Water-Use Characteristics and Physiological Response of Moso Bamboo to Flash Droughts

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Meteorological and ET Measurements
2.3. Data Analyses
3. Results
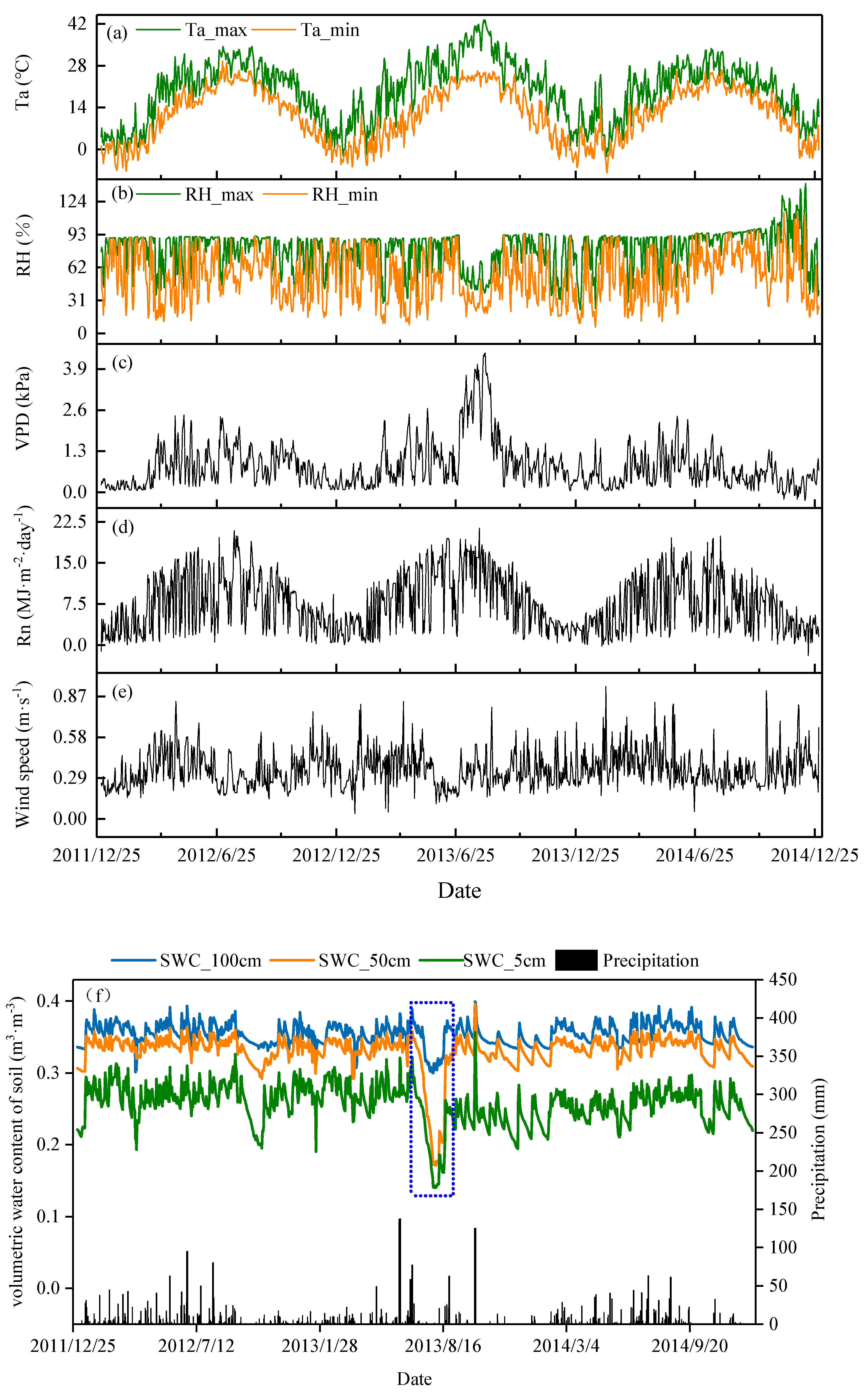
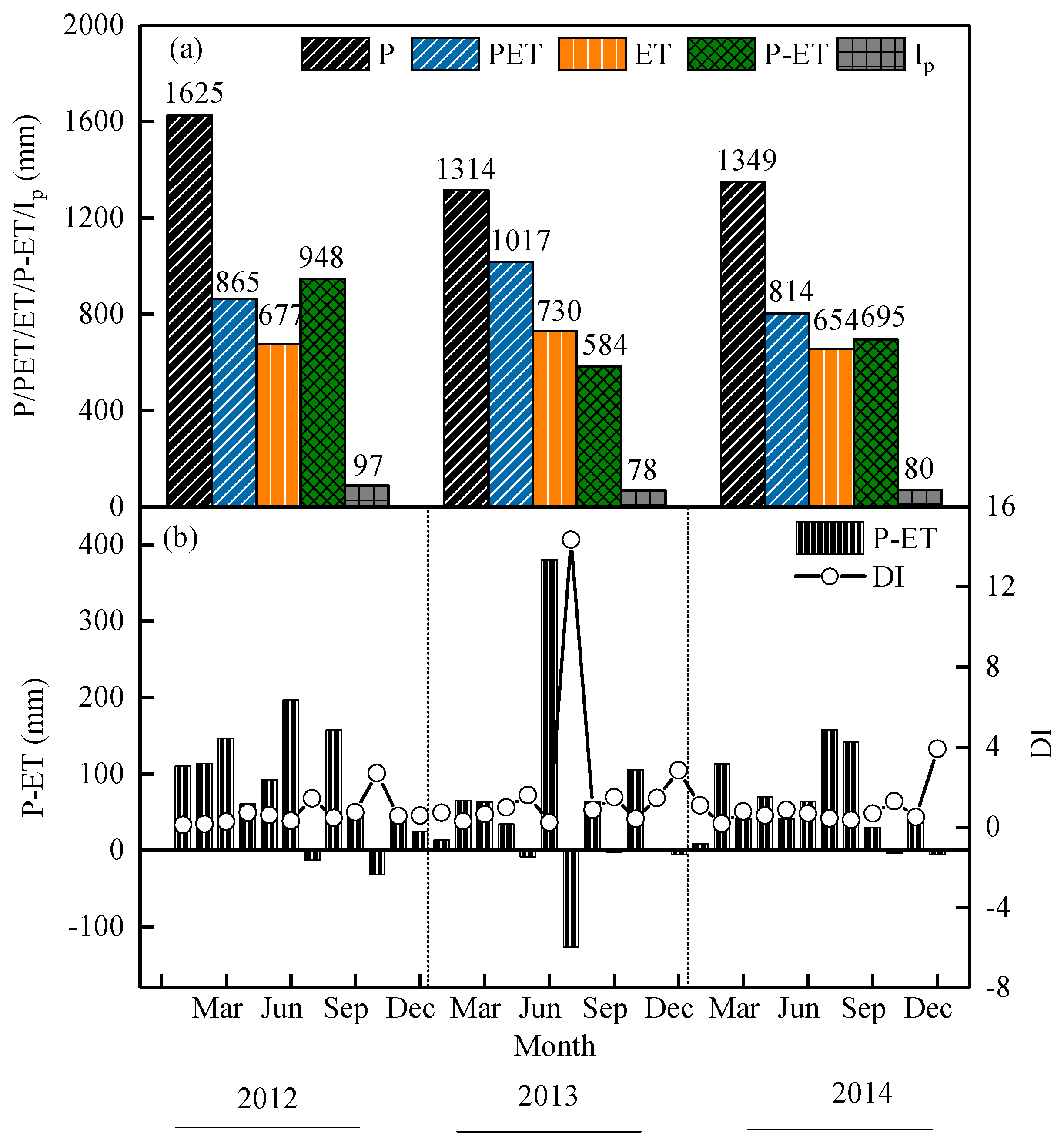
3.1. Meteorological Variables
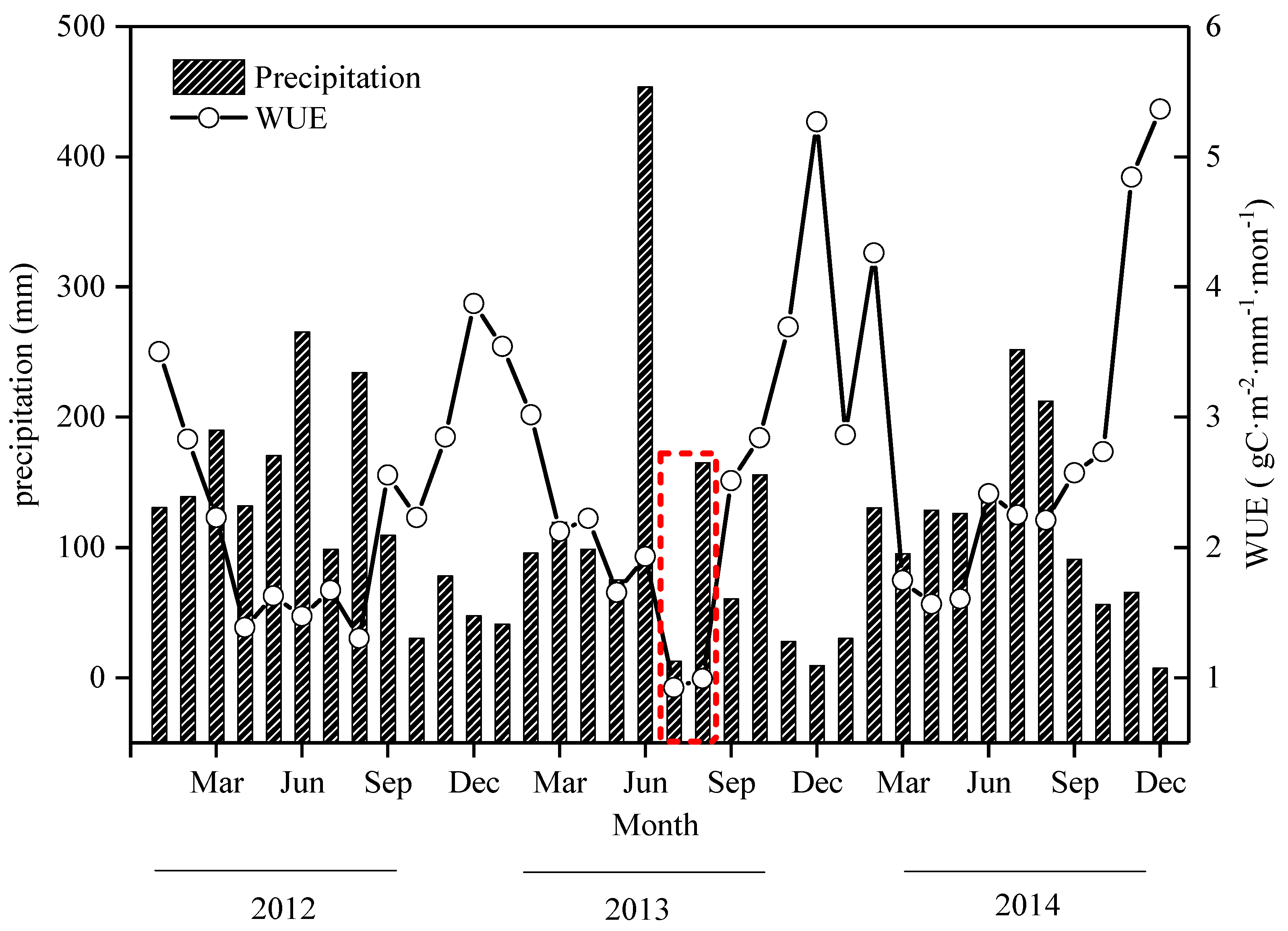
3.2. The Monthly Dynamic Characteristics of WUE and Precipitation
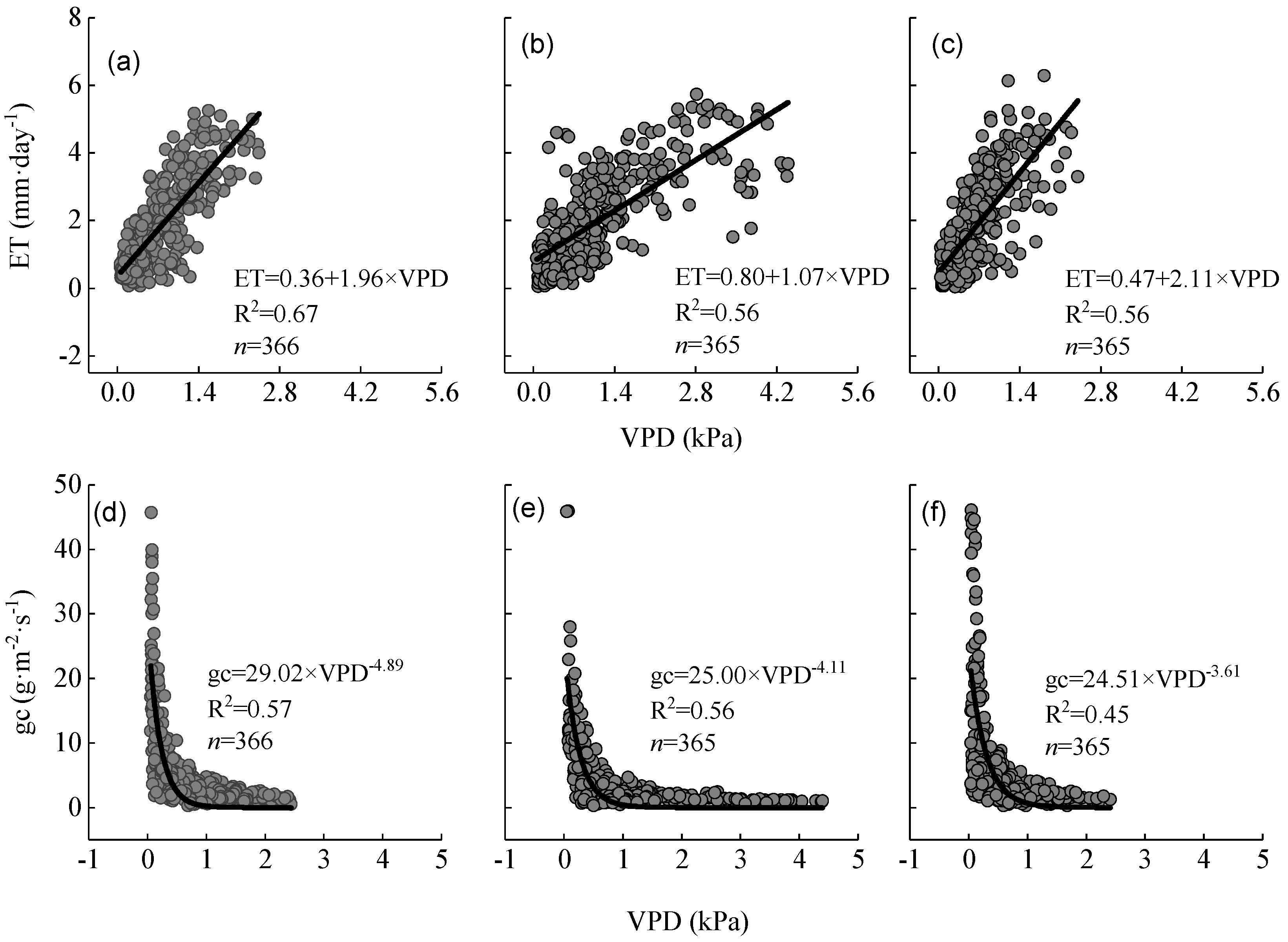
3.3. Dynamic Characteristics of ET, PET, and LAI
4. Discussion
4.1. The Response of SWC and LAI of Moso Bamboo Forests to Flash Drought.
4.2. Response of Evapotranspiration and Canopy Stomatal Conductance to Drought
4.3. Water Budgets and Partitioning of the Moso Bamboo Forest
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Hoerling, M.; Eischeid, J.; Kumar, A.; Leung, R.; Mariotti, A.; Mo, K.; Schubert, S.; Seager, R. Causes and Predictability of the 2012 Great Plains Drought. Bull. Am. Meteorol. Soc. 2014, 95, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yuan, X.; Xie, Z.; Wu, P.; Li, Y. Increasing flash droughts over China during the recent global warming hiatus. Sci. Rep. 2016, 6, 30571. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.L. Tropical forests and the changing earth system. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Z.; Chen, X.F.; Zhou, G.M.; Jiang, H.; Peng, C.H. Observed high and persistent carbon uptake by Moso bamboo forests and its response to environmental drivers. Agric. For. Meteorol. 2017, 247, 467–475. [Google Scholar] [CrossRef]
- Ph, C.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, V.; Wedin, D.A. Hydrological behaviour of grasslands of the Sandhills of Nebraska: Water and energy-balance assessment from measurements, treatments, and modelling. Ecohydrology 2009, 2, 195–212. [Google Scholar] [CrossRef]
- Lau, W.K.M.; Wu, H.T.; Kim, K.M. A canonical response of precipitation characteristics to global warming from CMIP5 models. Geophys. Res. Lett. 2013, 40, 3163–3169. [Google Scholar] [CrossRef]
- Huntington, T.G. Evidence for intensification of the global water cycle: Review and synthesis. J. Hydrol. 2006, 319, 83–95. [Google Scholar] [CrossRef]
- Chang, X.; Zhao, W.; Bing, L.; Hu, L.; He, Z.; Du, J.J. Can forest water yields be increased with increased precipitation in a Qinghai spruce forest in arid northwestern China? Agric. For. Meteorol. 2017, 247, 139–150. [Google Scholar] [CrossRef]
- Sawano, S.; Hotta, N.; Tanaka, N.; Tsuboyama, Y.; Suzuki, M. Development of a simple forest evapotranspiration model using a process-oriented model as a reference to parameterize data from a wide range of environmental conditions. Ecol. Model. 2015, 309–310, 93–109. [Google Scholar] [CrossRef]
- Wang, H.L.; Tetzlaff, D.; Dick, J.J.; Soulsby, C. Assessing the environmental controls on Scots pine transpiration and the implications for water partitioning in a boreal headwater catchment. Agric. For. Meteorol. 2017, 240, 58–66. [Google Scholar] [CrossRef]
- Chen, Y.; Xia, J.Z.; Liang, S.L.; Feng, J.M.; Fisher, J.B.; Li, X.; Li, X.; Liu, S.; Ma, Z.; Miyata, A.; et al. Comparison of satellite-based evapotranspiration models over terrestrial ecosystems in China. Remote Sens. Environ. 2014, 140, 279–293. [Google Scholar] [CrossRef]
- Song, X.; Peng, C.; Zhou, G.; Gu, H.; Li, Q.; Zhang, C. Dynamic allocation and transfer of non-structural carbohydrates, a possible mechanism for the explosive growth of Moso bamboo (Phyllostachys heterocycla). Sci. Rep. 2016, 6, 25908. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Jiang, P. Density, Storage and Spatial Distribution of Carbon in Phyllostachy pubescens Forest. Sci. Silvae Sin. 2004, 40, 20–24. [Google Scholar]
- Komatsu, H.; Onozawa, Y.; Kume, T.; Tsuruta, K.; Shinohara, Y.; Otsuki, K. Canopy conductance for a Moso bamboo (Phyllostachys pubescens) forest in western Japan. Agric. For. Meteorol. 2012, 156, 111–120. [Google Scholar] [CrossRef]
- Jiang, P.K.; Xu, Q.F.; Xu, Z.H.; Cao, Z.H. Seasonal changes in soil labile organic carbon pools within a Phyllostachys praecox stand under high rate fertilization and winter mulch in subtropical China. For. Ecol. Manag. 2006, 236, 30–36. [Google Scholar] [CrossRef]
- Zhao, X.H.; Ping, Z.; Zhang, Z.Z.; Zhu, L.W.; Niu, J.F.; Ni, G.Y.; Hu, Y.T.; Ouyang, L. Sap flow-based transpiration in Phyllostachys pubescens: Applicability of the TDP methodology, age effect and rhizome role. Trees 2017, 31, 765–779. [Google Scholar]
- Xing, Y.; Ma, Z.; Ming, P.; Shi, C. Microwave remote sensing of short-term droughts during crop growing seasons. Geophys. Res. Lett. 2015, 42, 4394–4401. [Google Scholar]
- Xu, X.J.; Du, H.Q.; Zhou, G.M.; Li, P.H.; Shi, Y.J.; Zhou, Y.F. Eddy covariance analysis of the implications of drought on the carbon fluxes of Moso bamboo forest in southeastern China. Trees-Struct. Funct. 2016, 30, 1807–1820. [Google Scholar] [CrossRef]
- Ge, X.; Zhou, B.; Wang, X.; Qian, L.; Cao, Y.; Gu, L. Imposed drought effects on carbon storage of moso bamboo ecosystem in southeast China: Results from a field experiment. Ecol. Res. 2018, 33, 393–402. [Google Scholar] [CrossRef]
- Piouceau, J.; Panfili, F.; Bois, G.; Anastase, M.; Dufosse, L.; Arfi, V. Actual evapotranspiration and crop coefficients for five species of three-year-old bamboo plants under a tropical climate. Agric. Water Manag. 2014, 137, 15–22. [Google Scholar] [CrossRef]
- Chen, S.L.; Jiang, H.; Cai, Z.J.; Zhou, X.L.; Peng, C.H. The response of the net primary production of Moso bamboo forest to the On and Off-year management: A case study in Anji County, Zhejiang, China. For. Ecol. Manag. 2018, 409, 1–7. [Google Scholar] [CrossRef]
- Kunert, N.; Aparecido, L.M.T.; Wolff, S.; Higuchi, N.; Santos, J.D.; Araujo, A.C.D.; Trumbore, S. A revised hydrological model for the Central Amazon: The importance of emergent canopy trees in the forest water budget. Agric. For. Meteorol. 2017, 239, 47–57. [Google Scholar] [CrossRef]
- Yuli, L.; Hong, J.; Guomo, Z.; Xiaofeng, C.; Chen, S.; Shuang, Y. Water vapor flux variation characteristic and the relationship with its environment factors in phyllostachys edulis forest in Anji. Acta Ecol. Sin. 2014, 34, 4900–4909. [Google Scholar]
- Liu, Y.; Zhou, G.; Du, H.; Berninger, F.; Mao, F.; Li, X.; Chen, L.; Cui, L.; Li, Y.; Xu, L. Response of carbon uptake to abiotic and biotic drivers in an intensively managed Lei bamboo forest. J. Environ. Manag. 2018, 223, 713–722. [Google Scholar] [CrossRef]
- Aubinet, M.; Vesala, T.; Papale, D. Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Springer: Dordrecht, The Netherlands, 2012; pp. 365–376. [Google Scholar]
- Brutsaert, W.H. Evaporation into the Atmosphere, Theory, History and Applications; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1982. [Google Scholar]
- Nunn, A.J.; Wieser, G.; Metzger, U.; Low, M.; Wipfler, P.; Haberle, K.H.; Matyssek, R. Exemplifying whole-plant ozone uptake in adult forest trees of contrasting species and site conditions. Environ. Pollut. 2007, 146, 629–639. [Google Scholar] [CrossRef]
- Huang, M.; Piao, S.; Zeng, Z.; Peng, S.; Ciais, P.; Cheng, L.; Mao, J.; Poulter, B.; Shi, X.; Yao, Y.; et al. Seasonal responses of terrestrial ecosystem water-use efficiency to climate change. Glob. Chang. Biol. 2016, 22, 2165–2177. [Google Scholar] [CrossRef]
- Jin, J.; Ying, W.; Zhen, Z.; Magliulo, V.; Hong, J.; Min, C. Phenology Plays an Important Role in the Regulation of Terrestrial Ecosystem Water-Use Efficiency in the Northern Hemisphere. Remote Sens. 2017, 9, 664. [Google Scholar] [CrossRef]
- Milne, B.T.; Gupta, V.K.; Restrepo, C. A scale invariant coupling of plants, water, energy, and terrain. Ecoscience 2002, 9, 191–199. [Google Scholar] [CrossRef]
- Rajib, A.; Merwade, V.; Yu, Z.Q. Rationale and efficacy of assimilating remotely sensed potential evapotranspiration for reduced uncertainty of hydrologic models. Water Resour. Res. 2018, 54, 4615–4637. [Google Scholar] [CrossRef]
- Allen, R.G.; Pruitt, W.O.; Wright, J.L.; Howell, T.A.; Ventura, F. A recommendation on standardized surface resistance for hourly calculation of reference ETo by the FAO56 Penman-Monteith method. Agric. Water Manag. 2006, 81, 1–22. [Google Scholar] [CrossRef]
- Qiang, T.; Hu, H.; Tian, F.; Guan, H.; Lin, H. Environmental and physiological controls on sap flow in a subhumid mountainous catchment in North China. Agric. For. Meteorol. 2017, 240, 46–57. [Google Scholar]
- Small, E.E.; Mcconnell, J.R.J. Comparison of soil moisture and meteorological controls on pine and spruce transpiration. Ecohydrology 2010, 1, 205–214. [Google Scholar] [CrossRef]
- Granier, A.; Reichstein, M.; Breda, N.; Janssens, I.A.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Keenan, T.; Sabate, S.; Gracia, C. The importance of mesophyll conductance in regulating forest ecosystem productivity during drought periods. Glob. Chang. Biol. 2010, 16, 1019–1034. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y. Hydrological characteristics of a moso-bamboo (Phyllostachys pubescens) forest in south china. Hydrol. Process. 2010, 9, 797–808. [Google Scholar] [CrossRef]
- Kumagai, T.; Saitoh, T.M.; Sato, Y.; Morooka, T.; Manfroi, O.J.; Kuraji, K.; Suzuki, M. Transpiration, canopy conductance and the decoupling coefficient of a lowland mixed dipterocarp forest in Sarawak, Borneo: Dry spell effects. J. Hydrol. 2004, 287, 237–251. [Google Scholar] [CrossRef]
- Morton, D.C.; Nagol, J.; Carabajal, C.C.; Rosette, J.; Palace, M.; Cook, B.D.; Vermote, E.F.; Harding, D.J.; North, P.R. Amazon forests maintain consistent canopy structure and greenness during the dry season. Nature 2014, 506, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Y.; Wang, Y.K.; Liu, S.Y.; Wei, X.G.; Wang, X. Response of relative sap flow to meteorological factors under different soil moisture conditions in rainfed jujube (Ziziphus jujuba Mill.) plantations in semiarid Northwest China. Agric. Water Manag. 2014, 136, 23–33. [Google Scholar] [CrossRef]
- Schwinning, S.; Davis, K.; Richardson, L.; Ehleringer, J.R. Deuterium enriched irrigation indicates different forms of rain use in shrub/grass species of the Colorado Plateau. Oecologia 2002, 130, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.X.; Zhang, Z.Q.; Zha, T.G.; Mo, K.L.; Zhang, Y.; Fang, X.R. Soil water affects transpiration response to rainfall and vapor pressure deficit in poplar plantation. New For. 2014, 45, 235–250. [Google Scholar] [CrossRef]
- Asko, N.; Mcnulty, S.G.; Deforest, J.L.; Ge, S.; Qinglin, L.; Jiquan, C. Drought during canopy development has lasting effect on annual carbon balance in a deciduous temperate forest. New Phytol. 2010, 179, 818–828. [Google Scholar]
- Gartner, K.; Nadezhdina, N.; Englisch, M.; Cermak, J.; Leitgeb, E. Sap flow of birch and Norway spruce during the European heat and drought in summer 2003. For. Ecol. Manag. 2009, 258, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Pataki, D.E.; Oren, R.; Smith, W.K. Sap Flux of Co-Occurring Species in a Western Subalpine Forest during Seasonal Soil Drought. Aspen Bibliogr. 2000, 81, 2557–2566. [Google Scholar] [CrossRef]
- Clausnitzer, F.; Köstner, B.; Kai, S.; Bernhofer, C. Relationships between canopy transpiration, atmospheric conditions and soil water availability—Analyses of long-term sap-flow measurements in an old Norway spruce forest at the Ore Mountains/Germany. Agric. For. Meteorol. 2011, 151, 1023–1034. [Google Scholar] [CrossRef]
- Lagergren, F.; Lindroth, A. Transpiration response to soil moisture in pine and spruce trees in Sweden. Agric. For. Meteorol. 2002, 112, 67–85. [Google Scholar] [CrossRef]
- Yu, G.; Song, X.; Wang, Q.; Liu, Y.; Guan, D.; Yan, J.; Sun, X.; Zhang, L.; Wen, X. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2010, 177, 927–937. [Google Scholar] [CrossRef]
- Reichstein, M.; Tenhunen, J.O.; Ourcival, J.M.; Rambal, S. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: Revision of current hypotheses? Glob. Chang. Biol. 2010, 8, 999–1017. [Google Scholar] [CrossRef]
- Lu, X.; Zhuang, Q. Evaluating evapotranspiration and water-use efficiency of terrestrial ecosystems in the conterminous United States using MODIS and AmeriFlux data. Remote Sens. Environ. 2010, 114, 1924–1939. [Google Scholar] [CrossRef]
- Bovard, B.D.; Curtis, P.S.; Vogel, C.S.; Su, H.B.; Schmid, H.P. Environmental controls on sap flow in a northern hardwood forest. Tree Physiol. 2005, 25, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, P.J.; Drake PLFroend, R.H. Anisohydric but isohydrodynamic: Seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance. Plant Cell Environ. 2010, 30, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.J.; Farquhar, G.D. A relationship between humidity response, growth form and photosynthetic operating point in C3 plants. Plant Cell Environ. 2010, 22, 1337–1349. [Google Scholar] [CrossRef]
- Komatsu, H.; Onozawa, Y.; Kume, T.; Tsuruta, K.; Kumagai, T.; Shinohara, Y.; Otsuki, K. Stand-scale transpiration estimates in a Moso bamboo forest: II. Comparison with coniferous forests. For. Ecol. Manag. 2010, 260, 1295–1302. [Google Scholar] [CrossRef]
- Ichihashi, R.; Komatsu, H.; Kume, T.; Onozawa, Y.; Shinohara, Y.; Tsuruta, K.; Otsuki, K. Stand-scale transpiration of two Moso bamboo stands with different culm densities. Ecohydrology 2015, 8, 450–459. [Google Scholar] [CrossRef]
- Barbour, M.M.; Hunt, J.E.; Walcroft, A.S.; Rogers, G.N.D.; McSeveny, T.M.; Whitehead, D. Components of ecosystem evaporation in a temperate coniferous rainforest, with canopy transpiration scaled using sapwood density. New Phytol. 2005, 165, 549–558. [Google Scholar] [CrossRef]
- Sridhar, V.; Hubbard, K.G.; Wedin, D.A. Assessment of soil moisture dynamics of the Nebraska Sandhills using long-term measurements and a hydrology model. J. Irrig. Drain. Eng. 2006, 132, 463–473. [Google Scholar] [CrossRef]
- Tomer, M.D.; Schilling, K.E. A simple approach to distinguish land-use and climate-change effects on watershed hydrology. J. Hydrol. 2009, 376, 24–33. [Google Scholar] [CrossRef]
- Rajib, A.; Merwade, V. Hydrologic response to future land use change in the Upper Mississippi River Basin by the end of 21st century. Hydrol. Process. 2017, 31, 3645–3661. [Google Scholar] [CrossRef]
- Mehan, S.; Kannan, N.; Neupane, R.P.; McDaniel, R.; Kumar, S. Climate Change Impacts on the Hydrological Processes of a Small Agricultural Watershed. Climate 2016, 4, 56. [Google Scholar] [CrossRef]
- Shinohara, Y.; Komatsu, H.; Kuramoto, K.; Otsuki, K. Characteristics of canopy interception loss in Moso bamboo forests of Japan. Hydrol. Process. 2013, 27, 2041–2047. [Google Scholar] [CrossRef]
- Ellison, D.; Futter, M.N.; Bishop, K. On the forest cover–water yield debate: From demand- to supply-side thinking. Glob. Chang. Biol. 2012, 18, 806–820. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Chen, S.; Jiang, H.; Lin, Y.; Zhang, J.; Song, X.; Zhou, G. Water-Use Characteristics and Physiological Response of Moso Bamboo to Flash Droughts. Int. J. Environ. Res. Public Health 2019, 16, 2174. https://doi.org/10.3390/ijerph16122174
Zhang M, Chen S, Jiang H, Lin Y, Zhang J, Song X, Zhou G. Water-Use Characteristics and Physiological Response of Moso Bamboo to Flash Droughts. International Journal of Environmental Research and Public Health. 2019; 16(12):2174. https://doi.org/10.3390/ijerph16122174
Chicago/Turabian StyleZhang, Minxia, Shulin Chen, Hong Jiang, Yong Lin, Jinmeng Zhang, Xinzhang Song, and Guomo Zhou. 2019. "Water-Use Characteristics and Physiological Response of Moso Bamboo to Flash Droughts" International Journal of Environmental Research and Public Health 16, no. 12: 2174. https://doi.org/10.3390/ijerph16122174