Effect of Temperature on the Survival of F-Specific RNA Coliphage, Feline Calicivirus, and Escherichia coli in Chlorinated Water

Abstract
:Introduction
Methods
Virus and Cell Culture
Coliphage
Bacteria
Experimental Plan
Data Analysis
Results and Discussions

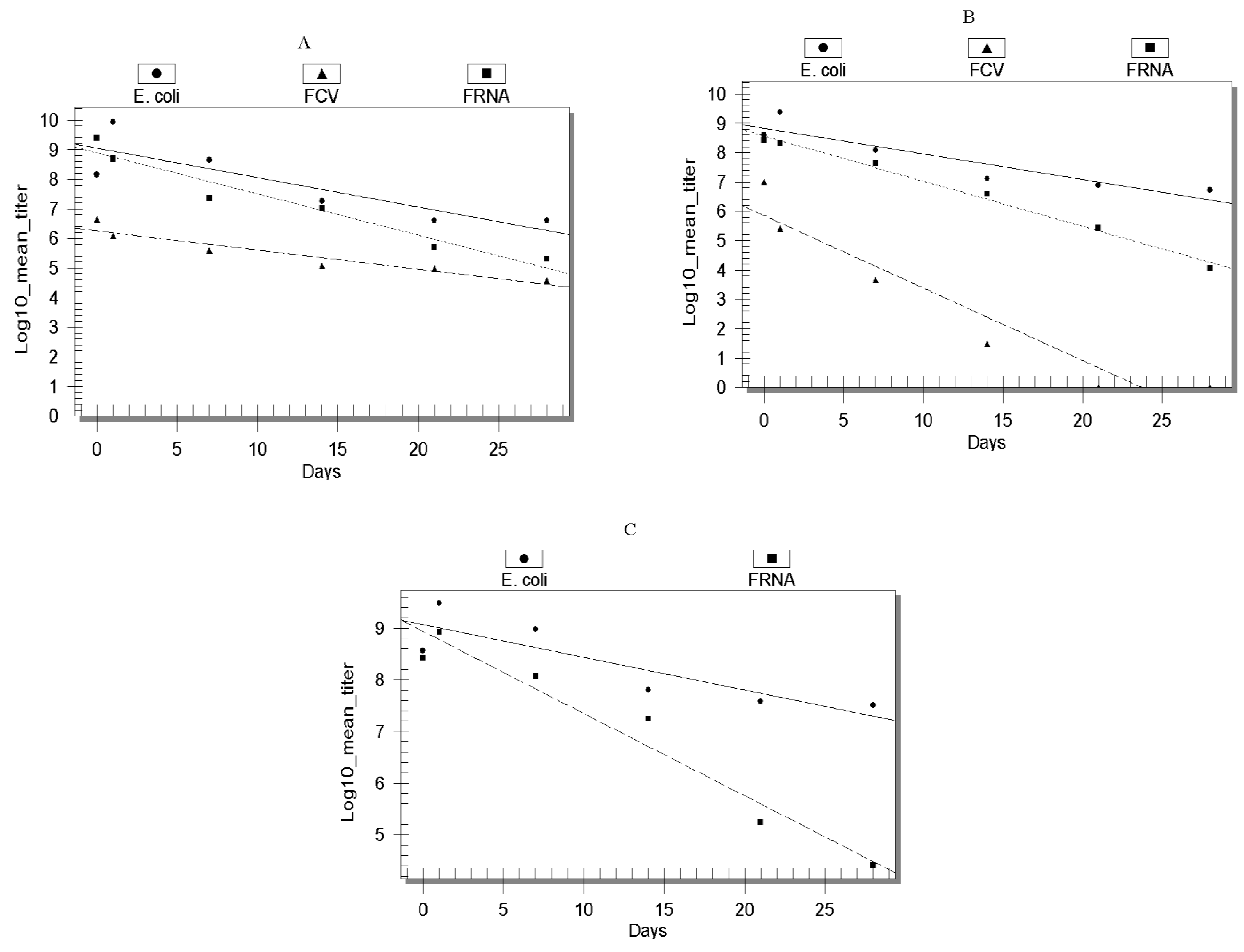{kind=link}
| Organism | Water Typea | Days to Detection Limit | ||
| 4°C | 25°C | 37°C | ||
| E. coli | Tap Water | >28 <1 | >28 | >28 |
| 50 ppm | <1 | <1 | ||
| Feline calicivirus | Tap Water | >28 | 14-21 | 1-7 |
| 50 ppm | <1 | <1 | <1 | |
| F-specific coliphage | Tap Water | >28 | >28 | >28 |
| 50 ppm | 7-14 | 7-14 | 7-14 | |
| Organism | Water Temperature | D-value (days) | r Squaredb |
| E. coli | 4°C | 10.1 | 0.73 |
| 25°C | 11.5 | 0.85 | |
| 37°C | 15.9 | 0.75 | |
| FCVc | 4°C | 15.6 | 0.89 |
| 25°C | 4.0 | 0.91 | |
| 37°C | N/Ad | N/A | |
| F-specific coliphage | 4°C | 7.2 | 0.94 |
| 25°C | 6.5 | 0.99 | |
| 37°C | 6.2 | 0.95 |

References
- Anderson, A.D.; Heryford, A.G.; Sarisky, J.P.; Higgins, C.; Monroe, S.S.; Beard, R.S.; Newport, C.M.; Cashdollar, J.L.; Fout, G.S.; Robbins, D.E.; Seys, S.A.; Musgrave, K.J.; Medus, C.; Vinje, J.; Bresee, J.S.; Mainzer, H.M.; Glass, R.I. A waterborne outbreak of Norwalk-like virus among snowmobilers-Wyoming, 2001. J. Infect. Dis. 2003, 187(2), 303–306. [Google Scholar]
- Pedalino, B.; Feely, E.; McKeown, P.; Foley, B.; Smyth, B.; Moren, A. An outbreak of Norwalk-like viral gastroenteritis in holidaymakers travelling to Andorra, January-February 2002. Euro Surveill 2003, 8(1), 1–8. [Google Scholar]
- Lopman, B.A.; Reacher, M.H.; Van Duijnhoven, Y.; Hanon, F.X.; Brown, D.; Koopmans, M. Viral gastroenteritis outbreaks in Europe, 1995-2000. Emerg. Infect. Dis. 2003, 9(1), 90–96. [Google Scholar]
- Griffin, D.W.; Donaldson, K.A.; Paul, J.H.; Rose, J.B. Pathogenic human viruses in coastal waters. Clin. Microbiol. Rev. 2003, 16(1), 129–143. [Google Scholar]
- Fleet, G.H.; Heiskanen, P.; Reid, I.; Buckle, K.A. Foodborne viral illness--status in Australia. Int. J. Food Microbiol. 2000, 59(1-2), 127–136. [Google Scholar]
- Grabow, W.O.; Taylor, M.B.; de Villiers, J.C. New methods for the detection of viruses: call for review of drinking water quality guidelines. Water Sci. Technol. 2001, 43(12), 1–8. [Google Scholar]
- Tree, J.A.; Adams, M.R.; Lees, D.N. Chlorination of indicator bacteria and viruses in primary sewage effluent. Appl. Environ. Microbiol. 2003, 69(4), 2038–2043. [Google Scholar] [CrossRef]
- Boccia, D.; Tozzi, A.E.; Cotter, B.; Rizzo, C.; Russo, T.; Buttinelli, G.; Caprioli, A.; Marziano, M.L.; Ruggeri, F.M. Waterborne outbreak of Norwalk-like virus gastroenteritis at a tourist resort, Italy. Emerg. Infect. Dis. 2002, 8(6), 563–568. [Google Scholar]
- Leclerc, H.; Schwartzbrod, L.; Dei-Cas, E. Microbial agents associated with waterborne diseases. Crit. Rev. Microbiol. 2002, 28(4), 371–409. [Google Scholar]
- Gofti-Laroche, L.; Gratacap-Cavallier, B.; Demanse, D.; Genoulaz, O.; Seigneurin, J.M.; Zmirou, D. Are waterborne astrovirus implicated in acute digestive morbidity (E.MI.R.A. study)? J. Clin. Virol. 2003, 27(1), 74–82. [Google Scholar]
- Carducci, A.; Casini, B.; Bani, A.; Rovini, E.; Verani, M.; Mazzoni, F.; Giuntini, A. Virological control of groundwater quality using biomolecular tests. Water Sci. Technol. 2003, 47(3), 261–266. [Google Scholar]
- Borchardt, M.A.; Bertz, P.D.; Spencer, S.K.; Battigelli, D.A. Incidence of enteric viruses in groundwater from household wells in Wisconsin. Appl. Environ. Microbiol. 2003, 69(2), 1172–1180. [Google Scholar]
- Jacangelo, J.G.; Loughran, P.; Petrik, B.; Simpson, D.; McIlroy, C. Removal of enteric viruses and selected microbial indicators by UV irradiation of secondary effluent. Water Sci. Technol. 2003, 47(9), 193–198. [Google Scholar]
- Formiga-Cruz, M.; Allard, A.K.; Conden-Hansson, A.C.; Henshilwood, K.; Hernroth, B.E.; Jofre, J.; Lees, D.N.; Lucena, F.; Papapetropoulou, M.; Rangdale, R.E.; Tsibouxi, A.; Vantarakis, A.; Girones, R. Evaluation of potential indicators of viral contamination in shellfish and their applicability to diverse geographical areas. Appl. Environ. Microbiol. 2003, 69(3), 1556–1563. [Google Scholar]
- Brion, G.M.; Meschke, J.S.; Sobsey, M.D. F-specific RNA coliphages: occurrence, types, and survival in natural waters. Water Res. 2002, 36(9), 2419–2425. [Google Scholar]
- Havelaar, A.H.; van Olphen, M.; Drost, Y.C. F-specific RNA bacteriophages are adequate model organisms for enteric viruses in fresh water. Appl. Environ. Microbiol. 1993, 59(9), 2956–2962. [Google Scholar]
- Dore, W.J.; Henshilwood, K.; Lees, D.N. Evaluation of F-specific RNA bacteriophage as a candidate human enteric virus indicator for bivalve molluscan shellfish. Appl. Environ. Microbiol. 2000, 66(4), 1280–1285. [Google Scholar]
- Schaper, M.; Jofre, J.; Uys, M.; Grabow, W.O. Distribution of genotypes of F-specific RNA bacteriophages in human and non-human sources of faecal pollution in South Africa and Spain. J. Appl. Microbiol. 2002, 92(4), 657–667. [Google Scholar] [CrossRef]
- Thurston-Enriquez, J.A.; Haas, C.N.; Jacangelo, J.; Riley, K.; Gerba, C.P. Inactivation of feline calicivirus and adenovirus type 40 by UV radiation. Appl. Environ. Microbiol. 2003, 69(1), 557–582. [Google Scholar]
- Slomka, M.J.; Appleton, H. Feline calicivirus as a model system for heat inactivation studies of small round structured viruses in shellfish. Epidemiol. Infect. 1998, 121(2), 401–407. [Google Scholar] [CrossRef]
- Gulati, B.R.; Allwood, P.B.; Hedberg, C.W.; Goyal, S.M. Efficacy of commonly used disinfectants for the inactivation of calicivirus on strawberry, lettuce, and a food-contact surface. J. Food Prot. 2001, 64(9), 1430–1434. [Google Scholar]
- Doultree, J.C.; Druce, J.D.; Birch, C.J.; Bowden, D.S.; Marshall, J.A. Inactivation of feline calicivirus, a Norwalk virus surrogate. J. Hosp. Infect. 1999, 41(1), 51–57. [Google Scholar]
- Taku, A.; Gulati, B.R.; Allwood, P.B.; Palazzi, K.; Hedberg, C.W.; Goyal, S.M. Concentration and detection of caliciviruses from food contact surfaces. J. Food Prot. 2002, 65(6), 999–1004. [Google Scholar]
- Allwood, P.B.; Malik, Y.S.; Hedberg, C.W.; Goyal, S.M. Survival of F-specific RNA coliphage, feline calicivirus, and Escherichia coli in water: a comparative study. Appl. Environ. Microbiol. 2003, 69(9), 5707–5710. [Google Scholar]
- Reed, L. J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- USEPA. Method 1601: male-specific (F+) and somatic coliphage in water by two-step enrichment procedure. Available online: http://www.epa.gov/microbes/1601ap01.pdf (Accessed December 10, 2004).
- Jackson, R.W.; Osborne, K.; Barnes, G.; Jolliff, C.; Zamani, D.; Roll, B.; Stillings, A.; Herzog, D.; Cannon, S.; Loveland, S. Multiregional evaluation of the SimPlate heterotrophic plate count method compared to the standard plate count agar pour plate method in water. Appl. Environ. Microbiol. 2000, 66(1), 453–454. [Google Scholar]
- Sutton, S.V.; Franco, R.J.; Porter, D.A.; Mowrey-McKee, M.F.; Busschaert, S.C.; Hamberger, J.F.; Proud, D.W. D-value determinations are an inappropriate measure of disinfecting activity of common contact lens disinfecting solutions. Appl. Environ. Microbiol. 1999, 57(7), 2021–2026. [Google Scholar]
- World Health Organization. The right to water. Available online: http://www.who.int/water_sanitation_health/Documents/righttowater/righttowater.htm (Accessed June 24, 2005).
- World Health Organization. Water resource quality. Available online: http://www.who.int/water_sanitation_health/resourcesquality/en/ (Accessed October 10, 2003).
- Karim, M.R.; Pontius, F.W.; LeChevallier, M.W. Detection of noroviruses in water--summary of an international workshop. J. Infect. Dis. 2004, 189(1), 21–28. [Google Scholar]
- Cheesbrough, J.S.; Barkess-Jones, L.; Brown, D.W. Possible prolonged environmental survival of small round structured viruses. J. Hosp. Infect. 1997, 35(4), 325–326. [Google Scholar] [CrossRef]
- Cheesbrough, J.S.; Green, J.; Gallimore, C.I.; Wright, P.A.; Brown, D.W. Widespread environmental contamination with Norwalk-like viruses (NLV) detected in a prolonged hotel outbreak of gastroenteritis. Epidemiol. Infect. 2000, 125(1), 93–98. [Google Scholar]
- Hewitt, J.; Greening, G.E. Survival and persistence of norovirus, hepatitis A virus, and feline calicivirus in marinated mussels. J. Food Prot. 2004, 67(8), 1743–1750. [Google Scholar]
- Duizer, E.; Bijkerk, P.; Rockx, B.; De Groot, A.; Twisk, F.; Koopmans, M. Inactivation of caliciviruses. Appl. Environ. Microbiol. 2004, 70(8), 4538–454. [Google Scholar]
- Bidawid, S.; Malik, N.; Adegbunrin, O.; Sattar, S.A.; Farber, J.M. Norovirus cross-contamination during food handling and interruption of virus transfer by hand antisepsis: experiments with feline calicivirus as a surrogate. J. Food Prot. 2004, 67(1), 103–109. [Google Scholar]
- Gehrke, C.; Steinmann, J.; Goroncy-Bermes, P. Inactivation of feline calicivirus, a surrogate of norovirus (formerly Norwalk-like viruses), by different types of alcohol in vitro and in vivo. J. Hosp. Infect. 2004, 56(1), 49–55. [Google Scholar]
- Kadoi, K.; Kadoi, B. K. Stability of feline caliciviruses in marine water maintained at different temperatures. New Microbiol. 2001, 24(1), 17–21. [Google Scholar]
- Kingsley, D.H.; Hoover, D.G.; Papafragkou, E.; Richards, G.P. Inactivation of hepatitis A virus and a calicivirus by high hydrostatic pressure. J. Food Prot. 2002, 65(10), 1605–1609. [Google Scholar]
- Lee, K.M.; Gillespie, J.H. Thermal and pH stability of feline calicivirus. Infect. Immun. 1973, 7(4), 678–679. [Google Scholar]
- Nuanualsuwan, S.; Mariam, T.; Himathongkham, S.; Cliver, D.O. Ultraviolet inactivation of feline calicivirus, human enteric viruses and coliphages. Photochem. Photobiol. 2002, 76(4), 406–410. [Google Scholar]
- USEPA. Combined sewer overflow technology fact sheet: Chlorine disinfection; EPA 832-F-99-034: Washington, DC, 1999. [Google Scholar]
© 2005 MDPI. All rights reserved.
Share and Cite
Allwood, P.B.; Malik, Y.S.; Maherchandani, S.; Hedberg, C.W.; Goyal, S.M. Effect of Temperature on the Survival of F-Specific RNA Coliphage, Feline Calicivirus, and Escherichia coli in Chlorinated Water. Int. J. Environ. Res. Public Health 2005, 2, 442-446. https://doi.org/10.3390/ijerph2005030008
Allwood PB, Malik YS, Maherchandani S, Hedberg CW, Goyal SM. Effect of Temperature on the Survival of F-Specific RNA Coliphage, Feline Calicivirus, and Escherichia coli in Chlorinated Water. International Journal of Environmental Research and Public Health. 2005; 2(3):442-446. https://doi.org/10.3390/ijerph2005030008
Chicago/Turabian StyleAllwood, Paul B., Yashpal S. Malik, Sunil Maherchandani, Craig W. Hedberg, and Sagar M. Goyal. 2005. "Effect of Temperature on the Survival of F-Specific RNA Coliphage, Feline Calicivirus, and Escherichia coli in Chlorinated Water" International Journal of Environmental Research and Public Health 2, no. 3: 442-446. https://doi.org/10.3390/ijerph2005030008
APA StyleAllwood, P. B., Malik, Y. S., Maherchandani, S., Hedberg, C. W., & Goyal, S. M. (2005). Effect of Temperature on the Survival of F-Specific RNA Coliphage, Feline Calicivirus, and Escherichia coli in Chlorinated Water. International Journal of Environmental Research and Public Health, 2(3), 442-446. https://doi.org/10.3390/ijerph2005030008