
Pathological Impairment, Cell Cycle Arrest and Apoptosis of Thymus and Bursa of Fabricius Induced by Aflatoxin-Contaminated Corn in Broilers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diet, and Study Design
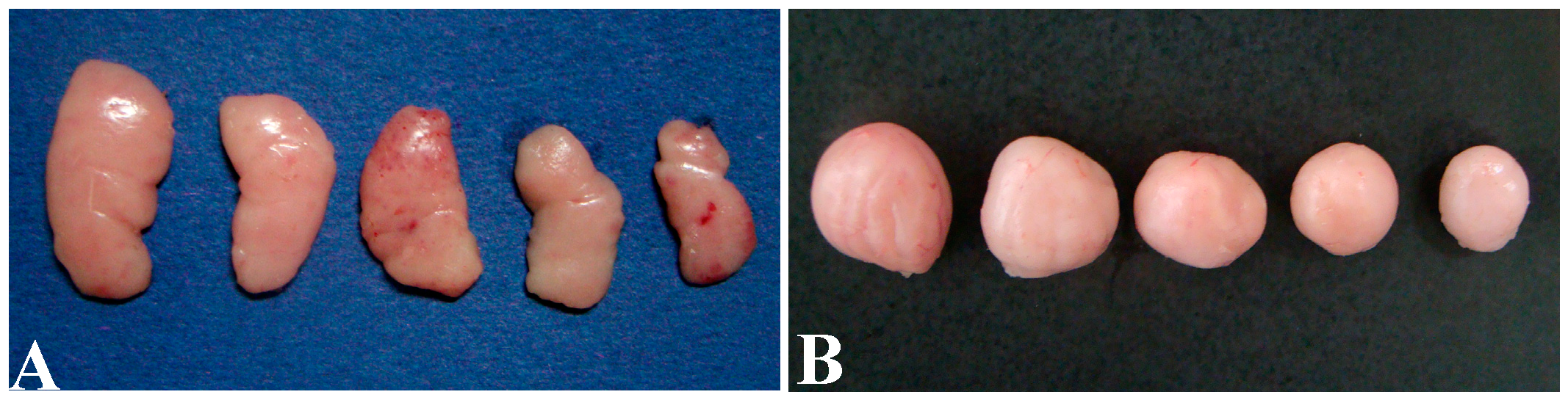
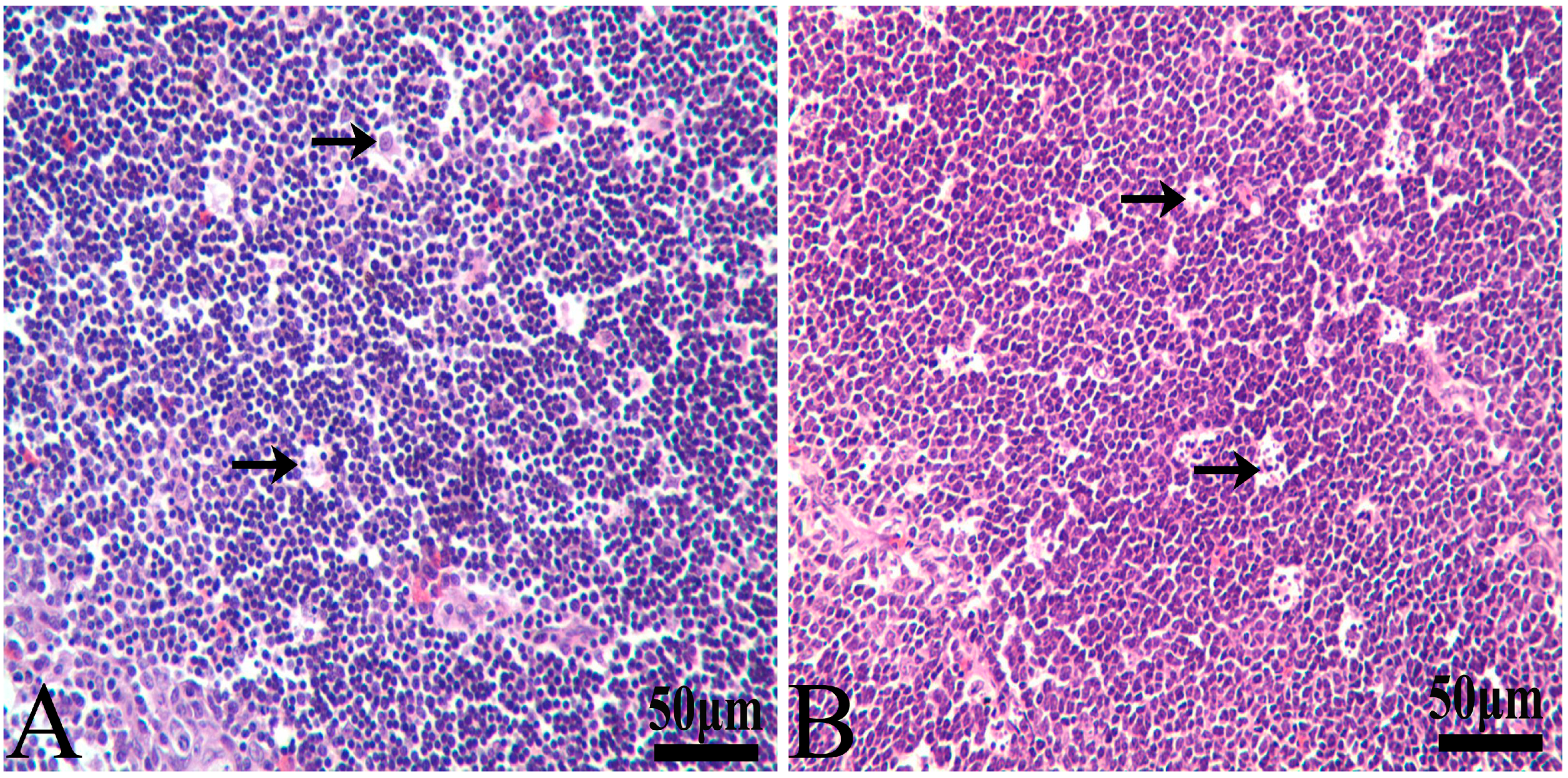
2.2. Pathological Observation
2.3. Cell Cycle Phase Detection by Flow Cytometry
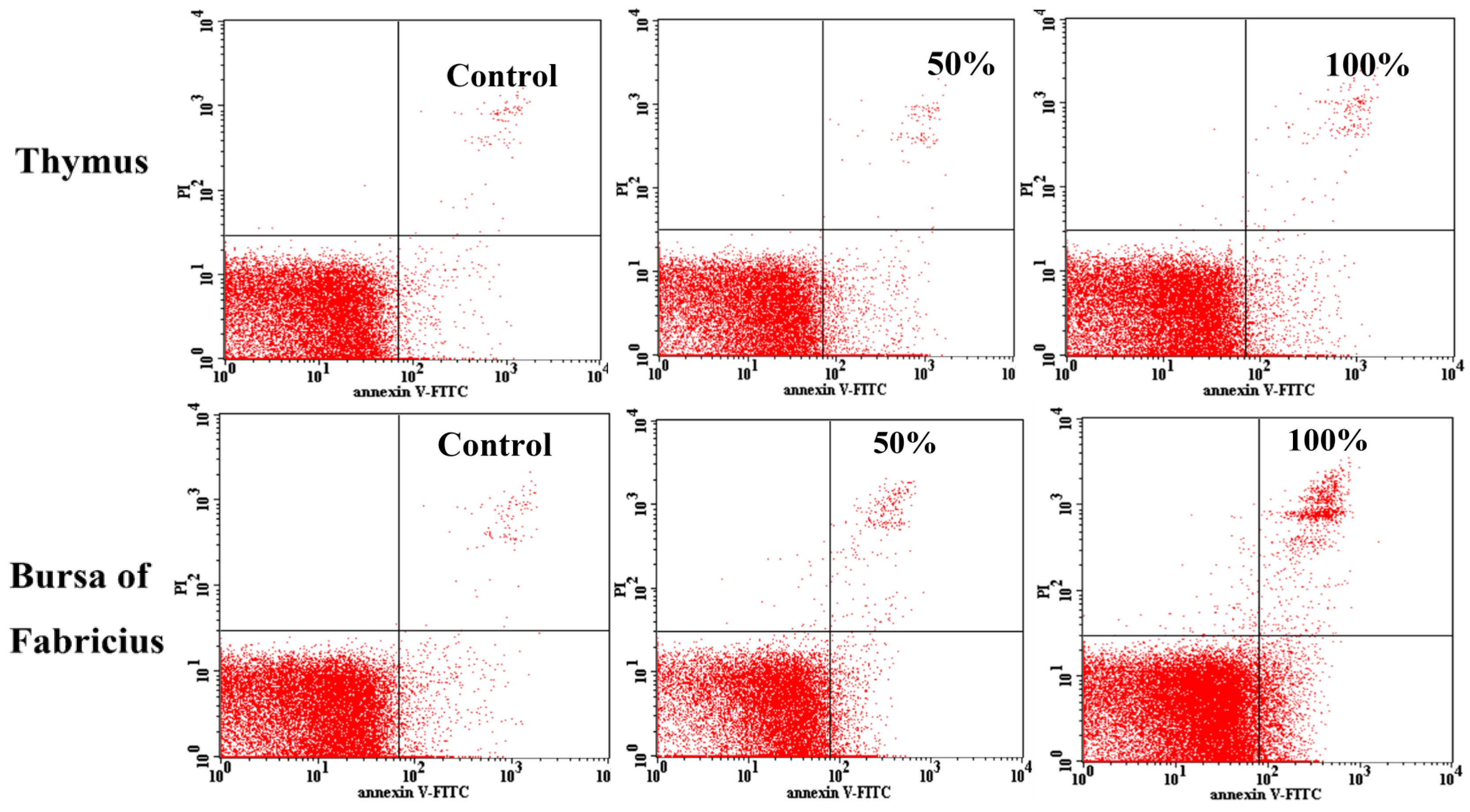
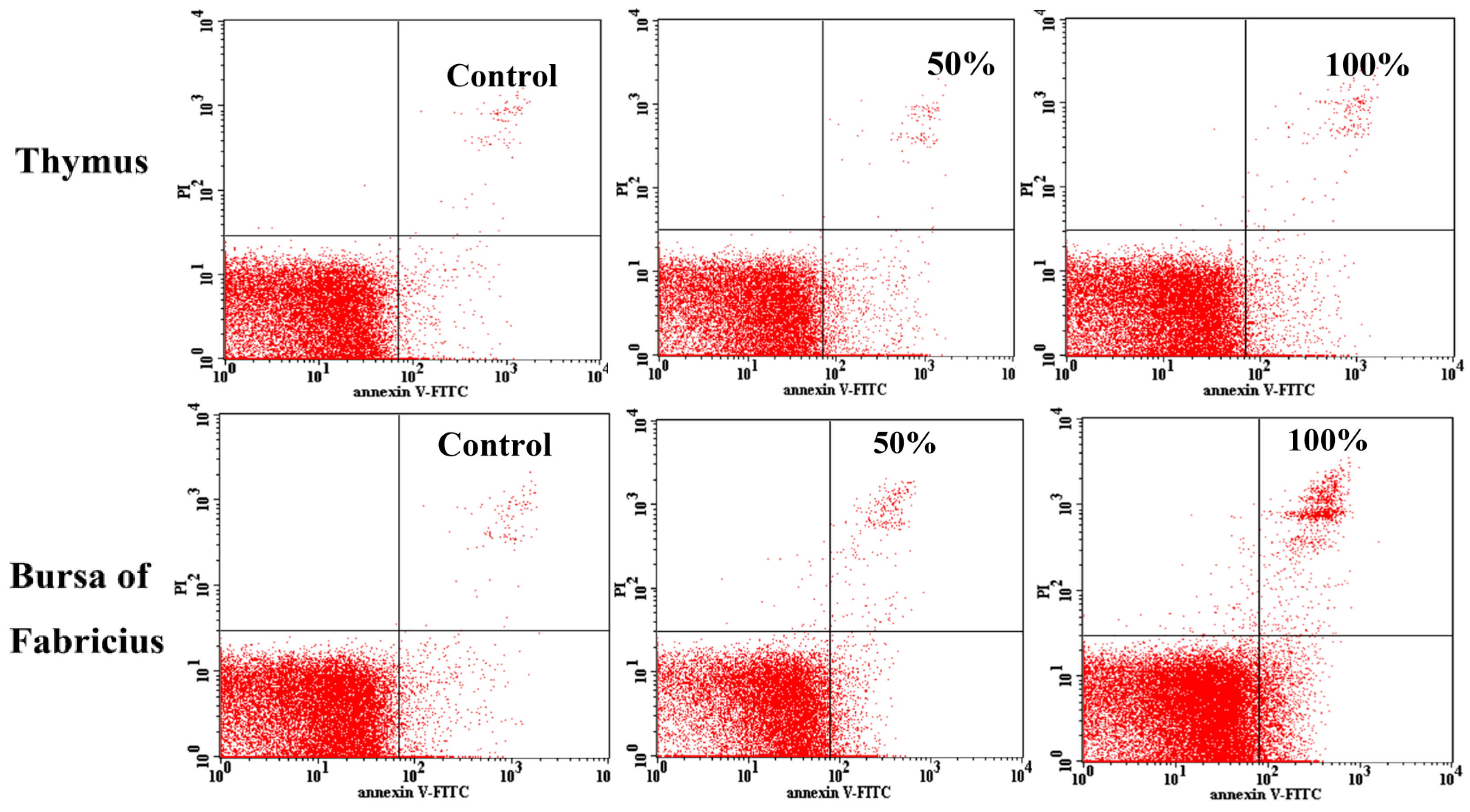
2.4. Annexin V Apoptosis Detection by Flow Cytometry
3. Results
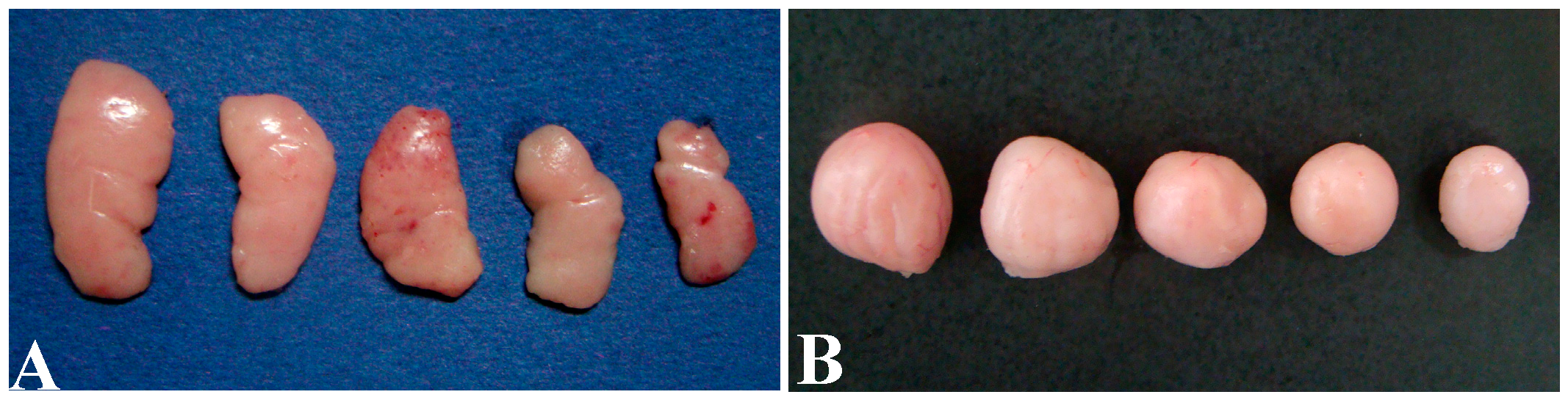
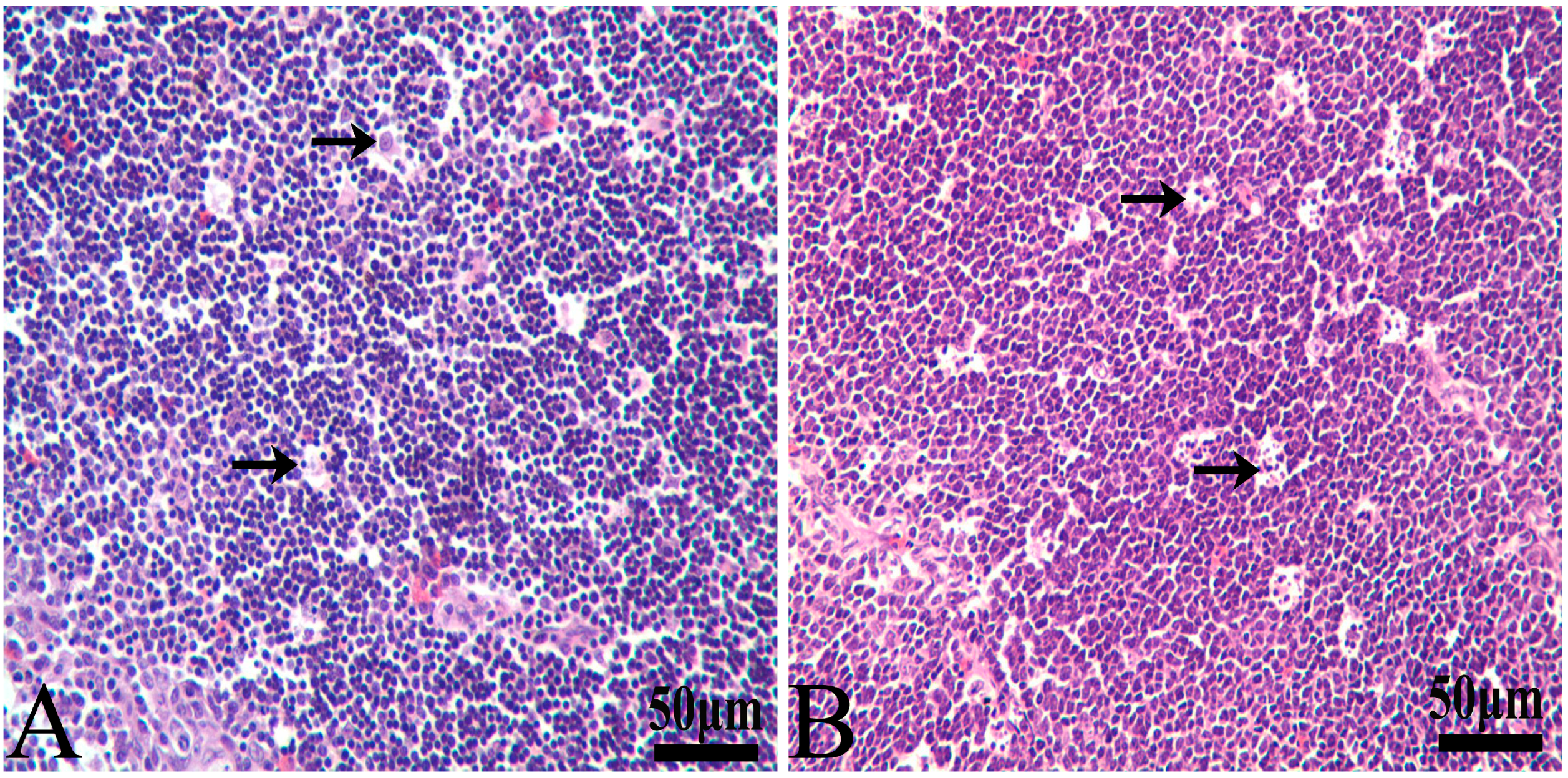
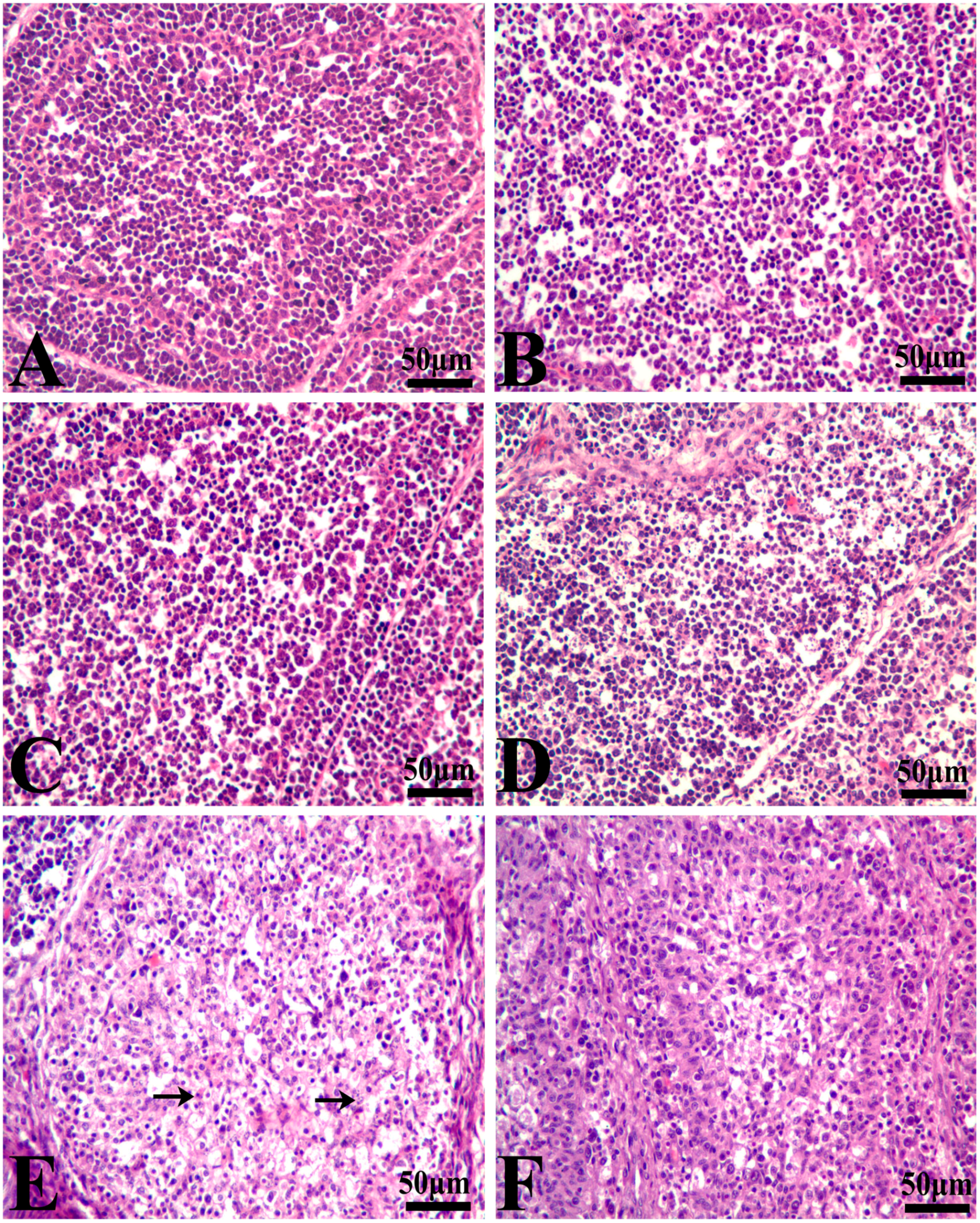
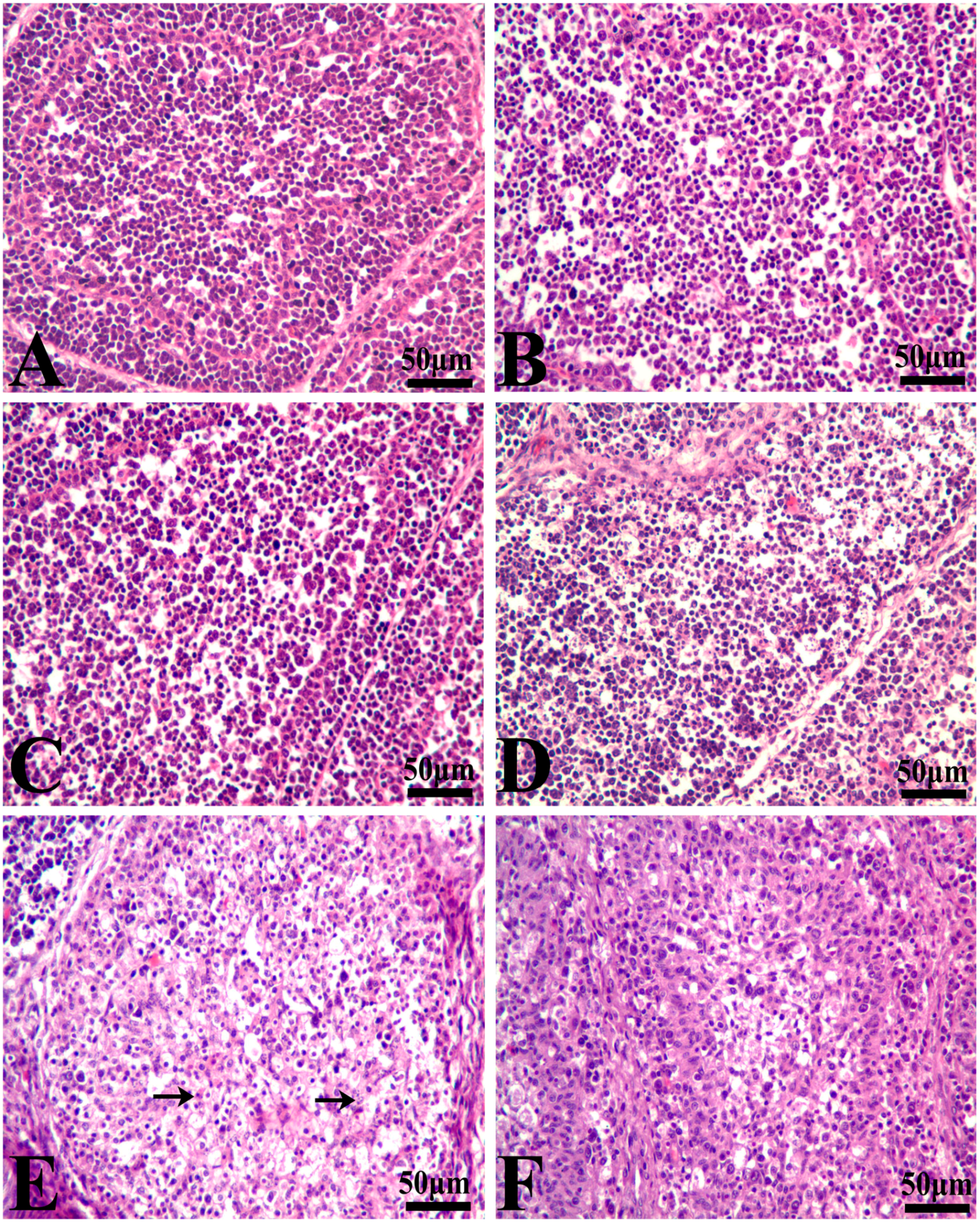
3.1. Histopathological Lesions
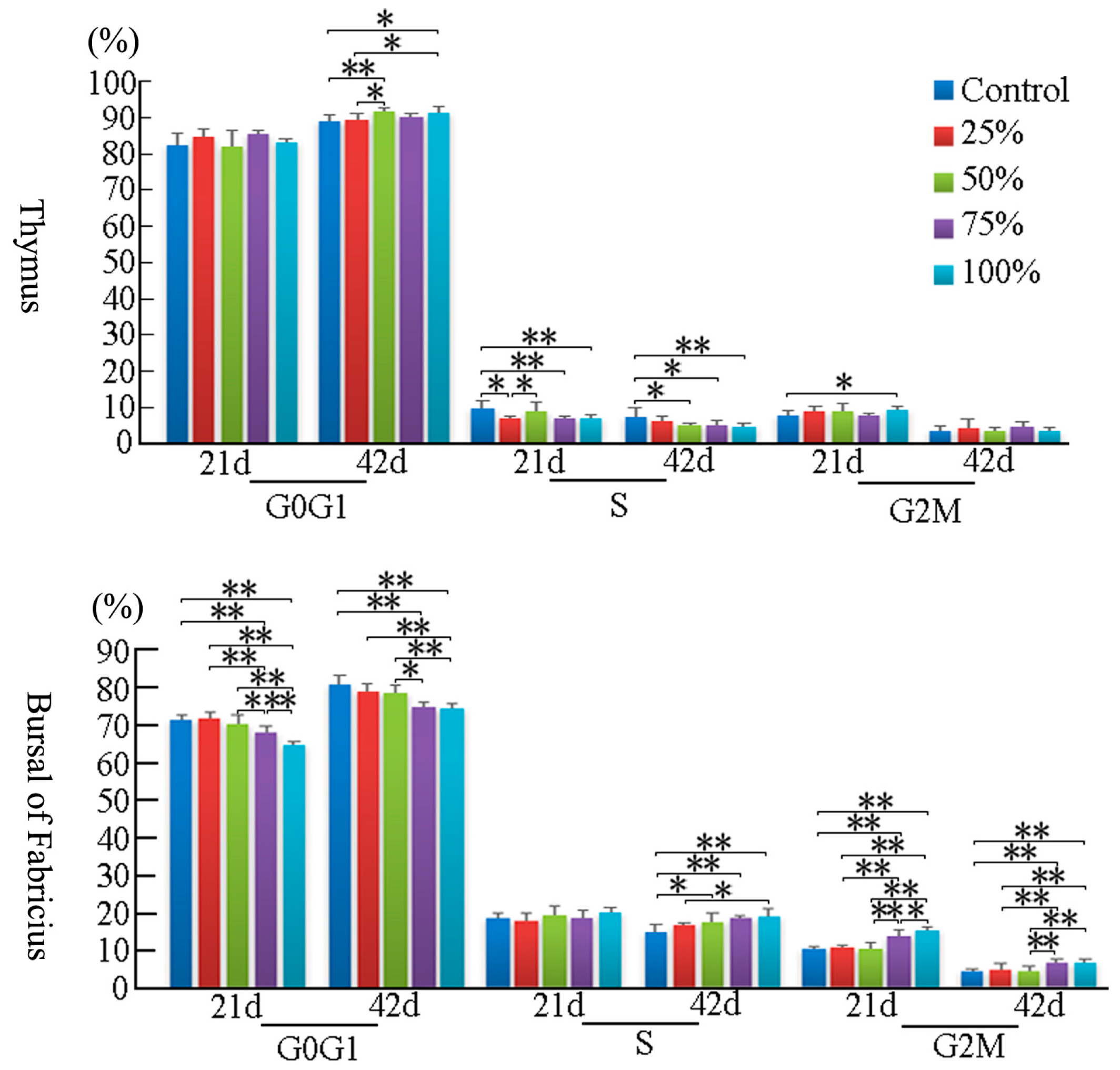
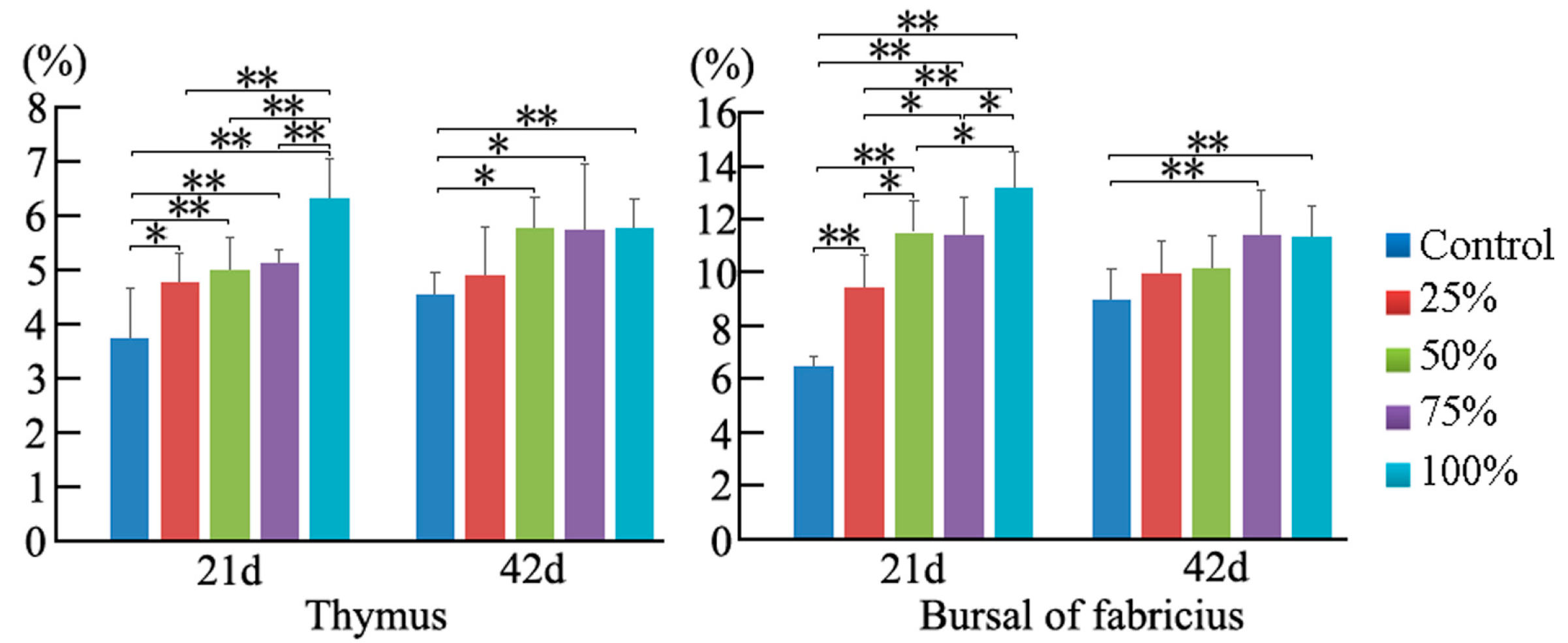
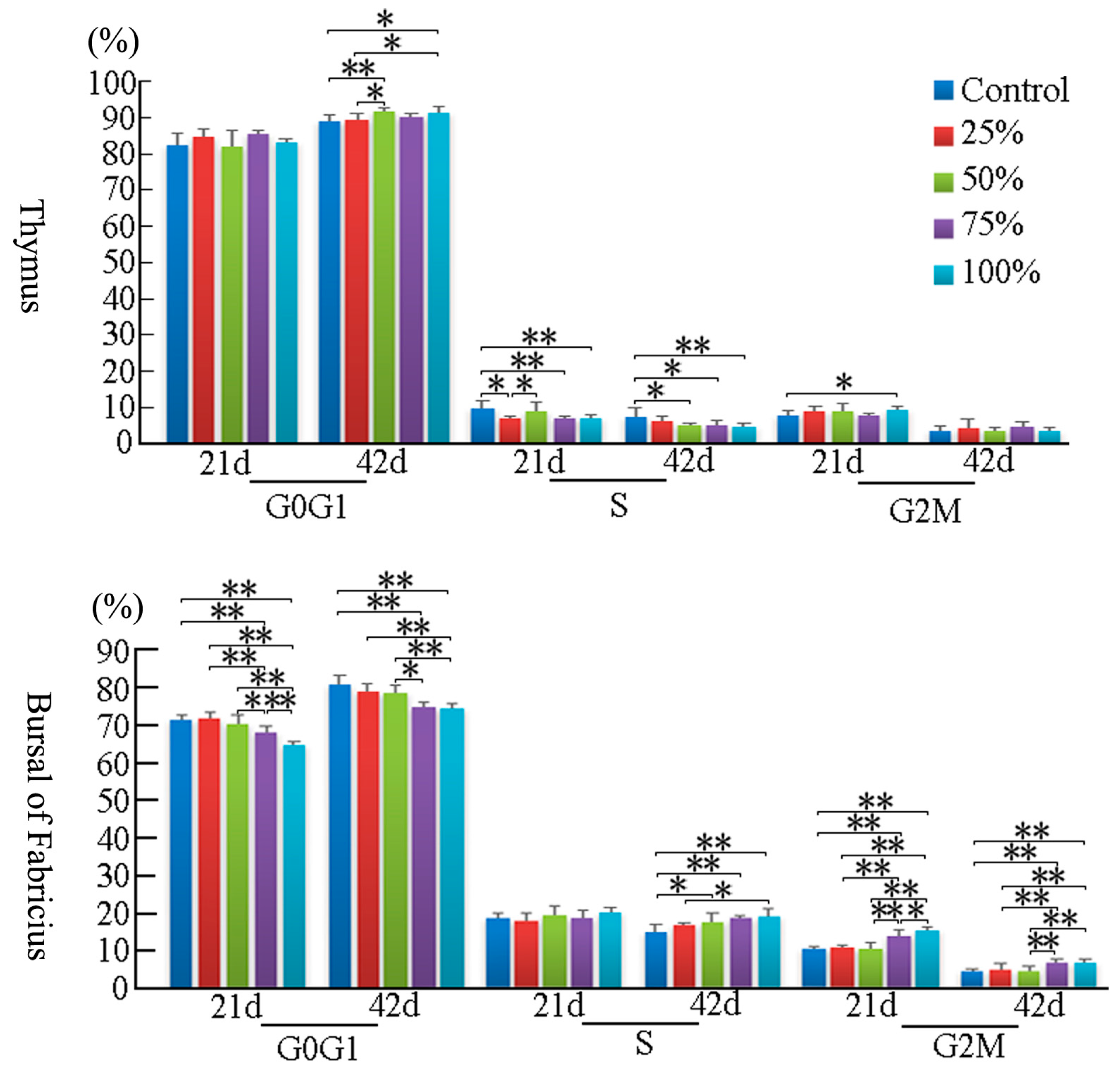
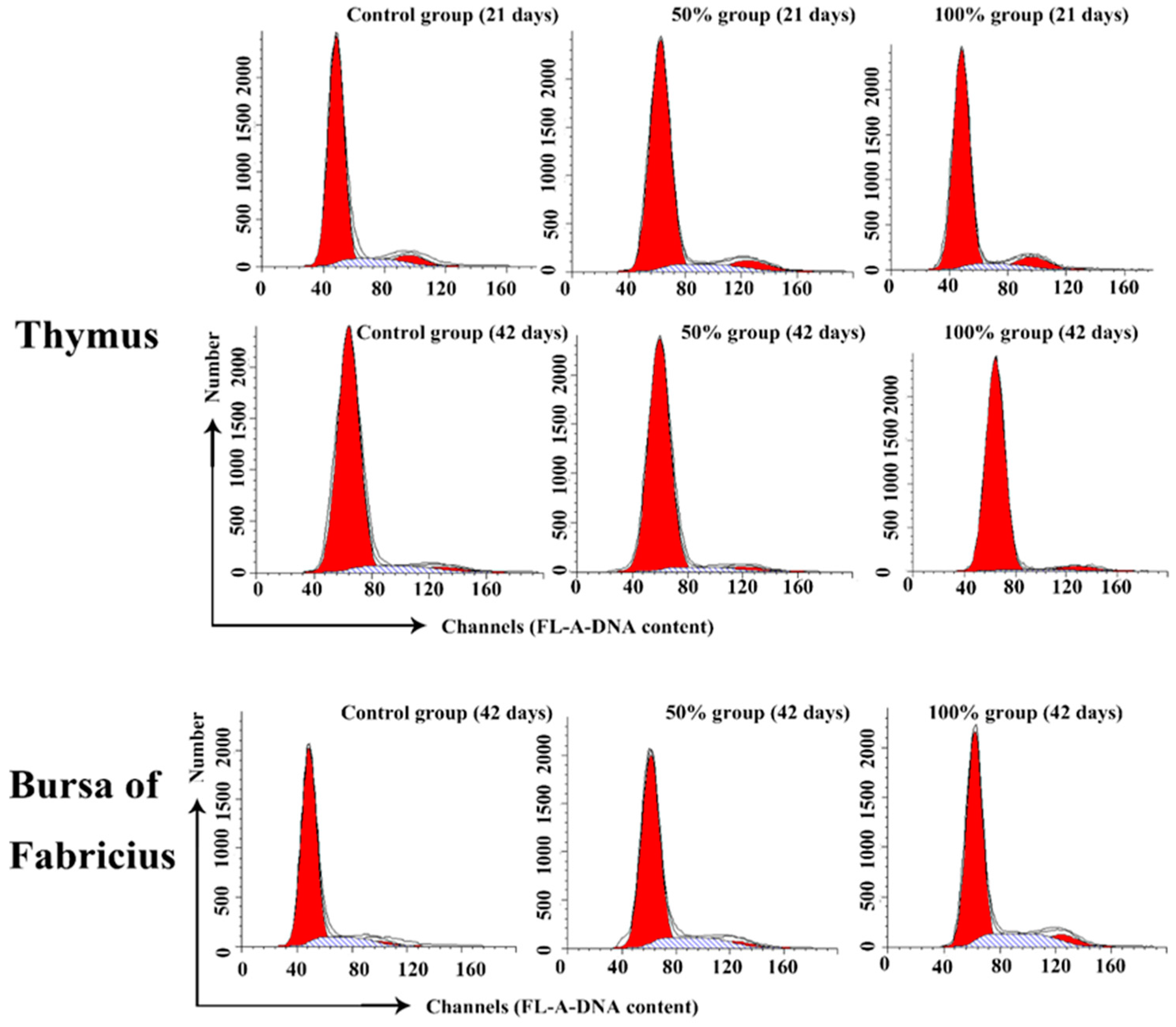
3.2. Cell Cycle Phase Distribution of Immune Organs
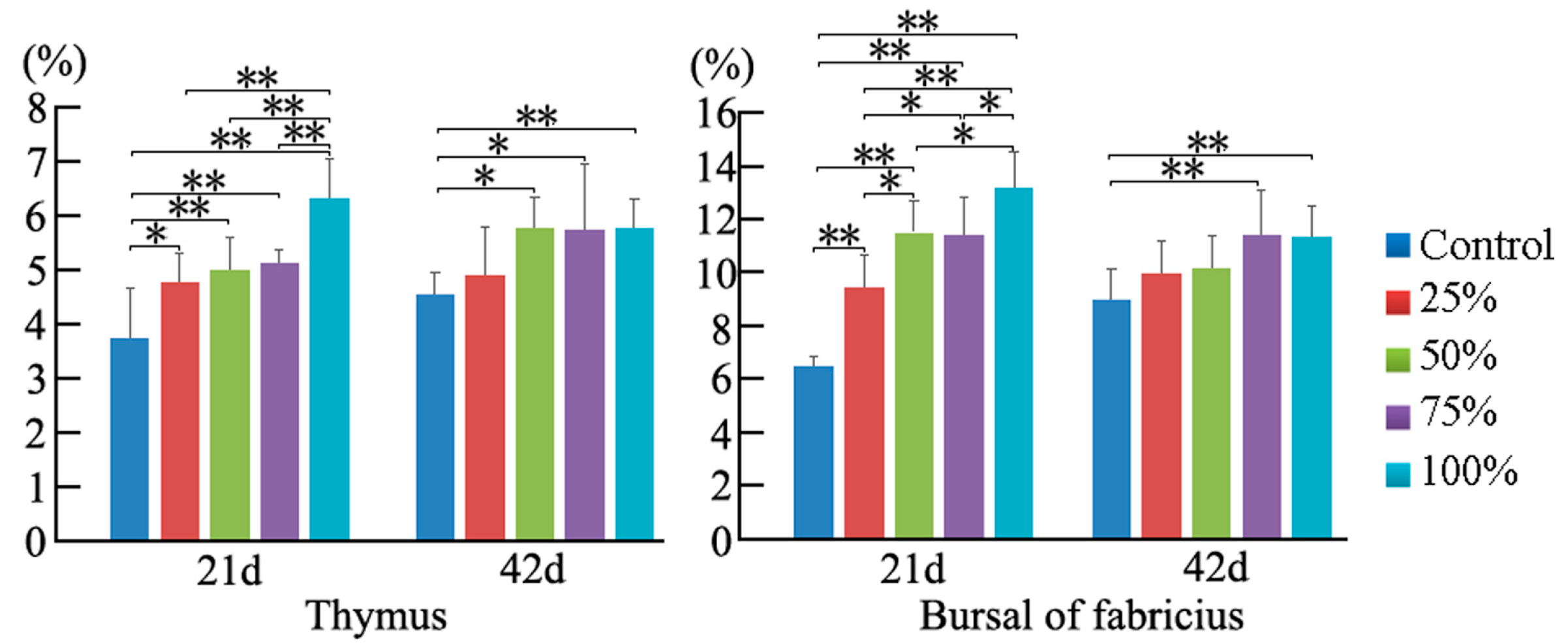
3.3. Annexin V-FITC Staining Assay by Flow Cytometry
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16 (Suppl. 1), 95–101. [Google Scholar] [PubMed]
- Mannon, J.; Johnson, E. Fungi down on the farm. New Sci. 1985, 105, 12–16. [Google Scholar]
- Mclean, M.; Dutton, M.F. Cellular interactions and metabolism of aflatoxin: An update. Pharmacol. Ther. 1995, 65, 163–192. [Google Scholar] [CrossRef]
- Eaton, D.L.; Groopman, J.D. The Toxicology of Aflatoxins: Human Health, Veterinary, and Agricultural Significance. Anim. Feed Sci. Technol. 1997, 68, 368–369. [Google Scholar]
- Benford, D.; Dinovi, M.; Setzer, R.W. Application of the margin-of-exposure (MoE) approach to substances in food that are genotoxic and carcinogenic e.g., Benzo[a]pyrene and polycyclic aromatic hydrocarbons. Food Chem. Toxicol. 2010, 48 (Suppl. 1), S42–S48. [Google Scholar] [CrossRef] [PubMed]
- Wogan, G.; Newberne, P. Dose-response characteristics of aflatoxin B1 carcinogenesis in the rat. Cancer Res. 1967, 27, 2370–2376. [Google Scholar] [PubMed]
- Eaton, D.L.; Gallagher, E.P. Mechanisms of aflatoxin carcinogenesis. Annu. Rev. Pharmacol. 1994, 34, 135–172. [Google Scholar] [CrossRef] [PubMed]
- Newberne, P.M.; Butler, W.H. Acute and chronic effects of aflatoxin on the liver of domestic and laboratory animals: A review. Cancer Res. 1969, 29, 236–250. [Google Scholar] [PubMed]
- Yu, Z.Q.; Wang, F.Y.; Liang, N.; Wang, C.H.; Peng, X.; Fang, J.; Cui, H.M.; Jameel, M.; Lai, W.M. Effect of Selenium Supplementation on Apoptosis and Cell Cycle Blockage of Renal Cells in Broilers Fed a Diet Containing Aflatoxin B1. Biol. Trace Elem. Res. 2015, 168, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Mughal, M.J.; Peng, X.; Kamboh, A.A.; Zhou, Y.; Fang, J. Aflatoxin B1 induced systemic toxicity in poultry and rescue effects of Selenium and Zinc. Biol. Trace Elem. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhang, S.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J. Protective roles of sodium selenite against aflatoxin B1-induced apoptosis of jejunum in broilers. Int. J. Environ. Res. Public Health 2014, 11, 13130–13143. [Google Scholar] [CrossRef] [PubMed]
- Raisuddin, S.; Zaidi, S.I.; Singh, K.P.; Ray, P.K. Effect of subchronic aflatoxin exposure on growth and progression of Ehrlich’s ascites tumor in mice. Drug Chem. Toxicol. 1991, 14, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Bodine, A.B.; Fisher, S.F.; Gangjee, S.; Fisher, S.F.; Gangjee, S. Effect of aflatoxin B1 and major metabolites on phytohemeagglutinin-stimulated lymphoblastogenesis of bovine lymphocytes. J. Dairy Sci. 1984, 67, 110–114. [Google Scholar] [CrossRef]
- Solcan, C.; Solcan, G.; Oprisan, B.; Spataru, C.; Spataru, M.; Floristean, V. Immunotoxic action of aflatoxin B1 against lymphoid organs is coupled with the high expression of BCL-2 by reticuloepithelial cells in broiler chickens. J. Anim. Vet. Adv. 2014, 13, 537–544. [Google Scholar]
- Peng, X.; Yu, Z.; Liang, N.; Chi, X.; Li, X.; Jiang, M.; Fang, J.; Cui, H.; Lai, W.; Zhou, Y.; et al. The mitochondrial and death receptor pathways involved in the thymocytes apoptosis induced by aflatoxin B1. Oncotarget 2016, 7, 12222–12234. [Google Scholar] [PubMed]
- Yuan, S.; Wu, B.; Yu, Z.; Fang, J.; Liang, N.; Zhou, M.; Huang, C.; Peng, X. The mitochondrial and endoplasmic reticulum pathways involved in the apoptosis of bursa of Fabricius cells in broilers exposed to dietary aflatoxin B1. Oncotarget 2016, 7, 65295–65306. [Google Scholar] [PubMed]
- Liu, B.H.; Yu, F.Y.; Chan, M.H.; Yang, Y.L. The effects of mycotoxins, fumonisin B1 and aflatoxin B1, on primary swine alveolar macrophages. Toxicol. Appl. Pharmcol. 2002, 180, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Corradi, A.; Santarelli, L.; Tibaldi, A.; DeAngelis, E.; Borghetti, P.; Bonomi, A.; Fabris, N.; Cabassi, E. Zinc, thymic endocrine activity and mitogen responsiveness (PHA) in piglets exposed to maternal aflatoxicosis B1 and G1. Vet. Immunol. Immunopathol. 1998, 62, 245–260. [Google Scholar] [CrossRef]
- Hinton, D.M.; Myers, M.J.; Raybourne, R.A.; Francke-Carroll, S.; Sotomayor, R.E.; Shaddock, J.; Warbritton, A.; Chou, M.W. Immunotoxicity of aflatoxin B1 in rats: Effects on lymphocytes and the inflammatory response in a chronic intermittent dosing study. Toxicol. Sci. 2003, 73, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Silvotti, L.; Petterino, C.; Bonomi, A.; Cabassi, E. Immunotoxicological effects on piglets of feeding sows diets containing aflatoxins. Vet. Rec. 1997, 141, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmcol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Hashiba, Y.; Ohtsuka, H.; Kohiruimaki, M.; Masui, M.; Kawamura, S.; Endo, H.; Ogata, Y. Effects of mycotoxins on mitogen-stimulated proliferation of bovine peripheral blood mononuclear cells. J. Vet. Med. Sci. 2008, 70, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Methenitou, G.; Maravelias, C.; Athanaselis, S.; Dona, A.; Koutselinis, A. Immunomodulative effects of aflatoxins and selenium on human natural killer cells. Vet. Hum. Toxicol. 2001, 43, 232–234. [Google Scholar] [PubMed]
- Chen, K.J.; Shu, G.; Peng, X.; Fang, J.; Cui, H.M.; Chen, J.; Wang, F.Y.; Chen, Z.L.; Zuo, Z.C.; Deng, J.L.; et al. Protective role of sodium selenite on histopathological lesions, decreased T-cell subsets and increased apoptosis of thymus in broilers intoxicated with aflatoxin B1. Food Chem. Toxicol. 2013, 59, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.J.; Fang, J.; Peng, X.; Cui, H.M.; Chen, J.; Wang, F.Y.; Chen, Z.L.; Zuo, Z.C.; Deng, J.L.; Lai, W.M.; et al. Effect of selenium supplementation on aflatoxin B1-induced histopathological lesions and apoptosis in bursa of Fabricius in broilers. Food Chem. Toxicol. 2014, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, K.; Yuan, S.; Peng, X.; Fang, J.; Wang, F.; Cui, H.; Chen, Z.; Yuan, J.; Geng, Y. Effects of aflatoxin B1 on oxidative stress markers and apoptosis of spleens in broilers. Toxicol. Ind. Health 2016, 32, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Bai, S.P.; Ding, X.M.; Zeng, Q.F.; Zhang, K.Y.; Fang, J. Pathological changes in the immune organs of broiler chickens fed on corn naturally contaminated with aflatoxins B1 and B2. Avian Pathol. 2015, 44, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bai, F.; Zhang, K.; Bai, S.; Peng, X.; Ding, X.; Li, Y.; Zhang, J.; Zhao, L. Effects of feeding corn naturally contaminated with aflatoxin B1 and B2 on hepatic functions of broilers. Poult. Sci. 2012, 91, 2792–2801. [Google Scholar] [CrossRef] [PubMed]
- Administration of Quality and Technology Supervision of Sichuan Province (AQTSSP). Official Standard of Administration of Quality and Technology Supervision of Sichuan Province of PRC, Determination of Aflatoxin B1, B2, G1, G2, T-2 Toxin, Deoxynicalenol, Zearalenone, Ochratoxin, Fumonisin B1 with HPLC (DB 51/T 1077-2010, DB 51/T 1078-2010, DB 51/T 1079-2010, DB 51/T 1080-2010, DB 51/T 1081-2010, DB 51/T 1082-2010); AQTSSP: Chengdu, China, 2010.
- Darzynkiewicz, Z.; Zhao, H. Cell Cycle Analysis by Flow Cytometry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 1–8. [Google Scholar]
- Ricordy, R.; Gensabella, G.; Cacci, E.; Augusti-Tocco, G. Impairment of cell cycle progression by aflatoxin B1 in human cell lines. Mutagenesis 2002, 17, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Ricordy, R.; Cacci, E.; Augusti-Tocco, G.; Preedy, V.R.; Watson, R.R. Aflatoxin B1 and cell cycle perturbation. In Reviews in Food and Nutrition Toxicity, 4th ed.; Preedy, V.R., Watson, R., Eds.; CRC Press: London, UK, 2005; pp. 213–231. [Google Scholar]
- Scott, T.R.; Rowland, S.M.; Rodgers, R.S.; Bodine, A.B. Genetic selection for aflatoxin B1 resistance influences chicken T-cell and thymocyte proliferation. Dev. Comp. Immunol. 1991, 15, 383–391. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, K.; Bai, S.; Ding, X.; Zeng, Q.; Yang, J.; Fang, J.; Chen, K. Histological lesion, cell cycle arrest, apoptosis and T cell subset changes of spleen in chicken fed aflatoxin-contaminated corn. Int. J. Environ. Res. Public Health 2014, 11, 8567–8580. [Google Scholar] [CrossRef] [PubMed]
- Elmre, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Kanduc, D.; Mittelman, A.; Serpico, R.; Sinigaglia, E.; Sinha, A.A.; Natale, C.; Santacroce, R.; Di Corcia, M.G.; Lucchese, A.; Dini, L.; et al. Cell death: Apoptosis versus necrosis (review). Int. J. Oncol. 2002, 21, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutellingsperger, C. A novel assay for apoptosis flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Peng, X.; Chen, K.; Chen, J.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W. Aflatoxin B1 affects apoptosis and expression of Bax, Bcl-2, and Caspase-3 in thymus and bursa of fabricius in broiler chickens. Environ. Toxicol. 2016, 31, 1113–1120. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet 1 | Control | 25% | 50% | 75% | 100% | Control Corn | Contaminated Corn |
|---|---|---|---|---|---|---|---|
| 1–21 days | |||||||
| AFB1 | ND 2 | 16.3 | 36.9 | 45.6 | 82.4 | ND | 149.6 |
| AFB2 | ND | 3.15 | 6.38 | 7.86 | 14.2 | ND | 24.2 |
| 22–42 days | |||||||
| AFB1 | ND | 34.3 | 69.3 | 95.2 | 134 | ND | 229 |
| AFB2 | ND | 6.17 | 12.1 | 17 | 23.6 | ND | 37.8 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Bai, S.; Ding, X.; Zhang, K. Pathological Impairment, Cell Cycle Arrest and Apoptosis of Thymus and Bursa of Fabricius Induced by Aflatoxin-Contaminated Corn in Broilers. Int. J. Environ. Res. Public Health 2017, 14, 77. https://doi.org/10.3390/ijerph14010077
Peng X, Bai S, Ding X, Zhang K. Pathological Impairment, Cell Cycle Arrest and Apoptosis of Thymus and Bursa of Fabricius Induced by Aflatoxin-Contaminated Corn in Broilers. International Journal of Environmental Research and Public Health. 2017; 14(1):77. https://doi.org/10.3390/ijerph14010077
Chicago/Turabian StylePeng, Xi, Shiping Bai, Xuemei Ding, and Keying Zhang. 2017. "Pathological Impairment, Cell Cycle Arrest and Apoptosis of Thymus and Bursa of Fabricius Induced by Aflatoxin-Contaminated Corn in Broilers" International Journal of Environmental Research and Public Health 14, no. 1: 77. https://doi.org/10.3390/ijerph14010077