Effect of Urban Wastewater Discharge on the Abundance of Antibiotic Resistance Genes and Antibiotic-Resistant Escherichia coli in Two Italian Rivers

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Water Sampling
2.2. Quantification of ARGs in Water Samples
2.3. Quantification of Total Heterotrophic Bacteria and E. coli
2.4. Phenotypic Antibiotic Resistance of E. Coli Isolates
2.5. Statistical Analyses
3. Results
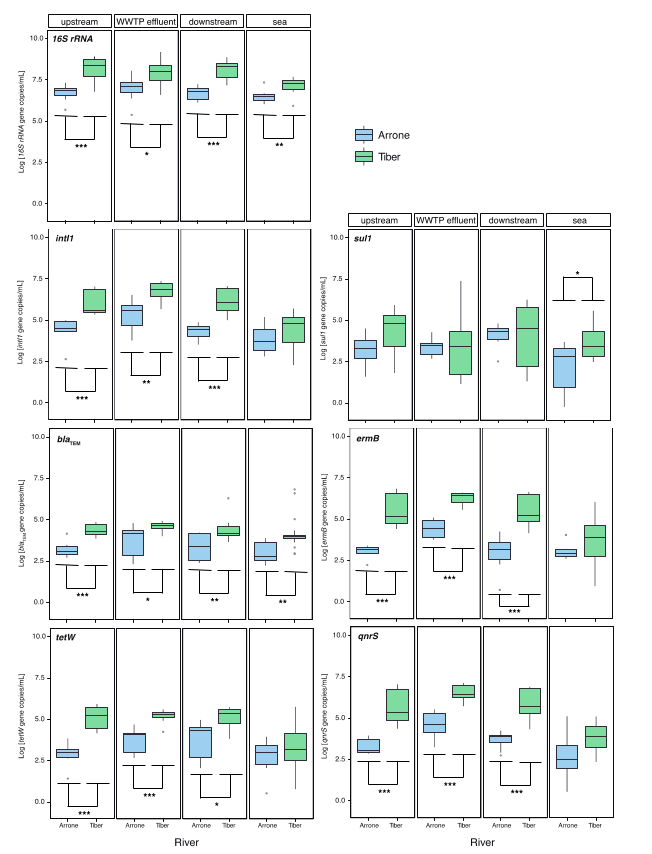
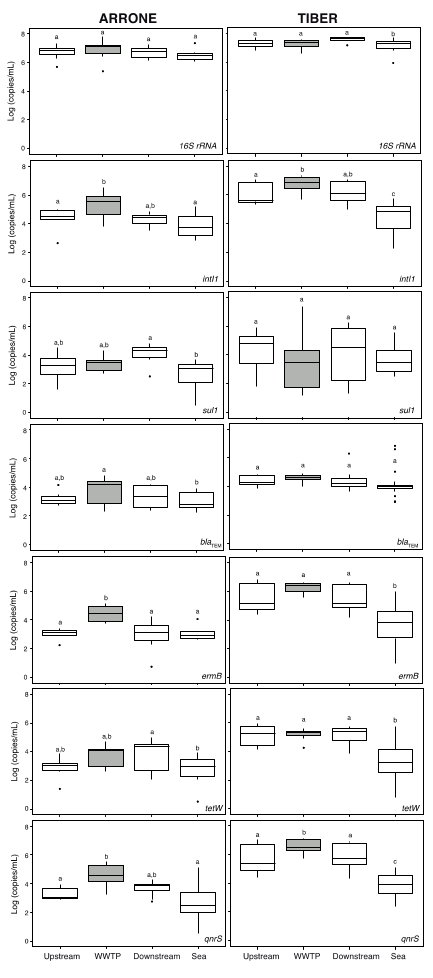
3.1. Concentration of Biomarker Genes and ARGs
3.2. Abundance of Escherichia Coli and Phenotypic Patterns of AR-E. Coli Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Windels, E.M.; Michiels, J.E.; Van den Bergh, B.; Fauvart, M.; Michiels, J. Antibiotics: Combatting Tolerance To Stop Resistance. MBio 2019, 10, e02095-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queenan, K.; Häsler, B.; Rushton, J. A One Health approach to antimicrobial resistance surveillance: Is there a business case for it? Int. J. Antimicrob. Agents 2016, 48, 422–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, D. A framework for improved one health governance and policy making for antimicrobial use. BMJ Glob. Health 2019, 4, e001807. [Google Scholar] [CrossRef] [Green Version]
- Lupo, A.; Coyne, S.; Berendonk, T.U. Origin and Evolution of Antibiotic Resistance: The Common Mechanisms of Emergence and Spread in Water Bodies. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Balcázar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I.; Mackie, R.I. Evolution and ecology of antibiotic resistance genes. FEMS Microbiol. Lett. 2007, 271, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Martínez, J.L.; Cantón, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2018, 42. [Google Scholar] [CrossRef]
- Manaia, C.M. Assessing the Risk of Antibiotic Resistance Transmission from the Environment to Humans: Non-Direct Proportionality between Abundance and Risk. Trends Microbiol. 2017, 25, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y.-G. Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Subirats, J.; Triadó-Margarit, X.; Mandaric, L.; Acuña, V.; Balcázar, J.L.; Sabater, S.; Borrego, C.M. Wastewater pollution differently affects the antibiotic resistance gene pool and biofilm bacterial communities across streambed compartments. Mol. Ecol. 2017, 26, 5567–5581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, E.; Balcázar, J.L. Real-Time PCR Assays for Quantification of qnr Genes in Environmental Water Samples and Chicken Feces. Appl. Environ. Microbiol. 2013, 79, 1743–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apat; Irsa-CNR. Metodi analitici per le acque; Manuali e Linee Guida; Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 2003; ISBN 88-448-0083-7. [Google Scholar]
- Eucast: The European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 7.1. Available online: www.eucast.org (accessed on 14 September 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2019. [Google Scholar]
- Grant, S.B.; Saphores, J.-D.; Feldman, D.L.; Hamilton, A.J.; Fletcher, T.D.; Cook, P.L.M.; Stewardson, M.; Sanders, B.F.; Levin, L.A.; Ambrose, R.F.; et al. Taking the “Waste” Out of “Wastewater” for Human Water Security and Ecosystem Sustainability. Science 2012, 337, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Gücker, B.; Brauns, M.; Solimini, A.G.; Voss, M.; Walz, N.; Pusch, M.T. Urban stressors alter the trophic basis of secondary production in an agricultural stream. Can. J. Fish. Aquat. Sci. 2011, 68, 74–88. [Google Scholar] [CrossRef]
- Aristi, I.; von Schiller, D.; Arroita, M.; Barceló, D.; Ponsatí, L.; García-Galán, M.J.; Sabater, S.; Elosegi, A.; Acuña, V. Mixed effects of effluents from a wastewater treatment plant on river ecosystem metabolism: Subsidy or stress? Freshw. Biol. 2015, 60, 1398–1410. [Google Scholar] [CrossRef]
- Proia, L.; Anzil, A.; Subirats, J.; Borrego, C.; Farrè, M.; Llorca, M.; Balcázar, J.L.; Servais, P. Antibiotic resistance along an urban river impacted by treated wastewaters. Sci. Total Environ. 2018, 628–629, 453–466. [Google Scholar] [CrossRef]
- Bréchet, C.; Plantin, J.; Sauget, M.; Thouverez, M.; Talon, D.; Cholley, P.; Guyeux, C.; Hocquet, D.; Bertrand, X. Wastewater Treatment Plants Release Large Amounts of Extended-Spectrum β-Lactamase–Producing Escherichia coli Into the Environment. Clin. Infect. Dis. 2014, 58, 1658–1665. [Google Scholar] [CrossRef]
- Pärnänen, K.M.M.; Narciso-da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Antibiotic resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 2019, 5, eaau9124. [Google Scholar] [CrossRef] [Green Version]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Vaz-Moreira, I.; Varela Della Giustina, S.; Llorca, M.; Barceló, D.; Schubert, S.; Berendonk, T.U.; Michael-Kordatou, I.; Fatta-Kassinos, D.; Martinez, J.L.; et al. Antibiotic residues in final effluents of European wastewater treatment plants and their impact on the aquatic environment. Environ. Int. 2020, 140, 105733. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Aubertheau, E.; Stalder, T.; Mondamert, L.; Ploy, M.-C.; Dagot, C.; Labanowski, J. Impact of wastewater treatment plant discharge on the contamination of river biofilms by pharmaceuticals and antibiotic resistance. Sci. Total Environ. 2017, 579, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.C.; Borowska, E.; Schwartz, T.; Horn, H. Impact of the particulate matter from wastewater discharge on the abundance of antibiotic resistance genes and facultative pathogenic bacteria in downstream river sediments. Sci. Total Environ. 2019, 649, 1171–1178. [Google Scholar] [CrossRef]
- Mahfouz, N.; Caucci, S.; Achatz, E.; Semmler, T.; Guenther, S.; Berendonk, T.U.; Schroeder, M. High genomic diversity of multi-drug resistant wastewater Escherichia coli. Sci. Rep. 2018, 8, 8928. [Google Scholar] [CrossRef]
- Suzuki, Y.; Hashimoto, R.; Xie, H.; Nishimura, E.; Nishiyama, M.; Nukazawa, K.; Ishii, S. Growth and antibiotic resistance acquisition of Escherichia coli in a river that receives treated sewage effluent. Sci. Total Environ. 2019, 690, 696–704. [Google Scholar] [CrossRef]
- Sykes, R.B.; Cimarusti, C.M.; Bonner, D.P.; Bush, K.; Floyd, D.M.; Georgopapadakou, N.H.; Koster, W.H.; Liu, W.C.; Parker, W.L.; Principe, P.A.; et al. Monocyclic β-lactam antibiotics produced by bacteria. Nature 1981, 291, 489–491. [Google Scholar] [CrossRef]
- Patterson, A.J.; Rincon, M.T.; Flint, H.J.; Scott, K.P. Mosaic Tetracycline Resistance Genes Are Widespread in Human and Animal Fecal Samples. Antimicrob. Agents Chemother. 2007, 51, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, G.; Sanseverino, I.; Della Libera, S.; Iaconelli, M.; Ferrero, V.E.V.; Barra Caracciolo, A.; Lettieri, T. The impact of anthropogenic pressure on the virological quality of water from the Tiber River, Italy. Lett. Appl. Microbiol. 2017, 65, 298–305. [Google Scholar] [CrossRef]
- Marcheggiani, S.; D’Ugo, E.; Puccinelli, C.; Giuseppetti, R.; D’Angelo, A.; Gualerzi, C.; Spurio, R.; Medlin, L.; Guillebault, D.; Baudart-Lenfant, J.; et al. Detection of Emerging and Re-Emerging Pathogens in Surface Waters Close to an Urban Area. Int. J. Environ. Res. Public Health 2015, 12, 5505–5527. Int. J. Environ. Res. Public Health 2015, 12, 13413–13414. [Google Scholar] [CrossRef] [Green Version]
- Patrolecco, L.; Capri, S.; Ademollo, N. Occurrence of selected pharmaceuticals in the principal sewage treatment plants in Rome (Italy) and in the receiving surface waters. Environ. Sci. Pollut. Res. 2015, 22, 5864–5876. [Google Scholar] [CrossRef] [PubMed]
- Bueno, I.; Williams-Nguyen, J.; Hwang, H.; Sargeant, J.M.; Nault, A.J.; Singer, R.S. Impact of point sources on antibiotic resistance genes in the natural environment: A systematic review of the evidence. Anim. Health Res. Rev. 2017, 18, 112–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy Chowdhury, P.; Ingold, A.; Vanegas, N.; Martínez, E.; Merlino, J.; Merkier, A.K.; Castro, M.; González Rocha, G.; Borthagaray, G.; Centrón, D.; et al. Dissemination of Multiple Drug Resistance Genes by Class 1 Integrons in Klebsiella pneumoniae Isolates from Four Countries: A Comparative Study. Antimicrob. Agents Chemother. 2011, 55, 3140–3149. [Google Scholar] [CrossRef] [Green Version]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.-C. Integron Involvement in Environmental Spread of Antibiotic Resistance. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Abella, J.; Fahy, A.; Duran, R.; Cagnon, C. Integron diversity in bacterial communities of freshwater sediments at different contamination levels. FEMS Microbiol. Ecol. 2015, 91, fiv140. [Google Scholar] [CrossRef] [Green Version]
- Ling, Z.; Yang, Y.; Huang, Y.; Zou, S.; Luan, T. A preliminary investigation on the occurrence and distribution of antibiotic resistance genes in the Beijiang River, South China. J. Environ. Sci. 2013, 25, 1656–1661. [Google Scholar] [CrossRef]
- Koczura, R.; Mokracka, J.; Taraszewska, A.; Łopacinska, N. Abundance of Class 1 Integron-Integrase and Sulfonamide Resistance Genes in River Water and Sediment Is Affected by Anthropogenic Pressure and Environmental Factors. Microb. Ecol. 2016, 72, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Sabri, N.A.; Schmitt, H.; Van der Zaan, B.; Gerritsen, H.W.; Zuidema, T.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. J. Environ. Chem. Eng. 2020, 8, 102245. [Google Scholar] [CrossRef]
- Leonard, A.F.C.; Zhang, L.; Balfour, A.J.; Garside, R.; Hawkey, P.M.; Murray, A.K.; Ukoumunne, O.C.; Gaze, W.H. Exposure to and colonisation by antibiotic-resistant E. coli in UK coastal water users: Environmental surveillance, exposure assessment, and epidemiological study (Beach Bum Survey). Environ. Int. 2018, 114, 326–333. [Google Scholar] [CrossRef]
- O’Flaherty, E.; Solimini, A.G.; Pantanella, F.; De Giusti, M.; Cummins, E. Human exposure to antibiotic resistant-Escherichia coli through irrigated lettuce. Environ. Int. 2019, 122, 270–280. [Google Scholar] [CrossRef]
- Na, G.; Lu, Z.; Gao, H.; Zhang, L.; Li, Q.; Li, R.; Yang, F.; Huo, C.; Yao, Z. The effect of environmental factors and migration dynamics on the prevalence of antibiotic-resistant Escherichia coli in estuary environments. Sci. Rep. 2018, 8, 1663. [Google Scholar] [CrossRef] [Green Version]
- Inghilesi, R.; Ottolenghi, L.; Orasi, A.; Pizzi, C.; Bignami, F.; Santoleri, R. Fate of river Tiber discharge investigated through numerical simulation and satellite monitoring. Ocean Sci. 2012, 8, 773–786. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Parameter | Arrone | Tiber | ||||||
|---|---|---|---|---|---|---|---|---|
| UP | WWTP | DW | SEA | UP | WWTP | DW | SEA | |
| E. coli mean (SD) | 3.4 (2.0) | 62.3 (7.7) | 21.9 (6.2) | 0.9 (9.9) | 102 (5.4) | 94.3 (3.3) | 120.8 (4.3) | 0.2 (3.3) |
| N° isolates | 40 | 36 | 53 | 33 | 57 | 31 | 51 | 42 |
| Tetracycline | 0.13 | 0.33 | 0.38 | 0.30 | 0.40 | 0.19 | 0.25 | 0.31 |
| Chloramphenicol | 0.08 | 0.11 | 0.11 | 0.12 | 0.14 | 0.23 | 0.16 | 0.12 |
| Ciprofloxacin | 0.03 | 0.33 | 0.21 | 0.12 | 0.16 | 0.10 | 0.06 | 0.05 |
| Trim/Sulfa | 0.15 | 0.14 | 0.21 | 0.30 | 0.26 | 0.19 | 0.10 | 0.07 |
| Amox/Clav | 0.10 | 0.11 | 0.09 | 0.12 | 0.11 | 0.13 | 0.18 | 0.05 |
| Imipenem | 0.03 | 0.08 | 0.08 | 0.00 | 0.04 | 0.03 | 0.04 | 0.00 |
| Cefotaxime | 0.00 | 0.08 | 0.11 | 0.00 | 0.07 | 0.03 | 0.04 | 0.00 |
| Amoxicillin | 0.10 | 0.19 | 0.19 | 0.30 | 0.37 | 0.32 | 0.29 | 0.36 |
| Gentamicin | 0.05 | 0.11 | 0.13 | 0.03 | 0.09 | 0.10 | 0.04 | 0.05 |
| Arrone | Tiber | |||||||
|---|---|---|---|---|---|---|---|---|
| Antibiotic | WWTP | DW | SEA | p-Value Site Effect | WWTP | DW | SEA | p-Value Site Effect |
| Tetracycline | 3.6 (1.1, 12.6) | 4.5 (1.6, 15) | 2.8 (0.8, 10) | 0.031 | 0.3 (0.1, 0.9) | 0.5 (0.2, 1.2) | 0.7 (0.3, 1.6) | 0.144 |
| Chloramphenicol | 1.4 (0.3, 7.6) | 1.6 (0.4, 8.3) | 2 (0.4, 11) | 0.846 | 1.9 (0.6, 5.9) | 1.1 (0.4, 3.3) | 0.8 (0.2, 2.6) | 0.579 |
| Ciprofloxacin | 19 (3.4, 358.6) | 10.7 (1.9, 201.4) | 6.7 (0.9, 136) | 0.002 | 0.5 (0.1, 2) | 0.3 (0.1, 1.2) | 0.3 (0, 1.2) | 0.226 |
| Trim/Sulfa | 0.9 (0.2, 3.2) | 1.5 (0.5, 4.7) | 2.7 (0.9, 9) | 0.234 | 0.7 (0.2, 1.9) | 0.3 (0.1, 0.9) | 0.2 (0, 0.7) | 0.026 |
| Amox/Clav | 1.1 (0.2, 5.2) | 0.9 (0.2, 4) | 1.3 (0.3, 6) | 0.976 | 1.4 (0.3, 5.5) | 1.8 (0.6, 5.8) | 0.4 (0.1, 2) | 0.278 |
| Imipenem | 3.3 (0.4, 68.2) | 3.3 (0.5, 65.8) | 0.1 (0.0,70.8) | 0.188 | 0.8 (0, 8.5) | 1.2 (0.1, 10.5) | 0.1 (0.0, 12.5) | 0.501 |
| Amoxicillin | 2.2 (0.6, 9) | 2.1 (0.6, 8.1) | 4.1 (1.2, 17) | 0.164 | 0.8 (0.3, 2.2) | 0.7 (0.3, 1.6) | 0.8 (0.3, 1.9) | 0.857 |
| Gentamicin | 2 (0.4, 15.7) | 3.3 (0.7, 23.6) | 0.7 (0, 8) | 0.305 | 1.2 (0.2, 5.6) | 0.4 (0.1, 2) | 0.4 (0, 1.9) | 0.435 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantanella, F.; Lekunberri, I.; Gagliardi, A.; Venuto, G.; Sànchez-Melsió, A.; Fabiani, M.; Balcázar, J.L.; Schippa, S.; De Giusti, M.; Borrego, C.; et al. Effect of Urban Wastewater Discharge on the Abundance of Antibiotic Resistance Genes and Antibiotic-Resistant Escherichia coli in Two Italian Rivers. Int. J. Environ. Res. Public Health 2020, 17, 6813. https://doi.org/10.3390/ijerph17186813
Pantanella F, Lekunberri I, Gagliardi A, Venuto G, Sànchez-Melsió A, Fabiani M, Balcázar JL, Schippa S, De Giusti M, Borrego C, et al. Effect of Urban Wastewater Discharge on the Abundance of Antibiotic Resistance Genes and Antibiotic-Resistant Escherichia coli in Two Italian Rivers. International Journal of Environmental Research and Public Health. 2020; 17(18):6813. https://doi.org/10.3390/ijerph17186813
Chicago/Turabian StylePantanella, Fabrizio, Itziar Lekunberri, Antonella Gagliardi, Giuseppe Venuto, Alexandre Sànchez-Melsió, Massimo Fabiani, José Luis Balcázar, Serena Schippa, Maria De Giusti, Carles Borrego, and et al. 2020. "Effect of Urban Wastewater Discharge on the Abundance of Antibiotic Resistance Genes and Antibiotic-Resistant Escherichia coli in Two Italian Rivers" International Journal of Environmental Research and Public Health 17, no. 18: 6813. https://doi.org/10.3390/ijerph17186813