1. Introduction
Ixodes ricinus (Linnaeus 1758) is the most widespread tick species in Europe. It widely occurs in sylvatic as well as in urban habitats with ample vegetation such as parks, recreational areas, gardens and urban forests. Emergence of ticks in urban areas is associated with a higher risk of tick-borne diseases for humans and companion animals [
1]. Occurrence of ticks in urban parks is significantly influenced by vegetation that maintains adequate humidity for ticks as well as the abundance and species composition of hosts [
2,
3,
4].
I. ricinus ticks transmit several bacterial agents. The most common ones are spirochetes from the
Borrelia burgdorferi sensu lato complex. This bacterial complex consists of 21 known genospecies [
5,
6,
7,
8,
9,
10,
11,
12]. Five out of eight genospecies that are present in Europe are potentially infectious for humans and can cause Lyme disease.
Reported mean prevalence of
B. burgdorferi s.l., detected by various detection methods including polymerase chain reaction (PCR), cultivation in BSK medium, dark-field microscopy and immunofluorescence assays, in ticks in Europe is 12.3–13.7% [
13,
14]. In Slovakia, the prevalence ranges from 4.5%, detected in questing
I. ricinus in a suburban forest in northern Slovakia [
15], to 46.1% for a mountain region of central Slovakia [
16]. Prevalences of this bacterial group in urban and suburban forests across various cities in Slovakia were recorded to reach 20.5% [
15,
17,
18]. Genetic variability within and between
Borrelia genospecies is linked to different clinical symptoms as well as different associations to reservoir hosts. Rodents, especially from genera
Myodes and
Apodemus act as the reservoirs of
B. afzelii,
B. burgdorferi s.s.,
B. bavariensis, and
B. spielmanii [
19,
20,
21,
22,
23]. The circulation of
B. afzelii in natural foci of urban parks and forests is also associated with squirrels and hedgehogs [
20,
24], while
B. spielmanii is associated with hedgehogs [
24] and dormice [
25]. Birds are the reservoirs of
B. garinii,
B. valaisiana [
26] and in a low degree also for
B. lusitaniae [
27] for which the main reservoir hosts are the lizards [
28].
Anaplasma phagocytophilum is another important bacterial agent transmitted by
I. ricinus in Europe. It causes granulocytic anaplasmosis of humans, horses and dogs, as well as tick-borne fever of domestic ruminants. In Slovakia, the prevalence of
A. phagocytophilum in questing
I. ricinus ticks varies between 1.1% and 8% [
15,
29,
30,
31,
32]. In Europe,
Anaplasma circulates in two different cycles. The most important reservoir hosts in Europe are wild cervids, sheep, cattle and goats [
29,
33,
34,
35,
36]. Rodents are reservoir hosts of
A. phagocytophilum genotypes that are transmitted by
I. trianguliceps, but not by
I. ricinus ticks [
36,
37,
38,
39]. In areas with abundant populations of wild cervids, the prevalence of
Anaplasma is usually higher [
34,
39].
Ixodes ricinus ticks are also known to carry some species of
Rickettsia, obligate, aerobic, gram-negative, intracellular bacterial parasites of eukaryotes with a worldwide occurrence, which can cause rickettsial diseases.
Rickettsia helvetica and
Rickettsia monacensis transmitted by
I. ricinus are rickettsial species associated with human cases of rickettsiosis [
40,
41]. The prevalences vary from 0.5% in an island in the Baltic Sea to 66% in the Netherlands for
R. helvetica, and from 0.5% in Germany to 34.6% in Turkey for
R. monacensis [
42]. In Slovakia, rickettsial prevalence in
I. ricinus ticks were recorded up to 20%, with
R. helvetica being a dominant species [
43,
44]. The role of vertebrates in their circulation is still not clear. Rickettsiae have been identified in a few randomly selected samples of the spleens of small rodents and ungulates [
45,
46].
All of
I. ricinus stages are obligate parasites. Immature stages of ticks feed most frequently on small mammals and birds. Howeverd, the most important host of adult ticks are large mammals, such as deer and wild boar. Immature ticks can compensate for the absence of birds and small mammals by feeding on larger animals, but their abundance may decrease [
47,
48].
The population structure of tick hosts in different sites and habitats can affect the abundance of tick-borne pathogens such as
Borrelia and
Anaplasma due to differences in host competence [
49,
50,
51]. For example, roe deer commonly acts as the main host for adult ticks and in a lower degree for immature ticks [
52]. The increase of roe deer abundance may cause a decrease in
Borrelia prevalence in a given habitat [
39], possibly due to the borreliacidal effects of deer blood-complement [
53]. Variation in the abundance and diversity of tick hosts has been suggested as a crucial determinant of the prevalence and density of tick-borne pathogens [
54]. For example, some might suggest that densities of infected ticks should be lower at sites with higher tick-host biodiversity [
55], but see [
56]. However, the abundance and diversity of tick hosts and, consequently, the abundance and diversity of tick-borne pathogens, may not only vary spatially, but also temporally within sites, e.g., due to fluctuating densities of bird tick hosts. Despite its importance for epidemiology, seasonality and its causes in the prevalence and community structure of vector-borne pathogens within sites have been scarcely studied (e.g., [
57] for prevalence of
B. burgdorferi s.l. and
A. phagocytophilum during May and August).
The aim of this study was to examine the seasonal dynamics in
I. ricinus abundance as well as the prevalence and community structure of
B. burgdorferi s.l.,
A. phagocytophilum and
Rickettsia spp. in an urban forest in Slovakia. A recent metaanalysis has revealed a strong negative correlation between the two most prevalent
Borrelia genospecies in Europe—
B. afzelii and
B. garinii [
14], suggesting that small rodents and birds represent the main alternative hosts for immature
I. ricinus ticks in the region. Given the abundance of temperate-zone birds undergoes abrupt seasonal changes corresponding to their reproductive period (e.g., [
58]), we hypothesized that the seasonal pattern in the prevalences of tick-borne pathogens at the study site would reflect the phenology of bird abundance. Consequently, we predicted that the prevalence of the commonest
Borrelia genospecies associated with bird hosts (
B. garinii) and small rodent hosts (
B. afzelii) should show divergent seasonal patterns, reflecting the breeding phenology of temperate-zone birds.
4. Discussion
Changes in biotic and abiotic conditions can increase or decrease the transmission of zoonotic diseases [
4]. Variation in tick-borne pathogen prevalence is traditionally viewed in the context of spatial variation in climatic and weather conditions [
75] or spatial variation, at different scales, in tick-host community structure [
54]. In this study we have investigated questing activity of ticks and their infection with three groups of tick-borne pathogens—
Borrelia,
Anaplasma and
Rickettsia. For an urban temperate forest, we examined whether the prevalence of different tick-borne pathogens varies temporally within a season. We hypothesized that the occurrence of the commonest
Borrelia genospecies in questing ticks should vary non-randomly in time with respect to the reproductive period of birds. This hypothesis is relevant because our study site is known to have a low abundance of small rodents [
53], thus stressing the role of bird hosts for local immature
I. ricinus ticks. We found that the occurrence of different tick-borne pathogens varied non-randomly with time of season.
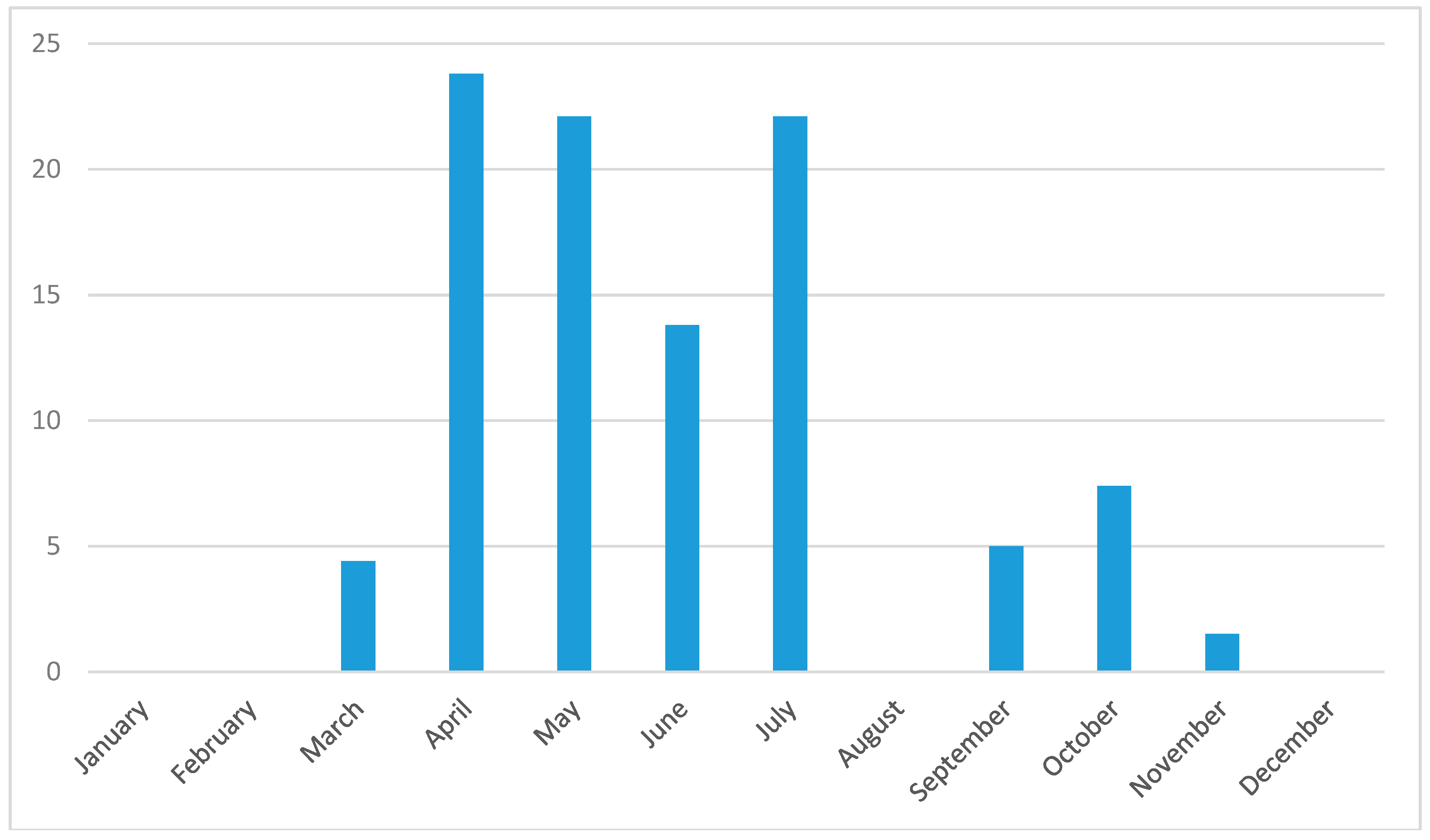
Tick questing activity showed a bimodal seasonal pattern. In both years, the first peak was recorded in April–May and the second one in July. This bimodal seasonal tick activity with one bigger peak in spring and the second one in early fall has been described for various tick habitats in Central Europe [
76,
77,
78,
79]. The overall prevalence of tick-borne pathogens followed the temporal trend in tick abudance. However, by focusing on specific tick-borne pathogens, we detected distinct seasonal trends in the occurrences of the most common pathogens (
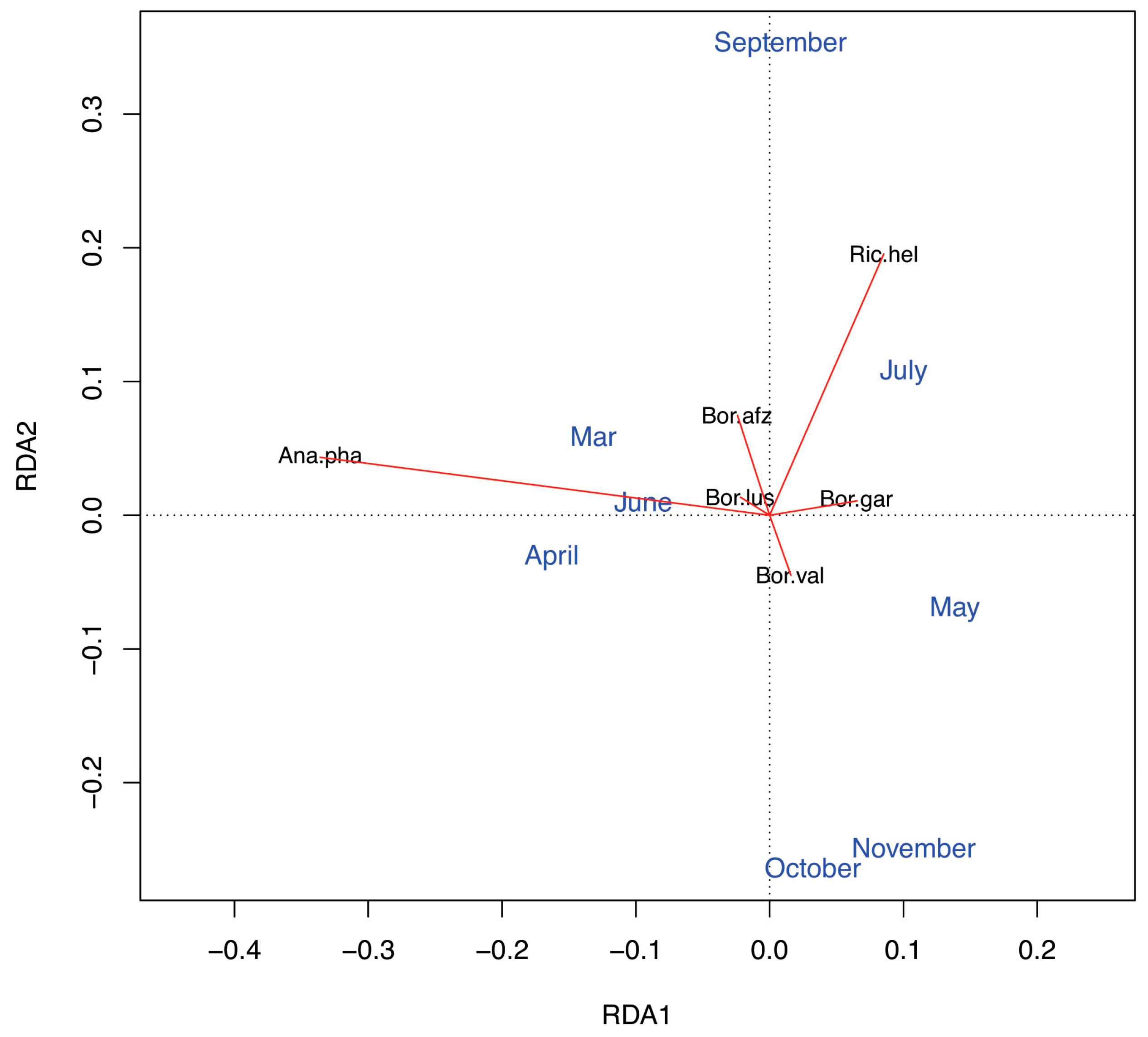
Figure 4). First, the early season (March–May) was characterised by the diverging occurrence of
A. phagocytophilum and
B. garinii. Second, the late season (September–November) was characterised by the divergent occurrence of
B. afzelii and
B. valaisiana. Hence, even though
B. afzelii and
B. garinii typically occurred at different parts of the year, our results do not support a divergent occurrence of
B. afzelii and
B. garinii at our study site. We suggest that this finding is due to a locally lower importance of small rodent hosts during the part of the year with the highest tick abundance (see below). Nevertheless, this study indicates that small rodents may still be important alternative tick hosts during some periods, because we found a divergent occurrence of
B. afzelii and
B. valaisiana during the late season. Importantly, our results lend support to our hypothesis that temporary increases in the abundance of bird hosts during their reproductive period can be reflected by the seasonal trends in the occurrence of the pathogens associated with these hosts, namely,
B. garinii. Intriguingly, the occurrence of
B. garinii in questing ticks was most likely in May, i.e., during the peak reproductive period of birds. Even though we do not possess knowledge of the duration of tick molt at our study site, our results suggest that the cohorts of questing
I. ricinus ticks infected by bird-associated
B. garinii are likely to transmit this pathogen to the same host group. Further research is necessary to address how seasonal fluctuations in abiotic factors, such as temperature and humidity, contribute to the seasonal trends in tick-borne pathogen occurrence and, specifically, to the synchronisation between the occurrence of infected ticks and the corresponding host reservoirs of infection.
A. phagocytophilum was detected in 5.9% of questing ticks. Prevalences of
A. phagocytophilum in questing
I. ricinus ticks in Slovakia and neigbouring European countries vary between 0.57% and 13.1% [
30,
37,
59,
80,
81,
82]. As compared to recent works on
A. phagocytophilum prevalence for various urban and sylvatic habitats in Slovakia [
30,
37,
59], the pathogen prevalence was significantly higher for our urban forest study area (
Table 4).
Anaplasma phagocytophilum shows a considerable genetic variability [
83], which is partly explained by reservoir host specificity. In Europe,
A. phagocytophilum consists of two genetically distinct ecotypes that circulate in two enzootic cycles—one involving rodents and
I. trianguliceps ticks and the other involving ungulates, carnivores, and insectivores and
I. ricinus ticks [
37,
84,
85,
86]. Moreover, Blaňárová et al. [
37] showed that
I. ricinus ticks could not contract
A. phagocytophilum while feeding on infected rodents, though
I. trianguliceps ticks could contract the pathogen from infected rodents. Consequently, roe deer and hedgehogs do not only appear to represent the main host group for ticks, but also the main reservoir hosts of
A. phagocytophilum in the study area.
The circulation of the causative agents of Lyme borreliosis, spirochetes of
B. burgdorferi s.l. complex, depends on the presence of vector ticks and competent reservoir hosts. In our study we detected the presence of
B. burgdorferi s.l. in 6.8% (37/543) ticks. The prevalence of this pathogen complex for questing ticks in our study was significantly lower than those detected for various habitats in Slovakia including urban as well as natural sites [
26,
87] (
Table 5).
In addition, the mean prevalence of
B. burgdorferi s.l. in
I. ricinus across Europe (12.3–13.7%) [
13,
14] is about twice as high as that detected in our study area. Finally, our results on the pathogen prevalence are at the lower range limit for prevalences reported for neighbouring countries (4–25.7%) [
67,
88,
89,
90,
91,
92].
The low prevalence of
B. burgdorferi s.l. was perhaps the best reflected by the low prevalence of
B. afzelii; this rodent-associated genospecies was present only in 16.2% of borrelia-positive questing ticks. For example, at a different site in Slovakia, Derdáková et al. [
67] detected the presence of
B. afzelii in 56.5% of positive ticks. Rauter and Hartung [
13] revealed that on average 38% of borrelia-positive ticks across Europe are infected with
B. afzelii. Rodents represent the most important host group for the maintenance of juvenile
I. ricinus in Central and Eastern Europe [
93] and the key reservoir hosts for
B. afzelii, B. spielmanii, B. bavariensis and
B. burgdorferi s.s. [
19,
21,
22,
25]. Importantly, the presence of
B. afzelii,
B. burgdorferi s.s. or
B. spielmanii can also be maintained in reservoirs, such as hedgehogs and squirrels, especially in urban areas [
20,
24]. Consequently, the low infection rate of
B burgdorferi s.l., and particularly low prevalences of
B. afzelii,
B. spielmanii, and
B. burgdorferi s.s., support the findings of a previous study [
53] about a low abundance of small mammals at our study site. In fact, given the pattern of
B. garinii vs.
A. phagocytophilum and
B. valaisiana vs.
B. afzelii occurrences (RDA1 and RDA2 axes in
Figure 4), our results indicate that birds and roe deer represented main tick hosts at our study site. Morevover, given the pattern of RDA1 axis, reptiles appear to have served as alternative tick hosts during the periods of low bird abundance (
Figure 4). Importantly, ruminants such as roe deer may not only be an incompetent reservoir host for borrelia, but due to the borreliacidal effect of their blood complement, they can eliminate borrelia in feeding ticks [
39,
53,
94]. Overall, the low prevalence of
B. burgdorferi s.l. points to locally low densities of rodents, but also to the fact that birds and other small vertebrate hosts were not able to compensate for the lack of small rodents. Thus, roe deer appear to constitute one of the main host groups for
I. ricinus ticks and an important biotic factor behind the abundance and community structure of tick-borne pathogens at the study area.
B. garinii (35.1% prevalence) and
B. valaisiana (24.3% prevalence), which are associated with bird reservoirs, were found to be the locally dominant genospecies of
B. burgdorferi s.l. in questing ticks. Of trapped birds, 12.5% of bird-feeding ticks carried
B. burgdorferi s.l. (
Table 1) with the almost exclusive occurrence of
B. garinii. Tarageľová et al. [
26] detected 9.4% to 31.1% prevalence of
B. burgdorferi s.l. in bird-feeding ticks for different natural habitats in Slovakia. In that study, only 44% of borrelia-positive bird-feeding ticks carried
B. garinii, which was significantly less than we detected at our study site (
Table 6). This result confirms that birds represent the main
Borrelia reservoir at the study site. Our results, with respect to the prevalences of
A. phagocytophilum and
B. lusitaniae, also suggest that birds may not be available as hosts to questing tick cohorts during certain parts of the season when ruminants and reptiles seem to serve as the main tick hosts. These findings are consistent with the results by Tarageľová et al. [
16] who found for a montane habitat that
B. lusitaniae could be locally dominant
Borrelia genospecies under a shortage of tick hosts.
R. helvetica and
R. monacensis were recorded in 5.9% (28/477) of questing
I. ricinus ticks with
R. helvetica being the dominant
Rickettsia species. This prevalence value is again at the lower range limit of
Rickettsia prevalences detected in Slovakia and other European countries (0.5–66%) [
42,
43,
44]. The role of vertebrates in the life cycle of
R. helvetica is still unclear. Rickettsiae were identified in ticks feeding on rodents and birds [
95,
96,
97,
98], but also in blood and tissue samples from birds, rodents, roe deer, sika deer and wild boar [
46,
95,
98,
99,
100].
Rickettsia spp. is transmitted transovarially and transstadially. We can only speculate that rickettsial infections of attached ticks are the result of either a vertical route of infection (transovarial and/or transstadial transmission) in ticks or a very short rickettsiaemia in hosts or ticks co-feeding. While
I. ricinus can be considered as the most important reservoir host for Rickettsiae [
100], we found that the occurrence of
R. helvetica coincided with the occurrence of
B. afzelii (
Figure 4). Therefore, the enzootic cycle of
R. helvetica may be related to that of rodent-associated
B. afzelii. Moreover, our findings on a relatively low prevalence of
Rickettsia and
B. burgdorferi s.l. and a high prevalence of
A. phagocytophilum are consistent with the results by Václav et al. [
101]. Václav et al. [
101] found that
Anaplasma prevalence was negatively assotiated with the prevalences of
Rickettsia and
Borrelia. Thus, our study indicates that (1)
Rickettsia and
Borrelia circulate in similar hosts and (2) hosts responsible for
Anaplasma circulation contribute not only to the dillution of
Borrelia but also of
Rickettsia pathogens in feeding ticks.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}