The Toxic Effect of Herbicidal Ionic Liquids on Biogas-Producing Microbial Community

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ionic Liquids
2.2. Anaerobic Digestion
2.3. Determination of Herbicidal Ionic Liquids
2.3.1. Sample Preparation
2.3.2. HPLC-MS Analysis
2.4. Metagenomic Analysis
2.4.1. DNA Extraction
2.4.2. PCR Amplification
2.4.3. Bioinformatic Analysis
3. Results and Discussion
3.1. The Effect of Herbicidal Ionic Liquids on the Amount of Obtained Biogas
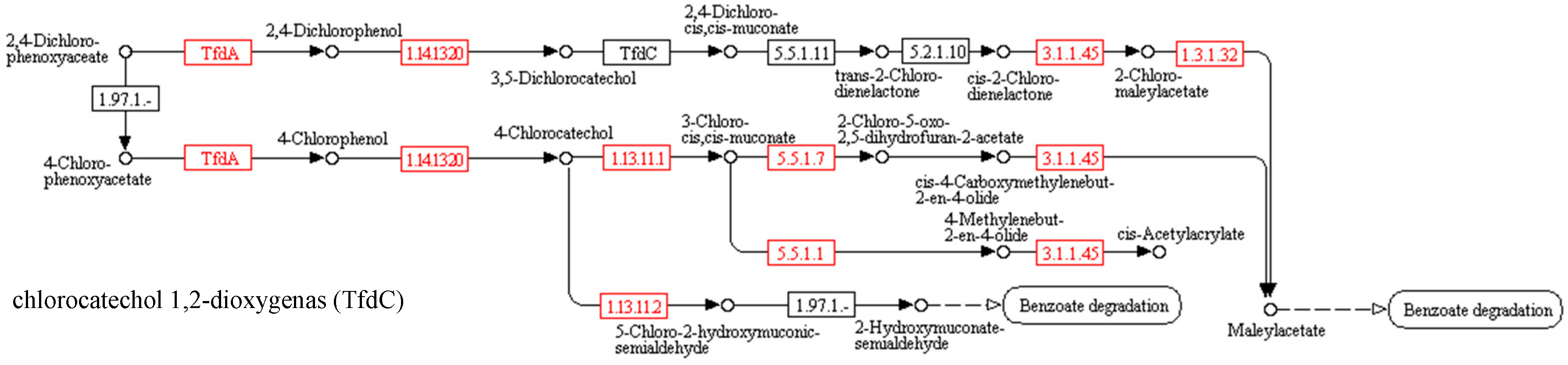
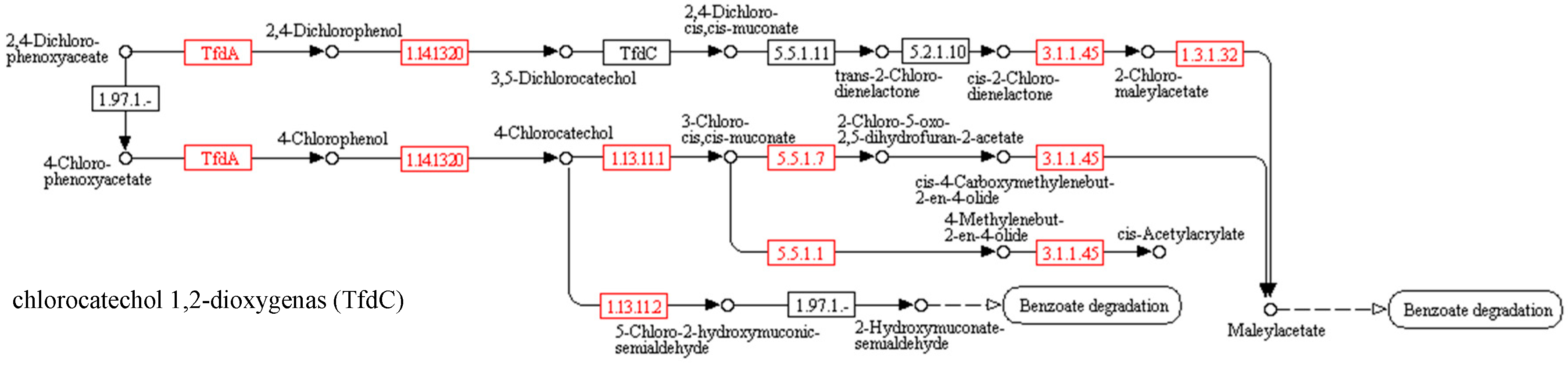
3.2. Biodegradation of Herbicidal Ionic Liquids
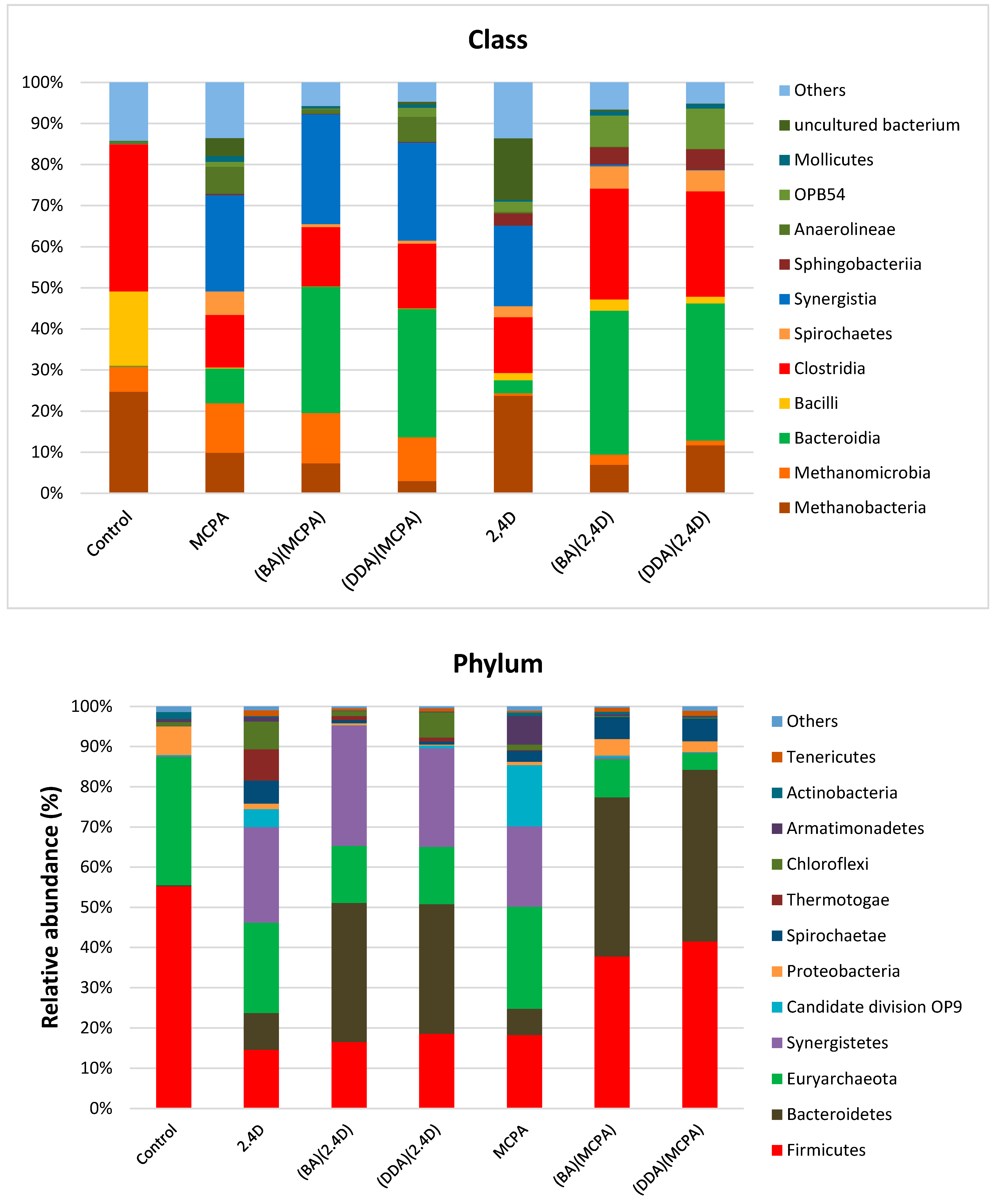
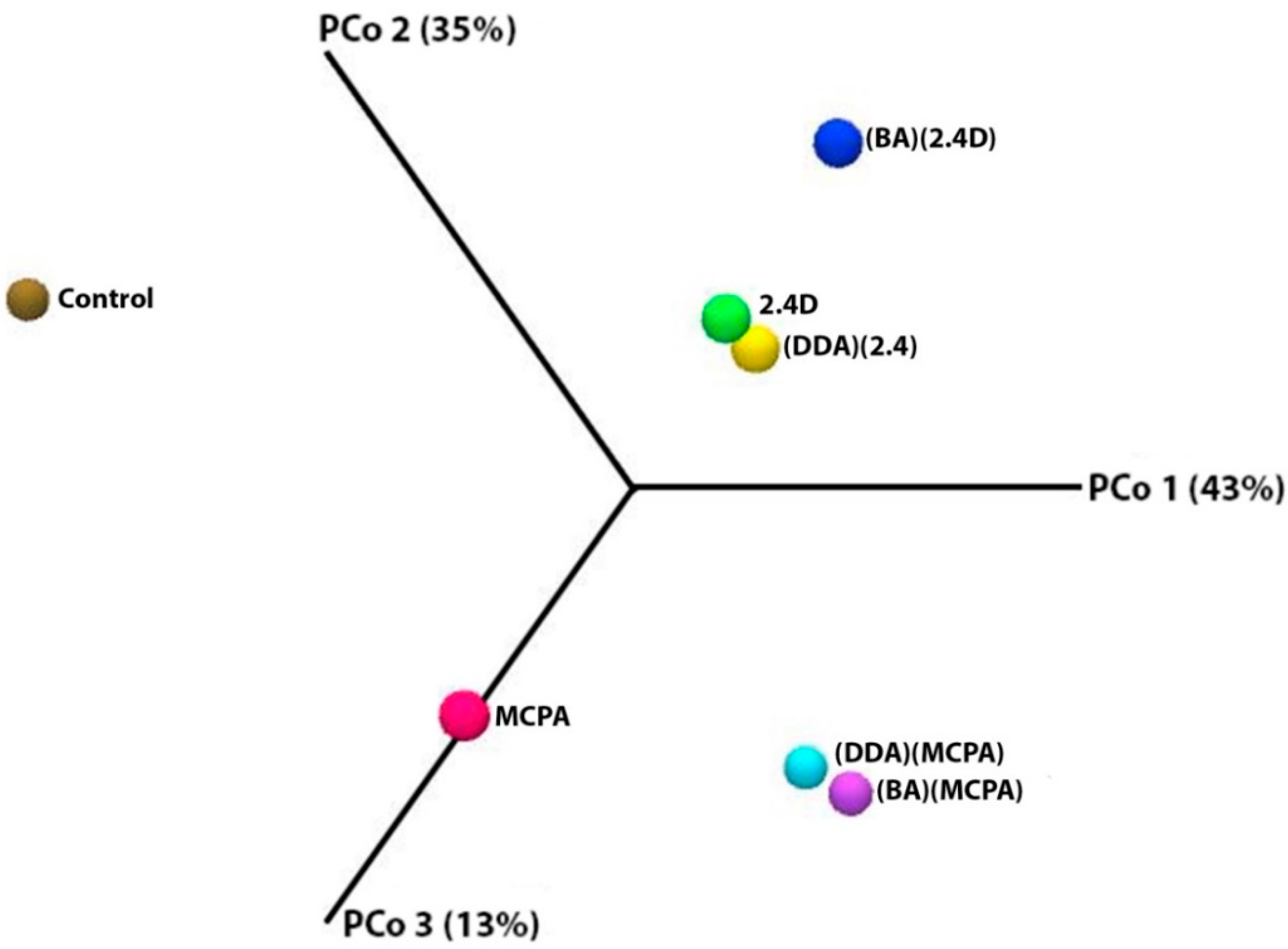
3.3. Metapopulation Analysis
Predicted functional gene abundance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cieslik, M.; Dach, J.; Lewicki, A.; Smurzynska, A.; Janczak, D.; Pawlicka-Kaczorowska, J.; Boniecki, P.; Cyplik, P.; Czekala, W.; Jozwiakowski, K. Methane fermentation of the maize straw silage under meso- and thermophilic conditions. Energy 2016, 115, 1495–1502. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Anjum, M.; Mahmood, T.; Dawson, L. Review—The anaerobic digestion ofsolidorganicwaste. Waste Manag. 2011, 31, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Ortiz, R.; Steele, T.W.J.; Stuckey, D.C. Toxicants inhibiting anaerobic digestion: A review. Biotechnol. Adv. 2014, 32, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Kwietniewska, E.; Tys, J. Process characteristics, inhibition factors and methane yields of anaerobic digestion process, with particular focus on microalgal biomass fermentation. Renew. Sustain. Energ. Rev. 2014, 34, 491–500. [Google Scholar] [CrossRef]
- Piotrowska-Cyplik, A.; Czarnecki, Z. Phytoextraction of heavy metals by hemp during anaerobic sewage sludge management in the non-industrial sites. Pol. J. Environ. Stud. 2003, 12, 779–784. [Google Scholar]
- Ward, A.J.; Hobbs, P.J.; Holliman, P.J.; Jones, D.L. Optimisation of the anaerobic digestion of agricultural resources. Bioresour. Technol. 2008, 99, 7928–7940. [Google Scholar] [CrossRef]
- Osorio, F.; Torres, J.C. Biogas purification from anaerobic digestion in a wastewater treatment plant for biofuel production. Renew. Energ. 2009, 34, 2164–2171. [Google Scholar] [CrossRef]
- Yenigün, O.; Demirel, B. Ammonia inhibition in anaerobic digestion: A review. Process. Biochem. 2013, 48, 901–911. [Google Scholar] [CrossRef]
- Siegert, I.; Banks, C. The effect of volatile fatty acid additions on the anaerobic digestion of cellulose and glucose in batch reactors. Process. Biochem. 2005, 40, 3412–3418. [Google Scholar] [CrossRef]
- Aydin, S.; Ince, B.; Ince, O. Assessment of anaerobic bacterial diversity and its effects on anaerobic system stability and the occurrence of antibiotic resistance genes. Bioresour. Technol. 2016, 207, 332–338. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Syguda, A.; Gielnik, A.; Borkowski, A.; Woźniak-Karczewska, M.; Parus, A.; Piechalak, A.; Olejnik, A.; Marecik, R.; Ławniczak, Ł.; Chrzanowski, Ł. Esterquat herbicidal ionic liquids (HILs) with two different herbicides: Evaluation of activity and phytotoxicity. New J. Chem. 2018, 42, 9819–9827. [Google Scholar] [CrossRef]
- Pernak, J.; Łęgosz, B.; Walkiewicz, F.; Klejdysz, T.; Borkowski, A.; Chrzanowski, L. Ammonium ionic liquids with anions of natural origin. RSC Adv. 2015, 5, 65471–65480. [Google Scholar] [CrossRef]
- Pernak, J.; Niemczak, M.; Chrzanowski, Ł.; Ławniczak, Ł.; Fochtman, P.; Marcinkowska, K.; Praczyk, T. Betaine and Carnitine Derivatives as Herbicidal Ionic Liquids. Chem. A Eur. J. 2016, 22, 12012–12021. [Google Scholar] [CrossRef]
- Ghandi, K. A Review of Ionic Liquids, Their Limits and Applications. Green Sustain. Chem. 2014, 4, 44–53. [Google Scholar] [CrossRef]
- Pernak, J.; Syguda, A.; Janiszewska, D.; Materna, K.; Praczyk, T. Ionic liquids with herbicidal anions. Tetrahedron 2011, 67, 4838–4844. [Google Scholar] [CrossRef]
- Pernak, J.; Syguda, A.; Materna, K.; Janus, E.; Kardasz, P.; Praczyk, T. 2,4-D based herbicidal ionic liquids. Tetrahedron 2012, 68, 4267–4273. [Google Scholar] [CrossRef]
- Niemczak, M.; Chrzanowski, Ł.; Praczyk, T.; Pernak, J. Biodegradable herbicidal ionic liquids based on synthetic auxins and analogues of betainę. New J. Chem. 2017, 41, 8066–8077. [Google Scholar] [CrossRef]
- Piotrowska, A.; Syguda, A.; Wyrwas, B.; Chrzanowski, L.; Luckenbach, T.; Heipieper, H.J. Effects of ammonium-based ionic liquids and 2,4-dichlorophenol on the phospholipid fatty acid composition of zebrafish embryos. PLoS ONE 2018, 1, e0190779. [Google Scholar] [CrossRef]
- Ławniczak, Ł.; Materna, K.; Framski, G.; Szulc, A.; Syguda, A. Comparative study on the biodegradability of morpholinium herbicidal ionic liquids. Biodegradation 2015, 26, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Cyplik, P.; Juzwa, W.; Marecik, R.; Powierska-Czarny, J.; Piotrowska-Cyplik, A.; Czarny, J.; Drozdzynska, A.; Chrzanowski, L. Denitrification of industrial wastewater: Influence of glycerol addition on metabolic activity and community shifts in a microbial consortium. Chemosphere 2013, 93, 2823–2831. [Google Scholar] [CrossRef]
- Szczepaniak, Z.; Cyplik, P.; Juzwa, W.; Czarny, J.; Staninska, J.; Piotrowska-Cyplik, A. Antibacterial effect of the Trichoderma viride fungi on soil microbiome during PAH’s biodegradation. Int. Biodeterior. Biodegrad. 2015, 104, 170–177. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Ławniczak, Ł.; Syguda, A.; Borkowski, A.; Cyplik, P.; Marcinkowska, K.; Wolko, Ł.; Praczyk, T.; Chrzanowski, Ł.; Pernak, J. Influence of oligomeric herbicidal ionic liquids with MCPA and Dicamba anions on the community structure of autochthonic bacteria present in agricultural soil. Sci. Total Environ. 2016, 563–564, 247–255. [Google Scholar] [CrossRef]
- Sanchis, S.; Polo, A.M.; Tobajas, M.; Rodriguez, J.J.; Mohedano, A.F. Strategies to evaluate biodegradability: Application to chlorinated herbicides. Environ. Sci. Pollut. Res. 2014, 21, 9445–9452. [Google Scholar] [CrossRef]
- Fargašová, A. Comparative study of plant growth hormone (herbicide) toxicity in various biological subjects. Ecotoxicol. Environ. Saf. 1994, 29, 359–364. [Google Scholar] [CrossRef]
- Chakraborty, N.; Sarkar, G.M.; Lahiri, S.C. Effect of pesticide (Tara-909) on biomethanation of sewage sludge and isolated methanogens. Biomass Bioenergy 2002, 23, 75–80. [Google Scholar] [CrossRef]
- Kupper, T.; Bucheli, T.D.; Brändli, R.C.; Ortelli, D.; Edder, P. Dissipation of pesticides during composting and anaerobic digestion of source-separated organic waste at full-scale plants. Bioresour. Technol. 2008, 99, 7988–7994. [Google Scholar] [CrossRef]
- Luo, G.; Angelidaki, I. Analysis of bacterial communities and bacterial pathogens in a biogas plant by the combination of ethidium monoazide, PCR and Ion Torrent sequencing. Water Res. 2014, 60, 156–163. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.; Si, D.; Chen, Q. Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresour. Technol. 2016, 216, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, Z.; Czarny, J.; Staninska-Pieta, J.; Lisiecki, P.; Zgola-Grzeskowiak, A.; Cyplik, P.; Chrzanowski, Ł.; Wolko, Ł.; Marecik, R.; Glazar, K.; et al. Influence of soil contamination with PAH on microbial community dynamics and expression level of genes responsible for biodegradation of PAH and production of rhamnolipids. Environ. Sci. Pollut. Res. 2016, 23, 23043–23056. [Google Scholar] [CrossRef] [Green Version]
- Wyrwas, B.; Dymaczewski, Z.; Zgola-Grzeskowiak, A.; Szymanski, A.; Franska, M.; Kruszelnicka, I.; Ginter-Kramarczyk, D.; Cyplik, P.; Ławniczak, L.; Chrzanowski, Ł. Biodegradation of Triton X-100 and its primary metabolites by a bacterial community isolated from activated sludge. J. Environ. Manag. 2013, 128, 292–299. [Google Scholar] [CrossRef]
- Sydow, M.; Owsianiak, M.; Framski, G.; Woźniak-Karczewska, M.; Piotrowska-Cyplik, A.; Ławniczak, Ł.; Szulc, A.; Zgoła-Grześkowiak, A.; Heipieper, H.J.; Chrzanowski, Ł. Biodiversity of soil bacteria exposed to sub-lethal concentrations of phosphonium-based ionic liquids: Effects of toxicity and biodegradation. Ecotoxicol. Environ. Saf. 2018, 147, 157–164. [Google Scholar] [CrossRef]
- Gimeno, O.; Aguinaco, A.; Rey, A.; Novillo, F.J.B.; Toledo, J.R. Ozonation of 4-chloro-2-methylphenoxyacetic acid (MCPA) in an activated sludge system. J. Chem. Technol. Biotechnol. 2014, 89, 1219–1227. [Google Scholar] [CrossRef]
- Celis, E.; Elefsiniotis, P.; Singhal, N. Biodegradation of agricultural herbicides in sequencing batch reactors under aerobic or anaerobic conditions. Water Res. 2008, 42, 3218–3224. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HIL/Herbicide | Chemical Structure | Molecular Mass (g/mol) |
|---|---|---|
| (BA)(2,4-D) |  | 540.64 |
| (BA)(MCPA) |  | 520.27 |
| (DDA)(2,4-D) | 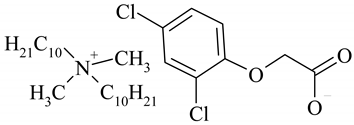 | 546.04 |
| (DDA)(MCPA) | 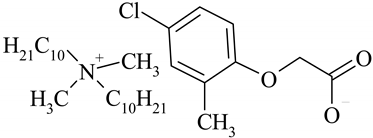 | 526.33 |
| 2,4-D |  | 243.02 |
| MCPA |  | 222.61 |
| Analyte | MRM Transitions (Precursor Ionm/z→Product Ion m/z) | |||
|---|---|---|---|---|
| MRM 1 | Collision Energy (V) | MRM 2 | Collision Energy (V) | |
| (BA) | 304→91 | 35 | 304→212 | 29 |
| (DDA) | 326→186 | 38 | - | - |
| (MCPA) | 199→141 | −23 | 199→155 | −15 |
| (2,4-D) | 219→161 | −30 | 219→125 | −37 |
| Herbicidal Ionic Liquids and Herbicides | EC50 | Liquid | Biomass | ||
|---|---|---|---|---|---|
| Cation | Anion | Cation | Anion | ||
| (DDA)(MCPA) | 38.6 | 4.1 ± 1.4 | 44.3 ± 6.9 | 52.5 ± 6.5 | 2.8 ± 1.6 |
| (DDA)(2,4-D) | 59.8 | 2.4 ± 1.6 | 37.4 ± 5.2 | 40.6 ± 4.6 | 6.4 ± 3.6 |
| (BA)(2,4-D) | 55.7 | 3.3 ± 2.3 | 45.4 ± 8.7 | 5.2 ± 2.1 | 1.7 ± 0.9 |
| (BA)(MCPA) | 40.2 | 0.4 ± 0.3 | 45.0 ± 9.3 | 42,0 ± 2.1 | 7.3 ± 2.8 |
| MCPA | 45.7 | - | 38.6 ± 6.8 | - | 9.3 ± 5.3 |
| 2,4-D | 65.8 | - | 48.6 ± 7.9 | - | 6.2 ± 4.6 |
| Alpha-Biodiversity Ratios | Control | MCPA | (BA)(MCPA) | (DDA)(MCPA) | 2,4-D | (BA)(2,4-D) | (DDA)(2,4-D) |
|---|---|---|---|---|---|---|---|
| Number of OTU | 366 | 291 | 295 | 275 | 296 | 301 | 317 |
| Shannon’s entropy | 5.4 | 4.4 | 4.45 | 4.6 | 4.9 | 4.8 | 5.0 |
| EC Number | Abbreviations of Names of Enzymes | Names of Enzymes | Control | MCPA | (BA)(MCPA) | (DDA)(MCPA) | 2,4D | (BA)(2,4-D) | (DDA)(2,4-D) |
|---|---|---|---|---|---|---|---|---|---|
| EC 1.14.11.- | tfdA | alpha-ketoglutarate-dependent 2,4-dichlorophenoxyacetate dioxygenase | 0.30 | 0.00 | 0.00 | 0.00 | 0.01 | 0.12 | 0.07 |
| EC 1.14.13.20 | tfdB | 2,4-dichlorophenol 6-monooxygenase | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 |
| EC 3.1.1.45 | E3.1.1.45 (tfdE) | carboxymethylenebutenolidase | 8.03 | 10.98 | 5.48 | 7.32 | 2.35 | 3.62 | 2.91 |
| EC 5.5.1.1 | catB | muconatecycloisomerase | 2.65 | 0.52 | 0.29 | 0.19 | 0.60 | 1.06 | 1.18 |
| EC 5.5.1.7 | E5.5.1.7 (tfdD) | chloromuconatecycloisomerase | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| EC 1.3.1.32 | E1.3.1.32 (tfdF) | maleylacetate reductase | 0.33 | 0.01 | 0.00 | 0.00 | 0.03 | 0.15 | 0.07 |
| EC 1.13.11.1 | catA | catechol 1,2-dioxygenase | 0.54 | 0.03 | 0.00 | 0.01 | 0.07 | 0.51 | 0.31 |
| EC 1.13.11.2 | dmpB | catechol 2,3-dioxygenase | 0.52 | 3.47 | 0.47 | 3.21 | 0.30 | 0.42 | 0.27 |
| EC 1.13.11.2 | catE | catechol 2,3-dioxygenase | 1.69 | 3.64 | 0.64 | 3.22 | 0.87 | 0.73 | 0.30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czarny, J.; Piotrowska-Cyplik, A.; Lewicki, A.; Zgoła-Grześkowiak, A.; Wolko, Ł.; Galant, N.; Syguda, A.; Cyplik, P. The Toxic Effect of Herbicidal Ionic Liquids on Biogas-Producing Microbial Community. Int. J. Environ. Res. Public Health 2019, 16, 916. https://doi.org/10.3390/ijerph16060916
Czarny J, Piotrowska-Cyplik A, Lewicki A, Zgoła-Grześkowiak A, Wolko Ł, Galant N, Syguda A, Cyplik P. The Toxic Effect of Herbicidal Ionic Liquids on Biogas-Producing Microbial Community. International Journal of Environmental Research and Public Health. 2019; 16(6):916. https://doi.org/10.3390/ijerph16060916
Chicago/Turabian StyleCzarny, Jakub, Agnieszka Piotrowska-Cyplik, Andrzej Lewicki, Agnieszka Zgoła-Grześkowiak, Łukasz Wolko, Natalia Galant, Anna Syguda, and Paweł Cyplik. 2019. "The Toxic Effect of Herbicidal Ionic Liquids on Biogas-Producing Microbial Community" International Journal of Environmental Research and Public Health 16, no. 6: 916. https://doi.org/10.3390/ijerph16060916