A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production

 ,
,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Growth Substrate Preparation
2.2. Rearing of Hermetia Illucens Larvae
2.3. Laboratory and Apparatus
2.4. Chemical Analyses and Quality Control
2.5. Bioaccumulation Factor
2.6. Statistical Analysis
3. Results and Discussion
3.1. Potentially Toxic Elements Content in Growth Substrate Ingredients
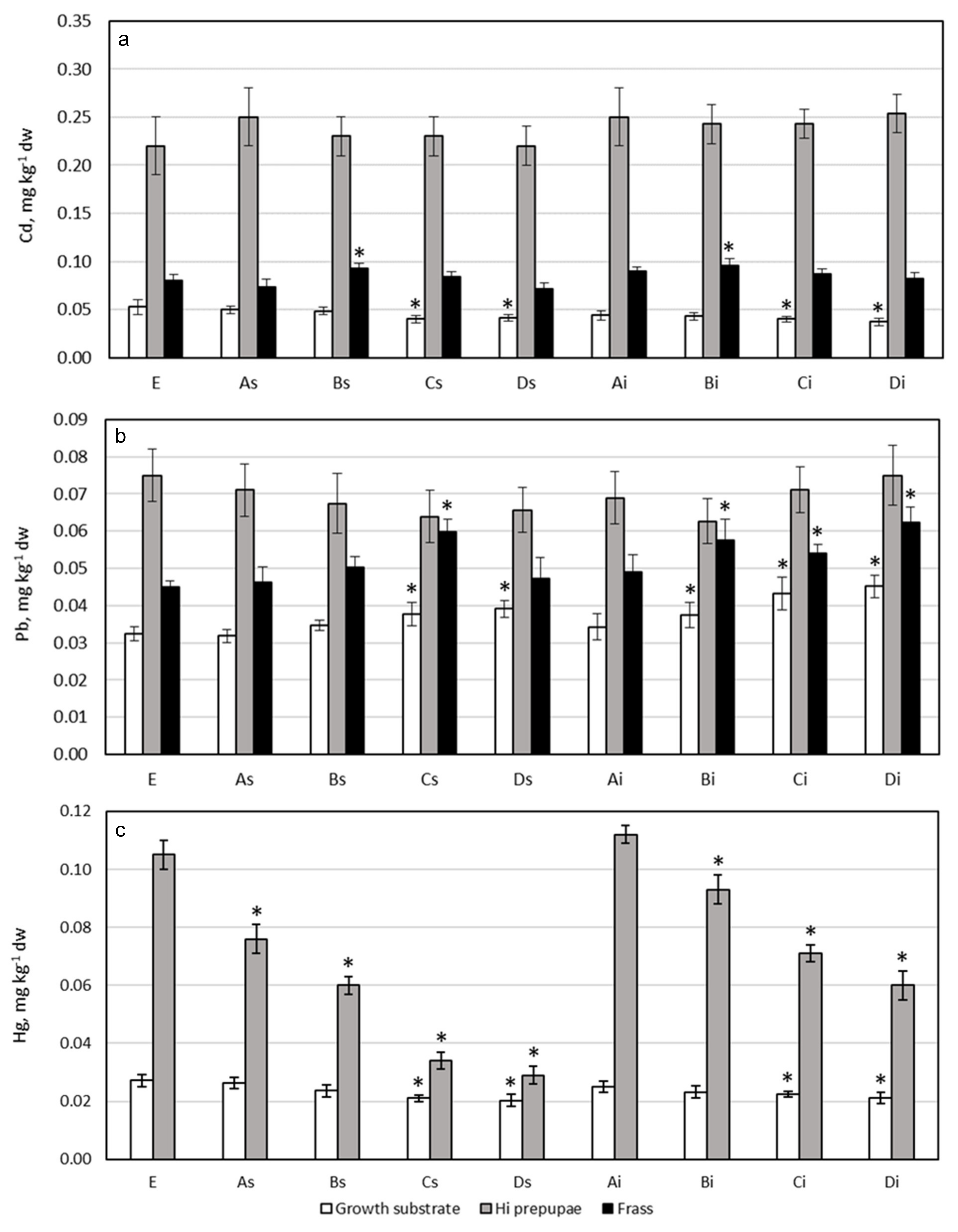
3.2. Cadmium
3.3. Lead
3.4. Mercury
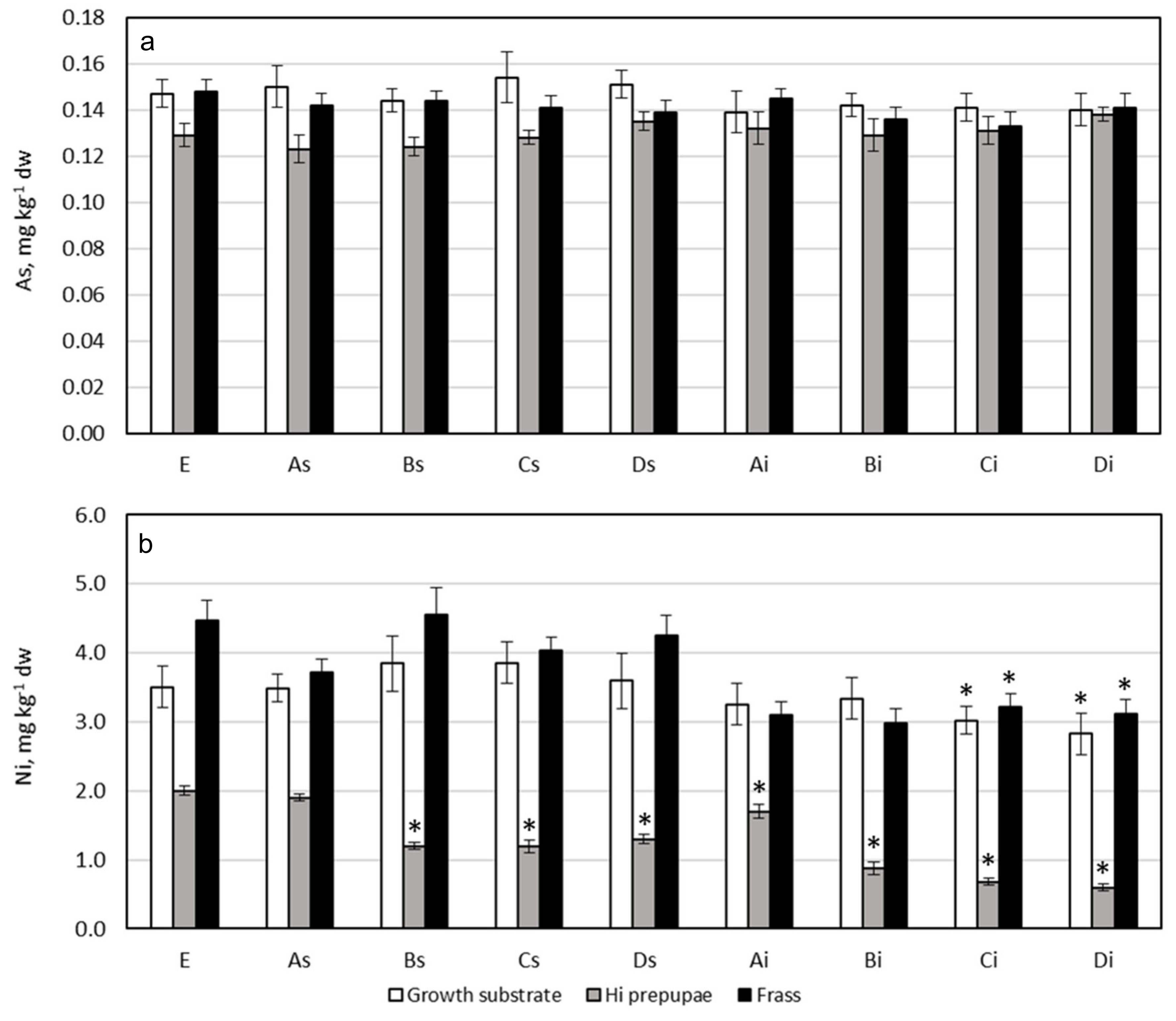
3.5. Arsenic
3.6. Nickel
3.7. Potentially Toxic Elements Content in HI Prepupae and Comparison with Legal Limit for Food
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bacchetti, T.; Annibaldi, A.; Comitini, F.; Ciani, M.; Damiani, E.; Norici, A.; Tiano, L.; Truzzi, C.; Olivotto, I. Alternative Ingredients for Feed and Food. In The First Outstanding 50 Years of Università Politecnica delle Marche; Longhi, S., Monteriù, A., Freddi, A., Frontoni, E., Germani, M., Revel, G.M., Eds.; Springer: Cham, Germany, 2020; ISBN 978-3-030-33831-2 978-3-030-33832-9. [Google Scholar] [CrossRef]
- da Silva Lucas, A.J.; de Oliveira, L.M.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive Compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef] [PubMed]
- van Raamsdonk, L.W.D.; van der Fels-Klerx, H.J.; de Jong, J. New feed ingredients: The insect opportunity. Food Addit. Contam: Part A 2017, 34, 1384–1397. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.; van Itterbeeck, J.; Heetkamp, M.J.; van den Brand, H.; van Loon, J.J.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- Costa-Neto, E.M. Insects as human food: An overview. Amaz. Rev. Antropol. 2013, 5, 562–582. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Scientific opinion on risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257–4317. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insect. Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Hall, R.D.; Gerhardt, R.R. Flies (Diptera). In Medical and Veterinary Entomology; Mullen, G., Durden, L., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 127–161. [Google Scholar]
- Sheppard, D.C.; Newton, G.L.; Thompson, S.A.; Savage, S. A value added manure management system using the black soldier fly. Bioresour. Technol. 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: stratiomyidae) larvae to recycle food waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef] [PubMed]
- Cutrignelli, M.I.; Messina, M.; Tulli, F.; Randazzo, B.; Olivotto, I.; Gasco, L.; Loponte, R.; Bovera, F. Evaluation of an insect meal of the black soldier fly (Hermetia illucens) as soybean substitute: Intestinal morphometry, enzymatic and microbial activity in laying hens. Res. Vet. Sci. 2018, 117, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial dietary inclusion of Hermetia illucens (black soldier fly) full-fat prepupae in zebrafish feed: Biometric, histological, biochemical, and molecular implications. Zebrafish 2018, 15, 519–553. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Delli Compagni, M.; Truzzi, C.; Riolo, P.; Loreto, N.; Ruschioni, S.; Osimani, A.; Giorgini, E.; et al. Black Soldier Fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518, 734659. [Google Scholar]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam: Part A 2017, 34, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metals toxicity and the environment. EXS 2012, 101, 133–164. [Google Scholar] [PubMed]
- Barbaro, E.; Zangrando, R.; Kirchgeorg, T.; Bazzano, A.; Illuminati, S.; Annibaldi, A.; Rella, S.; Truzzi, C.; Grotti, M.; Ceccarini, A.; et al. An integrated study of the chemical composition of Antarctic aerosol to investigate natural and anthropogenic sources. Environ. Chem. 2016, 125, 212–221. [Google Scholar] [CrossRef]
- Illuminati, S.; Annibaldi, A.; Truzzi, C.; Tercier-Waeber, M.L.; Nöel, N.; Braungardt, C.B.; Achterberg, E.P.; Howell, K.A.; Turner, D.; Marini, M.; et al. In-situ trace metal (Cd, Pb, Cu) speciation along the Po River plume (Northern Adriatic Sea) using submersible systems. Mar. Chem. 2019, 212, 47–63. [Google Scholar] [CrossRef]
- Truzzi, C.; Illuminati, S.; Girolametti, F.; Antonucci, M.; Scarponi, G.; Ruschioni, S.; Riolo, P.; Annibaldi, A. Influence of feeding substrates on the presence of toxic metals (Hg, Cd, Pb, Ni, As) in larvae of Tenebrio molitor: Risk assessment for human consumption. Int. J. Environ. Res. Public Health. 2019, 16, 4815. [Google Scholar] [CrossRef] [PubMed]
- Biancarosa, I.; Liland, N.S.; Biemans, D.; Araujo, P.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.J.; Amlund, H. Uptake of heavy metals and arsenic in black soldier fly (Hermetia illucens) larvae grown on seaweed-enriched media. J. Sci. Food Agric. 2018, 98, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- van der Fels-Klerx, H.J.; Camenzuli, L.; Belluco, S.; Meijer, N.; Ricci, A. Food safety issues related to uses of insects for feeds and foods. Compr. Rev. Food Sci. F. 2017, 17, 1172–1183. [Google Scholar] [CrossRef]
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Marone, P.A. Food Safety and Regulatory Concerns. In Insects as Sustainable Food Ingredients: Production, Processing and Food Applications, 1st ed.; Dossey, A.T., Morales-Ramos, J., Guadalupe Rojas, M., Eds.; Academic Press: Cambridge, UK, 2016. [Google Scholar]
- Zhang, Z.; Song, X.; Wang, Q.; Lu, X. Mercury bioaccumulation and prediction in terrestrial insects from soil in Huludao city, Northeast China. B Environ. Contam. Tox 2012, 89, 107–112. [Google Scholar] [CrossRef]
- Vijver, M.; Jager, T.; Posthuma, L.; Peijnenburg, W. Metal uptake from soils and soil–sediment mixtures by larvae of Tenebrio molitor (L.) (Coleoptera). Ecotoxicol. Environ. Safe 2003, 54, 277–289. [Google Scholar] [CrossRef]
- Charlton, A.J.; Dickinson, M.; Wakefield, M.E.; Fitches, E.; Kenis, M.; Han, R.; Zhu, F.; Kone, N.; Grant, M.; Devic, E.; et al. Exploring the chemical safety of fly larvae as a source of protein for animal feed. J. Insects Food Feed 2015, 1, 7–16. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Bioaccumulation of heavy metals in the black soldier fly, Hermetia illucens and effects on its life cycle. J. Insects Food Feed 2015, 1, 261–270. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, X.; Wang, W.; Lei, C.; Zhu, F. Influences of chromium and cadmium on the development of black soldier fly larvae. Environ. Sci. Pollut. Res. 2017, 24, 8637–8644. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; Camenzuli, L.; van der Lee, M.K.; Oonincx, D.G.A.B. Uptake of cadmium, lead and arsenic by Tenebrio molitor and Hermetia illucens from contaminated substrates. PLoS ONE 2016, 11, e0166186. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein; Eur-Lex: Brussels, Belgium, 2017. [Google Scholar]
- Truzzi, C.; Giorgini, E.; Annibaldi, A.; Antonucci, M.; Illuminati, S.; Scarponi, G.; Riolo, P.; Isidoro, N.; Conti, C.; Zarantoniello, M.; et al. Fatty acid composition and nutritional value of Hermetia illucens as novel ingredient for feed and food for the future: Influence of growth substrate based on coffee-waste silverskin and micro-algae. Anim. Feed Sci. Technol. 2020, 259, 114309. [Google Scholar] [CrossRef]
- European Union. Directive 2000/60/EC of the European Parliament and the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy; Eur-Lex: Brussels, Belgium, 2000. [Google Scholar]
- European Union. Directive 2006/1881 of the European Parliament and the Council of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs, and Amending Regulations No 420/2011 of 29 April 2011, and No 1006/2015 of 25 June 2015 as Regards Maximum Levels of Inorganic Arsenic in Foodstuffs; Eur-Lex: Brussels, Belgium, 2006. [Google Scholar]
- Van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: tenebrionidae) on diets composed of organic by-products. J. Insect. Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- May, B.M. The occurrence in New Zealand and the life-history of the soldier fly Hermetia illucens (L.) (Diptera: stratiomyidae). New Zeal. J. Sci. Wellingt. 1961, 4, 55–65. [Google Scholar]
- Illuminati, S.; Annibaldi, A.; Truzzi, C.; Scarponi, G. Recent Temporal Variations of Trace Metal Content in an Italian White Wine. Food Chem 2014, 159, 493–497. [Google Scholar] [CrossRef]
- Morgano, M.A.; Milani, R.F.; Perrone, A.A.M. Determination of total mercury in sushi samples employing direct mercury analyser. Food Anal. Method. 2015, 8, 2301–2307. [Google Scholar] [CrossRef]
- Annibaldi, A.; Truzzi, C.; Carnevali, O.; Pignalosa, P.; Api, M.; Scarponi, G.; Illuminati, S. Determination of Hg in farmed and wild atlantic bluefin tuna (Thunnus thynnus L.) muscle. Molecules 2019, 24, 1273. [Google Scholar] [CrossRef]
- Daniel, W.W.; Cross, C.L. Biostatistics: A Foundation for Analysis in the Health Sciences, 10th ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- STATGRAPHICS Centurion 18 software; Manugistics Inc.: Rockville, Maryland, 2018.
- Malik, J.; Szakova, J.; Drabek, O.; Balik, J.; Kokoska, L. Determination of certain micro and macroelements in plant stimulants and their infusions. Food Chem. 2008, 111, 520–525. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Chemical, functional, and structural properties of spent coffee grounds and coffee silverskin. Food Bioproc. Technol. 2014, 7, 3493–3503. [Google Scholar] [CrossRef]
- Jarošová, M.; Milde, D.; Kuba, M. Elemental analysis of coffee: A comparison of ICP-MS and AAS methods. Czech J. Food Sci. 2014, 32, 354–359. [Google Scholar] [CrossRef]
- Zarrinbakhsh, N.; Wang, T.; Rodriguez-Uribe, A.; Misra, M.; Mohanty, A.K. Characterization of wastes and coproducts from the coffee industry for composite material production. BioResources 2016, 11, 7637–7653. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Insect. Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- European Union. Directive 2002/32/EC of the European Parliament and the Council of 7 May 2002 on Undesirable Substances in Animal Feed; Eur-Lex: Brussels, Belgium, 2002. [Google Scholar]
- Hyun, S.H.; Kwon, K.H.; Park, K.H.; Jeong, H.C.; Kwon, O.; Tindwa, H.; Han, Y.S. Evaluation of nutritional status of an edible grasshopper. Oxya Chinensis Formosana. Entomol. Res. 2012, 42, 284–290. [Google Scholar] [CrossRef]
- Maryanski, M.; Kramarz, P.; Laskowski, R.; Niklinska, M. Decreased energetic reserves, morphological changes and accumulation of metals in carabid beetles (Poecilus cupreus L.) exposed to zinc-or cadmium-contaminated food. Ecotoxicol 2002, 11, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.X.; Ye, G.Y.; Hu, C.; Cheng, J.A. Accumulation of cadmium and its effects on growth, development and hemolymph biochemical compositions in Boettcherisca peregrina larvae (Diptera: Sarcophagidae). Insect. Sci. 2006, 13, 31–39. [Google Scholar] [CrossRef]
- Braeckman, B.; Smagghe, G.; Brutsaert, N.; Cornelis, R.; Raes, H. Cadmium uptake and defense mechanism in insect cells. Env. Res. 1999, 80, 231–243. [Google Scholar] [CrossRef]
- Payne, C.L.R.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Zhang, Z.S.; Lu, X.G.; Wang, Q.C.; Zheng, D.M. Mercury, cadmium and lead biogeochemistry in the soil–plant–insect system in Huludao city. Bull. Environ. Contam. Toxicol. 2009, 83, 255–259. [Google Scholar] [CrossRef]
- Sharma, R.K.; Agrawal, M.; Marshall, F. Heavy metal contamination of soil and vegetables in suburban areas of Varanasi, India. Ecotoxicol. Environ. Safety 2007, 66, 258–266. [Google Scholar] [CrossRef]
- Handley, M.A.; Hall, C.; Sanford, E.; Diaz, E.; GonzalezMendez, E.; Drace, K.; Wilson, R.; Villalobos, M.; Croughan, M. Globalization, binational communities, and imported food risks: Results of an outbreak investigation of lead poisoning in Monterey County, California. Amer. J. Public Health 2007, 97, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Zou, H.; Shu, W. Biotransfer of heavy metals along a soil-plant-insect-chicken food chain: Field study. J. Environ. Sci. 2009, 21, 849–853. [Google Scholar] [CrossRef]
- Köhler, R.; Kariuki, L.; Lambert, C.; Biesalski, H.K. Protein, amino acid and mineral composition of some edible insects from Thailand. J. Asia-Pac. Entomol 2019, 22, 372–378. [Google Scholar]
- Boudou, A.; Ribeyre, F. Metal ions in biological systems, vol. 34: “Mercury and its effects on environment and biology”. Met. Based Drugs 1997, 4, 287. [Google Scholar]
- Kidd, K.; Clayden, M.; Jardine, T. Bioaccumulation and biomagnification of mercury through food webs. In Environmental Chemistry and Toxicology of Mercury; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 455–499. [Google Scholar]
- Andrahennadi, R.; Pickering, I.J. Arsenic accumulation, biotransformation and localization in bertha armyworm moths. Environ. Chem. 2008, 5, 413–419. [Google Scholar] [CrossRef]
- Moriarty, M.M.; Koch, I.; Gordon, R.A.; Reimer, K.J. Arsenic speciation of terrestrial invertebrates. Environ. Sci. Technol. 2009, 43, 4818–4823. [Google Scholar] [CrossRef]
- Boyd, R.S. High-nickel insects and nickel hyperaccumulator plants: A review. Insect. Sci. 2009, 16, 19–31. [Google Scholar] [CrossRef]
- Sun, H.; Shu, Y.; Tang, W.; Wang, Q.; Zhou, Q.; Zhang, G.R. Nickel accumulation and its effects on the survival rate of Spodoptera litura fabricius under continuous nickel stress. Chim. Sci. Bull. 2007, 52, 1957–1963. [Google Scholar] [CrossRef]
- Poma, G.; Cuykx, M.; Amato, E.; Calaprice, C.; Focant, J.F.; Covaci, A. Evaluation of hazardous chemicals in edible insects and insect-based food intended for human consumption. Food Chem. Toxicol. 2017, 100, 70–79. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Element | Analytical Method | Analytical Result (n = 9) | Certified Value | Δ (%) |
|---|---|---|---|---|
| Cd | GF-AAS | 0.042 ± 0.005 | 0.043 ± 0.008 | −2 |
| Pb | GF-AAS | 0.068 ± 0.003 | 0.065 ± 0.007 | +5 |
| As | GF-AAS | 17.4 ± 0.6 | 18 ± 1.1 | −3 |
| Ni | GF-AAS | 18 ± 1.2 | 19.4 ± 3.1 | −7 |
| Hg | DMA-1 | 4.21 ± 0.06 | 4.58 ± 0.16 | −8 |
| Ingredients | Cd | Pb | Hg | As | Ni |
|---|---|---|---|---|---|
| Silverskin | 0.053 ± 0.008 b | 0.032 ± 0.002 a | 0.027 ± 0.001 c | 0.147 ± 0.006 a | 3.5 ± 0.2 b |
| Schizochytrium sp. | 0.0025 ± 0.0002 a | 0.065 ± 0.003 b | 0.009 ± 0.002 b | 0.184 ± 0.002 b | 3.6 ± 0.1 b |
| Isochrysis sp. | 0.0020 ± 0.0005 a | 0.084 ± 0.009 c | 0.0016 ± 0.0004 a | 0.153 ± 0.001 a | 1.18 ± 0.03 a |
| p-value | <0.0001 | 0.0002 | <0.0001 | <0.0001 | <0.0001 |
| HI Prepupae | Cd | Pb | Hg | As | Ni |
|---|---|---|---|---|---|
| HI E | 4.2 ± 0.9 | 2.3 ± 0.3 | 3.9 ± 0.2 | 0.88 ± 0.05 | 0.57 ± 0.05 |
| HI As | 5.0 ± 0.7 | 2.2 ± 0.3 | 2.9 ± 0.2 | 0.82 ± 0.05 | 0.54 ± 0.03 |
| HI Bs | 4.8 ± 0.5 | 1.9 ± 0.2 | 2.5 ± 0.1 | 0.86 ± 0.03 | 0.31 ± 0.03 |
| HI Cs | 5.7 ± 0.8 | 1.7 ± 0.2 | 1.6 ± 0.1 | 0.83 ± 0.06 | 0.31 ± 0.03 |
| HI Ds | 5.3 ± 0.7 | 1.7 ± 0.2 | 1.4 ± 0.1 | 0.89 ± 0.03 | 0.36 ± 0.04 |
| HI Ai | 5.7 ± 0.9 | 2.0 ± 0.3 | 4.5 ± 0.2 | 0.95 ± 0.08 | 0.52 ± 0.06 |
| HI Bi | 5.6 ± 0.7 | 1.7 ± 0.2 | 4.0 ± 0.2 | 0.91 ± 0.06 | 0.26 ± 0.04 |
| HI Ci | 6.1 ± 0.6 | 1.6 ± 0.2 | 3.2 ± 0.1 | 0.93 ± 0.06 | 0.23 ± 0.02 |
| HI Di | 6.9 ± 0.9 | 1.7 ± 0.2 | 2.8 ± 0.1 | 0.99 ± 0.05 | 0.21 ± 0.03 |
| HI Prepupae | Cd | Pb | Hg | As | Ni |
|---|---|---|---|---|---|
| Legal limit | 0.050-0.20 a (meat) | 0.10 a (meat) | 0.50 a (fish fillet) | 0.20 b (rice) | - |
| HI E | 0.076 ± 0.010 | 0.026 ± 0.002 | 0.030 ± 0.001 | 0.044 ± 0.002 | 0.76 ± 0.02 |
| HI As | 0.072 ± 0.009 | 0.021 ± 0.002 | 0.024 ± 0.002 | 0.036 ± 0.002 | 0.54 ± 0.01 |
| HI Bs | 0.072 ± 0.006 | 0.021 ± 0.002 | 0.020 ± 0.001 | 0.039 ± 0.001 | 0.36 ± 0.02 |
| HI Cs | 0.078 ± 0.007 | 0.022 ± 0.002 | 0.012 ± 0.001 | 0.043 ± 0.001 | 0.39 ± 0.03 |
| HI Ds | 0.076 ± 0.007 | 0.023 ± 0.002 | 0.010 ± 0.001 | 0.047 ± 0.001 | 0.46 ± 0.02 |
| HI Ai | 0.074 ± 0.005 | 0.020 ± 0.002 | 0.032 ± 0.003 | 0.037 ± 0.002 | 0.49 ± 0.03 |
| HI Bi | 0.076 ± 0.008 | 0.018 ± 0.002 | 0.029 ± 0.005 | 0.040 ± 0.002 | 0.27 ± 0.03 |
| HI Ci | 0.084 ± 0.006 | 0.020 ± 0.002 | 0.024 ± 0.003 | 0.045 ± 0.002 | 0.24 ± 0.02 |
| HI Di | 0.076 ± 0.006 | 0.021 ± 0.002 | 0.018 ± 0.005 | 0.041 ± 0.001 | 0.18 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truzzi, C.; Annibaldi, A.; Girolametti, F.; Giovannini, L.; Riolo, P.; Ruschioni, S.; Olivotto, I.; Illuminati, S. A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production. Int. J. Environ. Res. Public Health 2020, 17, 2121. https://doi.org/10.3390/ijerph17062121
Truzzi C, Annibaldi A, Girolametti F, Giovannini L, Riolo P, Ruschioni S, Olivotto I, Illuminati S. A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production. International Journal of Environmental Research and Public Health. 2020; 17(6):2121. https://doi.org/10.3390/ijerph17062121
Chicago/Turabian StyleTruzzi, Cristina, Anna Annibaldi, Federico Girolametti, Leonardo Giovannini, Paola Riolo, Sara Ruschioni, Ike Olivotto, and Silvia Illuminati. 2020. "A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production" International Journal of Environmental Research and Public Health 17, no. 6: 2121. https://doi.org/10.3390/ijerph17062121
APA StyleTruzzi, C., Annibaldi, A., Girolametti, F., Giovannini, L., Riolo, P., Ruschioni, S., Olivotto, I., & Illuminati, S. (2020). A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production. International Journal of Environmental Research and Public Health, 17(6), 2121. https://doi.org/10.3390/ijerph17062121