelling? A response to Warren. Trends Ecol. Evol. 2013, 28, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Bodbyl-Roels, S.; Peterson, A.T.; Xiao, X. Comparative analysis of remotely-sensed data products via ecological niche modeling of avian influenza case occurrences in Middle Eastern poultry. Int. J. Health Geogr. 2011, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.A.; Byler, D.; Eastman, J.R.; Fleishman, E.; Geller, G.; Goetz, S.; Guild, L.; Hamilton, H.; Hansen, M.; Headley, R.; et al. Ten ways remote sensing can contribute to conservation. Conserv. Biol. 2015, 29, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Kayali, G.; Kandeil, A.; El-Shesheny, R.; Kayed, A.S.; Maatouq, A.M.; Cai, Z.; McKenzie, P.P.; Webby, R.J.; El Refaey, S.; Kandeel, A.; et al. Avian influenza A(H5N1) virus in Egypt. Emerg. Infect. Dis. 2016, 22, 379–388. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health (OIE). Available online: http://www.oie.int/ (accessed on 18 July 2016).
- Global Animal Disease Information System (EMPRES-i) of the Food and Agriculture Organization of the United Nations (FAO). Emergency Prevention System against Transboundary Animal and Plant Pests and Diseases (EMPRES). Available online: http://empres-i.fao.org/eipws3g/ (accessed on 18 July 2016).
- Moriguchi, S.; Onuma, M.; Goka, K. Potential risk map for avian influenza A virus invading Japan. Divers. Distrib. 2013, 19, 78–85. [Google Scholar] [CrossRef]
- Williams, R.A.; Peterson, A.T. Ecology and geography of avian influenza (HPAI H5N1) transmission in the Middle East and Northeastern Africa. Int. J. Health Geogr. 2009, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Chettri, A.; Barik, S.K. Modelling the ecology and distribution of highly pathogenic avian influenza (H5N1) in the Indian subcontinent. Curr. Sci. 2009, 97, 73–78. [Google Scholar]
- Si, Y.; Wang, T.; Skidmore, A.K.; de Boer, W.F.; Li, L.; Prins, H.H. Environmental factors influencing the spread of the highly pathogenic avian influenza H5N1 virus in wild birds in Europe. Ecol. Soc. 2010, 15, 26. [Google Scholar]
- Shaman, J.; Kohn, M. Absolute humidity modulates influenza survival, transmission, and seasonality. Proc. Natl. Acad. Sci. USA 2009, 106, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.-Q.; de Vlas, S.J.; Liang, S.; Looman, C.W.N.; Gong, P.; Xu, B.; Yan, L.; Yang, H.; Richardus, J.H.; Cao, W.-C. Environmental factors contributing to the spread of H5N1 avian influenza in mainland China. PLoS ONE 2008, 3, e2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.C.; Gilbert, M.; Desvaux, S.; Rasamoelina Andriamanivo, H.; Peyre, M.; Khong, N.V.; Thanapongtharm, W.; Chevalier, V. Agro-environmental determinants of avian influenza circulation: A multisite study in Thailand, Vietnam and Madagascar. PLoS ONE 2014, 9, e101958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhamis, M.A.; Hijmans, R.J.; Al-Enezi, A.; Martínez-López, B.; Perez, A.M. The use of spatial and spatio-temporal modeling for surveillance of H5N1 highly pathogenic avian influenza in poultry in the Middle East. Avian Dis. 2016, 60 (Suppl. 1), 146–155. [Google Scholar] [CrossRef] [PubMed]
- NASA Land Processes Distributed Active Archive Center (LP DAAC). Vegetation Indices Monthly L3 Global 1 km. Available online: https://lpdaac.usgs.gov/dataset_discovery/modis/modis_products_table/mod13a3 (accessed on 18 July 2016).
- Reperant, L.A.; Fučkar, N.S.; Osterhaus, A.D.M.E.; Dobson, A.P.; Kuiken, T. Spatial and temporal association of outbreaks of H5N1 influenza virus infection in wild birds with the 0 °C isotherm. PLoS Pathog. 2010, 6, e1000854. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.; Xiao, X.; Pfeiffer, D.U.; Epprecht, M.; Boles, S.; Czarnecki, C.; Chaitaweesub, P.; Kalpravidh, W.; Minh, P.Q.; Otte, M.J.; et al. Mapping H5N1 highly pathogenic avian influenza risk in Southeast Asia. Proc. Natl. Acad. Sci. USA 2008, 105, 4769–4774. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Di, B.; Zhou, D.-H.; Zheng, B.-J.; Jing, H.; Lin, Y.-P.; Liu, Y.-F.; Wu, X.-W.; Qin, P.-Z.; Wang, Y.-L.; et al. Food markets with live birds as source of avian influenza. Emerg. Infect. Dis. 2006, 12, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Cumming, G.S. Risk mapping for avian influenza: A social-ecological problem. Ecol. Soc. 2010, 15, 32. [Google Scholar]
- Defense Mapping Agency. Digital Chart of the World 1992. Available online: http://www.diva-gis.org/gdata (accessed on 18 July 2016).
- OpenStreetMap. OpenStreetMap Foundation. Available online: http://www.openstreetmap.org/ (accessed on 18 July 2016).
- WorldPop. Available online: http://www.worldpop.org.uk/ (accessed on 18 July 2016).
- Robinson, T.P.; Franceschini, G.; Wint, W. The Food and Agriculture Organization’s Gridded Livestock of the World. Vet. Ital. 2007, 43, 745–751. [Google Scholar] [PubMed]
- Gething, P.; Tatem, A.; Bird, T.; Burgert-Brucker, C.R. Creating Spatial Interpolation Surfaces with DHS Data; United States Agency for International Development (USAID): Rockville, MD, USA, 2015; p. 104.
- Pulliam, H.R. On the relationship between niche and distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecologic niche modeling and spatial patterns of disease transmission. Emerg. Infect. Dis. 2006, 12, 1822. [Google Scholar] [CrossRef] [PubMed]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Levine, R.S.; Peterson, A.T.; Yorita, K.L.; Carroll, D.; Damon, I.K.; Reynolds, M.G. Ecological niche and geographic distribution of human Monkeypox in Africa. PLoS ONE 2007, 2, e176. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.L.; Gilbert, M.; Martin, V.; Cappelle, J.; Hosseini, P.; Njabo, K.Y.; Abdel Aziz, S.; Xiao, X.; Daszak, P.; Smith, T.B. Predicting hotspots for influenza virus reassortment. Emerg. Infect. Dis. 2013, 19, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heibl, C.; Calenge, C. Phyloclim: Integrating Phylogenetics and Climatic Niche Modeling. Available online: http://CRAN.R-project.org/package=phyloclim (accessed on 18 July 2016).
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, T.; Li, X.; Jiang, Z.; Jiang, Q.; Chen, Q.; Tu, X.; Chen, Z.; Chang, J.; Li, L.; Xu, B. Serological evidence of H7, H5 and H9 avian influenza virus co-infection among herons in a city park in Jiangxi, China. Sci. Rep. 2014, 4, 6345. [Google Scholar] [CrossRef] [PubMed]
- Wisedchanwet, T.; Wongpatcharachai, M.; Boonyapisitsopa, S.; Bunpapong, N.; Jairak, W.; Kitikoon, P.; Sasipreeyajun, J.; Amonsin, A. Influenza A Virus Surveillance in Live-Bird Markets: First Report of Influenza A Virus Subtype H4N6, H4N9, and H10N3 in Thailand. Avian Dis. 2011, 55, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, M.; Arenas, M.; Galán, J.C.; Palero, F.; González-Candelas, F. Recombination in viruses: Mechanisms, methods of study, and evolutionary consequences. Infect. Genet. Evol. 2015, 30, 296–307. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Potential Relevance | Spatial Resolution | Timeframe | Source |
|---|---|---|---|---|
| Relative Humidity | Viral Persistence and Transmission [34] | 5 arcminutes | 1960–1990 | Center for Sustainability and the Global Environment (SAGE) Atlas of the Biosphere (AoB) |
| Mean Air Temp | Viral Persistence [30] | 30 arcseconds | 1960–1990 | WORLDCLIM |
| Total Precipitation | Viral Persistence [30] | 30 arcseconds | 1960–1990 | WORLDCLIM |
| Mean Diurnal Surface Temp | Viral Persistence and Diffusion [33,39] | 3 arcminutes | 2009–2014 | Moderate Resolution Imaging Spectroradiometer (MODIS) |
| Normalized Difference Vegetation Index (NDVI) | Habitat Suitability for Hosts [31] | 30 arcseconds | 2009–2014 | MODIS |
| Elevation | Diffusion of Hosts [30,35] | 30 arcseconds | 2000 | Shuttle Radar Topography Mission (SRTM) |
| Population (Human) | Indication of Poultry Trade [33,40] | 3 arcseconds | 2010 | WorldPop |
| Live Bird Markets | Viral Mixing and Diffusion [41] | 3 arcseconds | 2009–2015 | Systematic Surveillance |
| Poultry Density | Viral Transmission and Diffusion [40] | 3 arcminutes | 2005 | Food and Agriculture Organization (FAO) |
| Fresh Water and Roads | Viral & Host Diffusion [33,35] | 1:1,000,000, 1:50,000 | 1992, 2015 | Defense Mapping Agency’s (DMA) Digital Chart of the World (DCW), OpenStreetMap (OSM) |
| Wealth Index | Biosecurity and Disease Mitigation [42] | Demographic and Health Survey (DHS) Clusters | 2008 | DHS |
| Water Scarcity | Habitat Suitability and Host Diffusion [42] | DHS Clusters | 2008 | DHS |
| Poultry Husbandry | Biosecurity [32] | DHS Clusters | 2008 | DHS |
| H5N1 | H9N2 | Co-Infection | Total | ||
|---|---|---|---|---|---|
| Source | Systematic Surveillance | 145 | 39 | 57 | 241 |
| World Organization for Animal Health (OIE) | 287 | 0 | 0 | 287 | |
| EMPRES-i | 1997 | 67 | 0 | 2064 | |
| Habitat | Commercial | 133 (65.84) | 28 (13.86) | 41 (20.30) | 202 |
| Backyard | 261 (94.91) | 6 (2.18) | 8 (2.91) | 275 | |
| Live-bird Market | 32 (71.11) | 5 (11.11) | 8 (17.78) | 45 | |
| Other/Unspecified | 2003 (96.76) | 67 (3.34) | 0 | 2070 | |
| Host | Chicken | 822 (84.39) | 100 (10.27) | 52 (5.34) | 974 |
| Duck | 390 (98.73) | 1 (0.25) | 4 (1.01) | 395 | |
| Mixed | 724 (99.86) | 0 | 1 (0.14) | 725 | |
| Other/Unspecified | 493 (99.00) | 5 (1.00) | 0 | 498 | |
| Total | - | 2429 (93.71) | 106 (4.09) | 57 (2.20) | 2592 |
| Dataset (Subset) | AUC | Bias | Standard Error |
|---|---|---|---|
| H5N1 | 0.9276 | −0.0005 | 0.0035 |
| H5N1 (Quadrant Test) | 0.7413 | 0.0003 | 0.0087 |
| H9N2 | 0.9379 | 0.0003 | 0.0260 |
| H9N2 (Quadrant Test) | 0.6902 | −0.0049 | 0.0485 |
| Co-Infections | 0.9918 | 0.00008 | 0.0078 |
| H5N1 × H9N2 1 | 0.9948 | −0.00002 | 0.0015 |
| Mean H5N1 & H9N2 1 | 0.9939 | 0.0004 | 0.0020 |
| H5N1 1 | 0.9784 | −0.0006 | 0.0044 |
| H9N2 1 | 0.9923 | 0.0001 | 0.0022 |
| Comparison (X vs. Y) | I Metric | X vs. Random Y | Y vs. Random X |
|---|---|---|---|
| H5N1 vs. H9N2 | 0.892 1 | (0.673, 0.763) | (0.754, 0.762) |
| Co-Infection vs. H5N1 | 0.478 1 | (0.261, 0.272) | (0.677, 0.780) |
| Co-Infection vs. H9N2 | 0.514 1 | (0.185, 0.272) | (0.682, 0.789) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, S.G.; Carrel, M.; Malanson, G.P.; Ali, M.A.; Kayali, G. Predicting Avian Influenza Co-Infection with H5N1 and H9N2 in Northern Egypt. Int. J. Environ. Res. Public Health 2016, 13, 886. https://doi.org/10.3390/ijerph13090886
Young SG, Carrel M, Malanson GP, Ali MA, Kayali G. Predicting Avian Influenza Co-Infection with H5N1 and H9N2 in Northern Egypt. International Journal of Environmental Research and Public Health. 2016; 13(9):886. https://doi.org/10.3390/ijerph13090886
Chicago/Turabian StyleYoung, Sean G., Margaret Carrel, George P. Malanson, Mohamed A. Ali, and Ghazi Kayali. 2016. "Predicting Avian Influenza Co-Infection with H5N1 and H9N2 in Northern Egypt" International Journal of Environmental Research and Public Health 13, no. 9: 886. https://doi.org/10.3390/ijerph13090886
APA StyleYoung, S. G., Carrel, M., Malanson, G. P., Ali, M. A., & Kayali, G. (2016). Predicting Avian Influenza Co-Infection with H5N1 and H9N2 in Northern Egypt. International Journal of Environmental Research and Public Health, 13(9), 886. https://doi.org/10.3390/ijerph13090886