
Estimating Field-Level Perennial Bioenergy Grass Biomass Yields Using the Normalized Difference Red-Edge Index and Linear Regression Analysis for Central Virginia, USA

Abstract
:1. Introduction
1.1. Use of Remote Sensing for Estimating Perennial Bioenergy Grass Biomass Yields
1.2. Relevance of the NDRE for Estimating Perennial Bioenergy Grass Biomass Yields
2. Materials and Methods
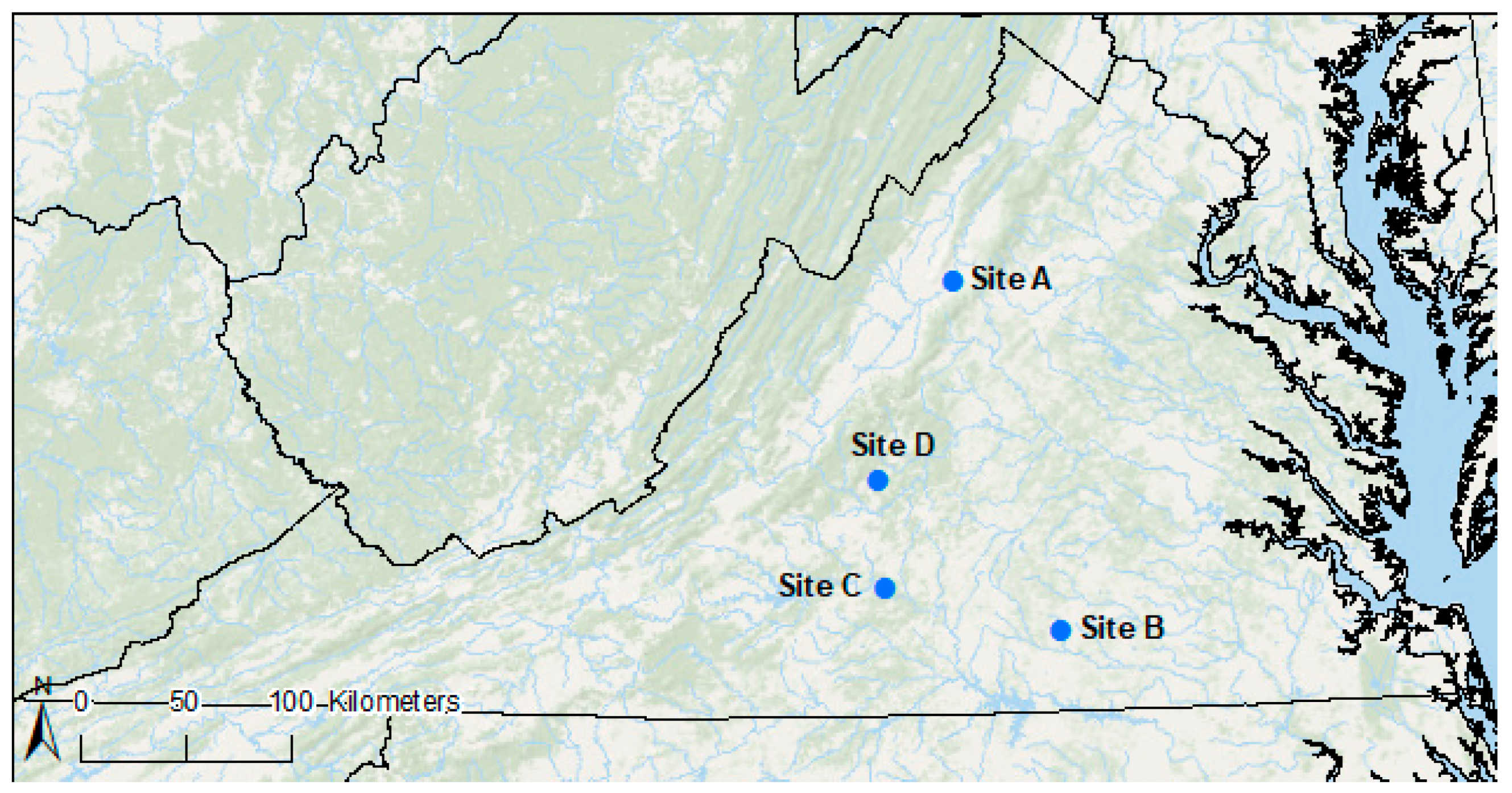
2.1. Study Area
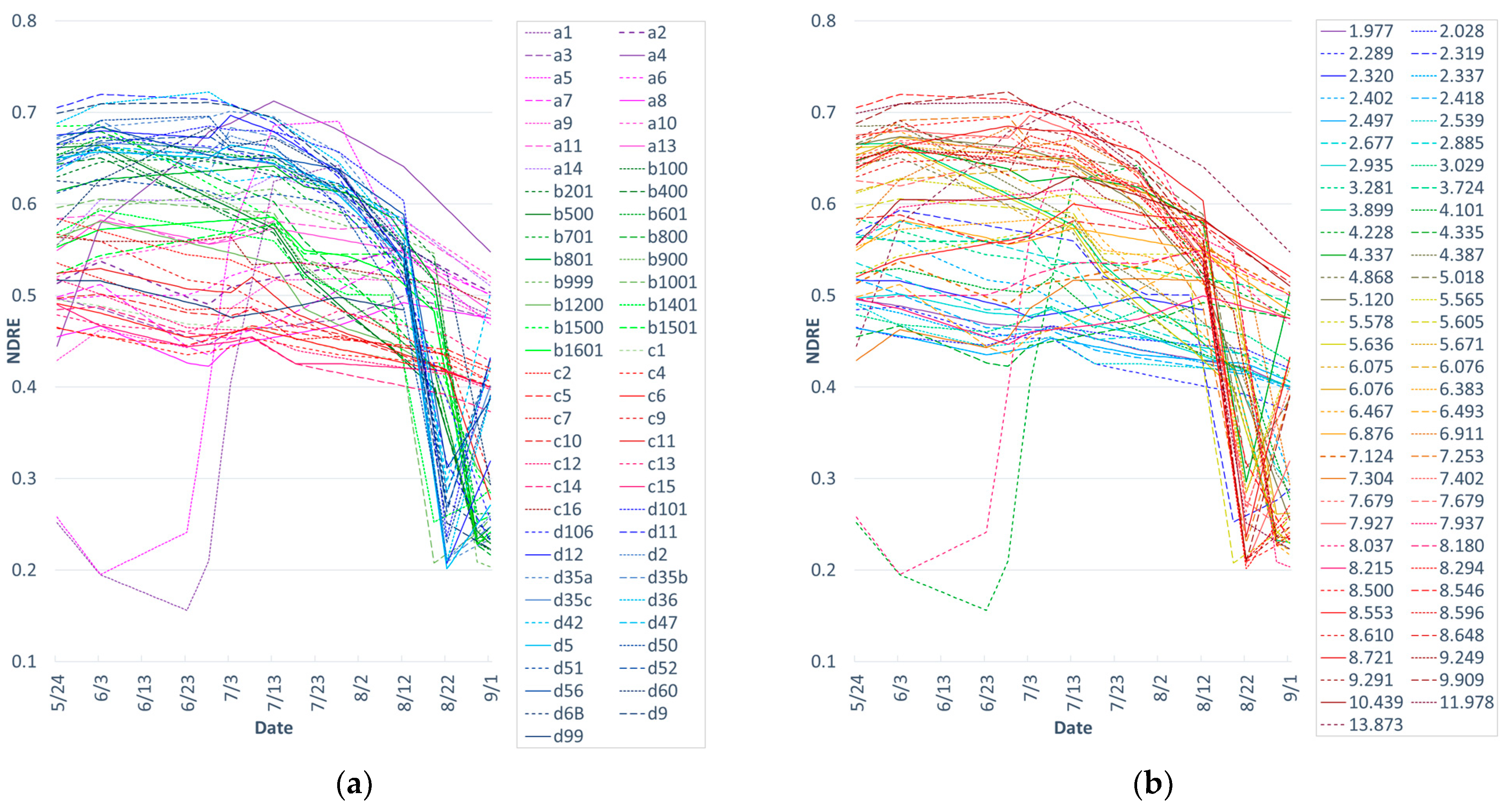
2.2. Data
2.3. Analysis
3. Results
3.1. Seasonal Trajectory of the NDRE
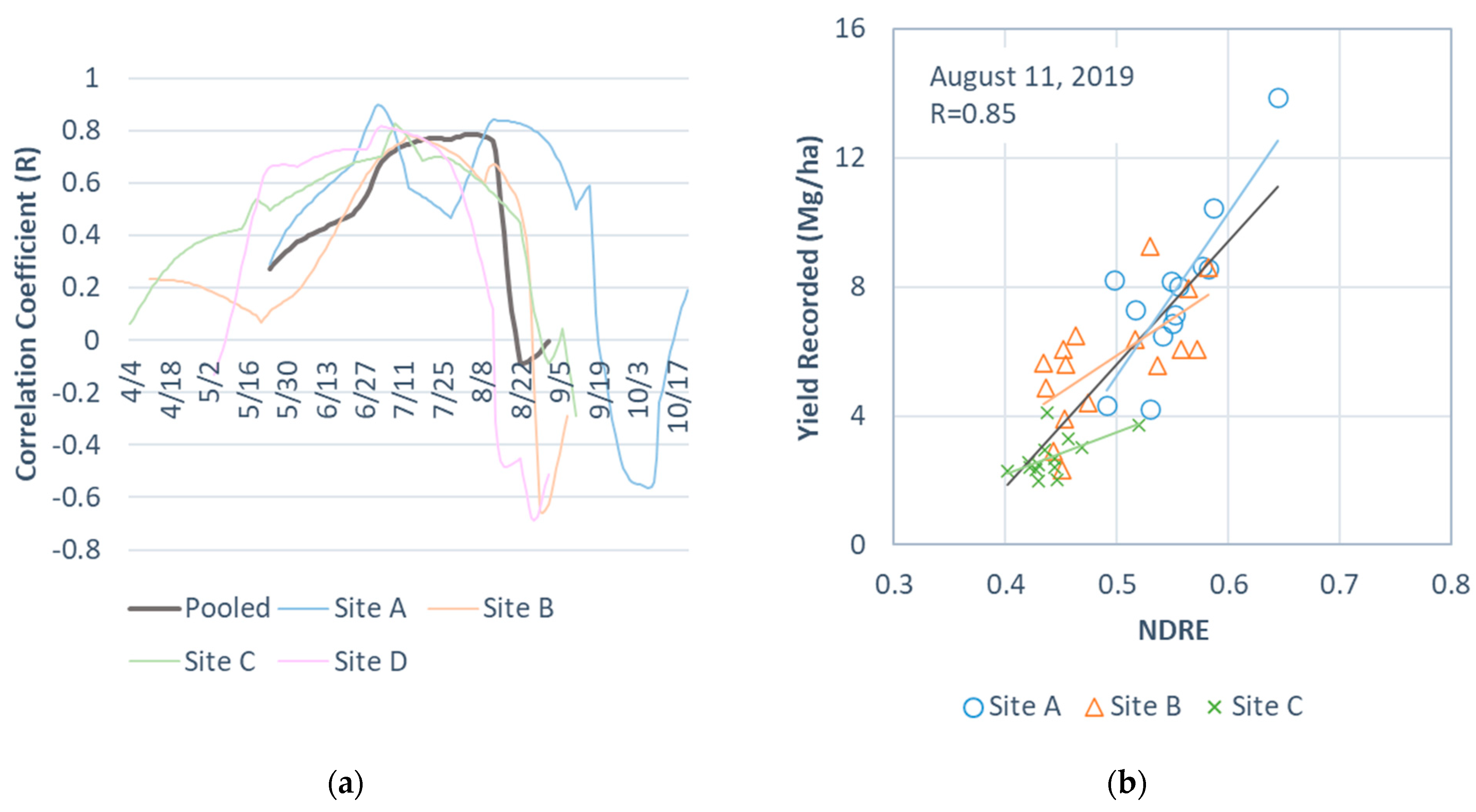
3.2. Optimal Date of NDRE for Estimating Perennial Bioenergy Grass Biomass Yields
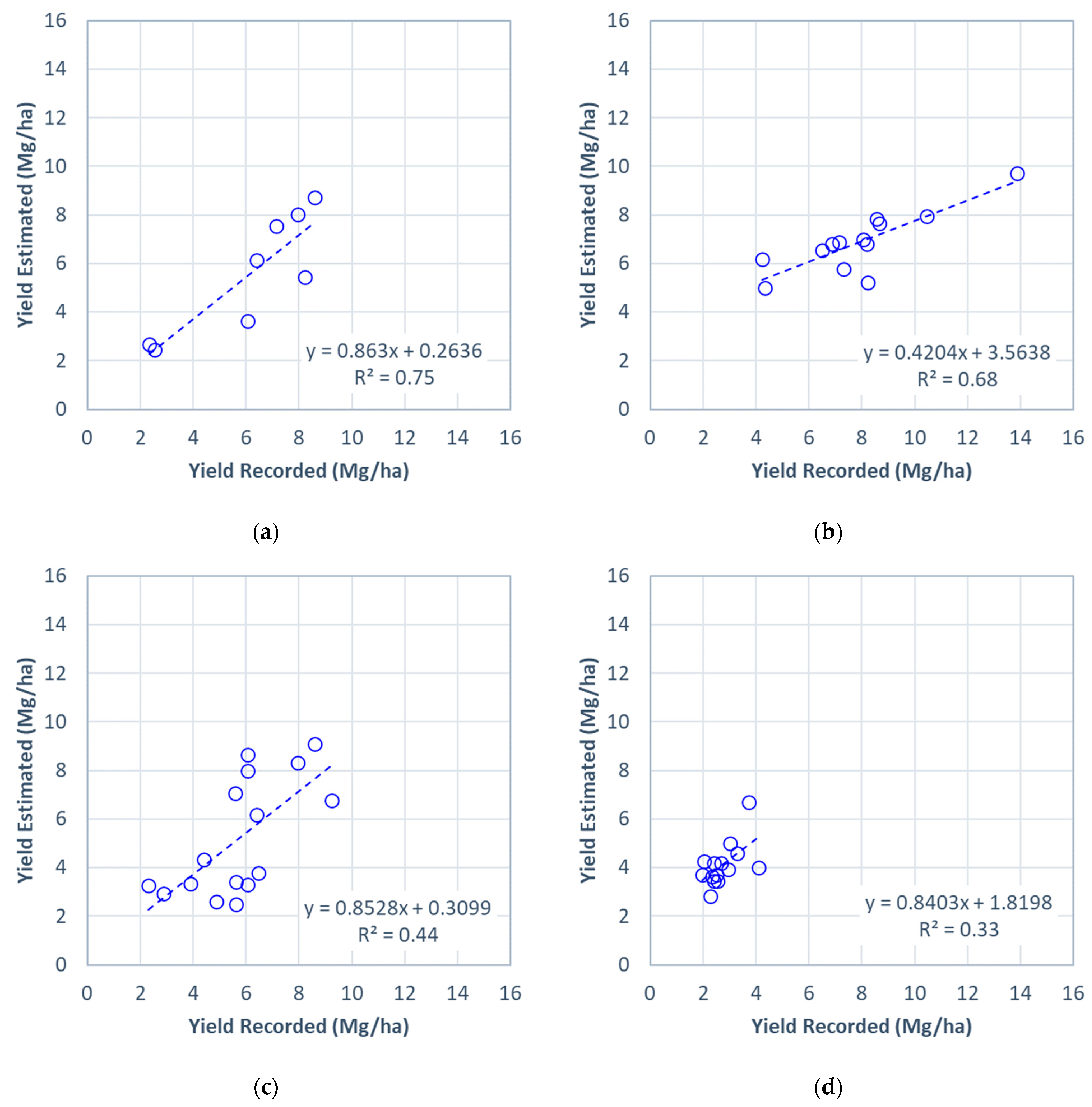
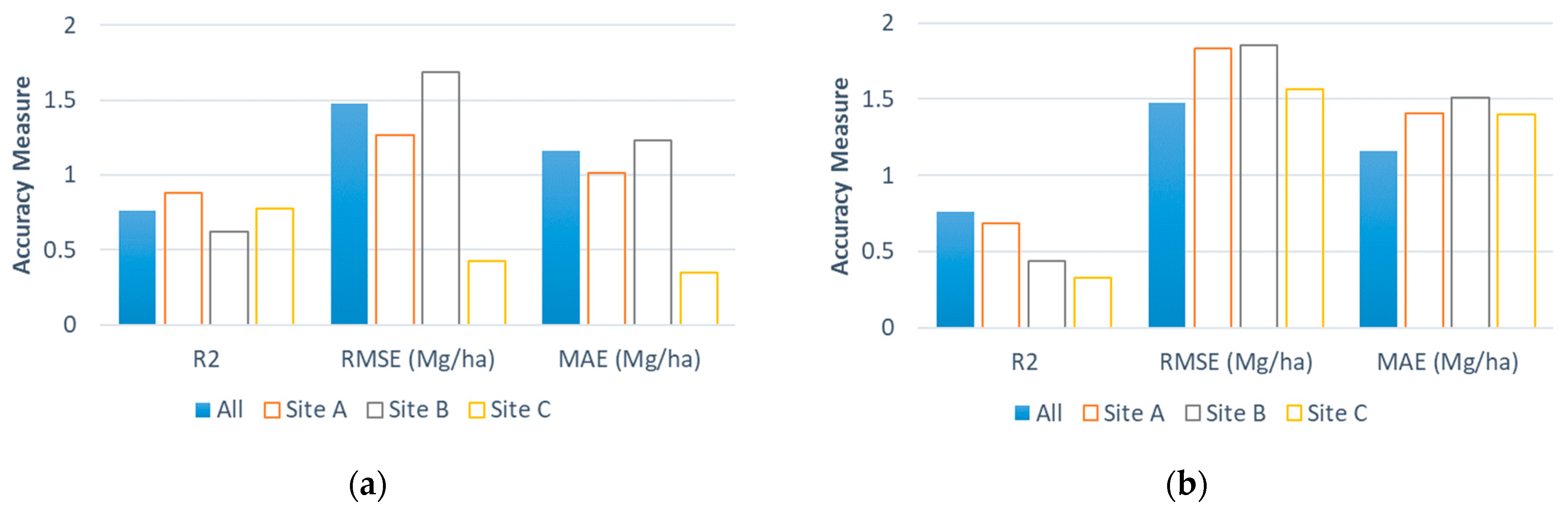
3.3. Accuracy of Estimated Field-Level Perennial Bioenergy Grass Biomass Yields Using a Linear NDRE Regression Model
3.4. Statistical Significance of the NDRE for Estimating the Perennial Bioenergy Grass Biomass Yields Using a Linear Regression Model
3.5. Bias and Consistency in Estimating the Field-Level Perennial Bioenergy Grass Biomass Yields Using a Linear NDRE Regression Model
4. Discussion
4.1. Impact of the Harvest Date on Determining a Common Optimal Date for Multiple Sites
4.2. Possible Transition of the NDRE Indication from the Early to the Late Growing Season
4.3. Challenge in Estimating Biomass Yields for Sites with Hetergeneous Perennial Bioenergy Grass Production
4.4. Limitations Associated with Inconsistent Yield Ranges across the Study Sites
5. Conclusions
- Inconsistent with the existing studies [24,25,32], the optimal date for estimating field-level perennial bioenergy grass biomass yields for the central Virginia sites using the pooled data was in late summer (11 August). We hypothesized that the index–biomass relationships specific to a perennial bioenergy grass type are the strongest in early summer to mid-summer [24,32] and become more indicative of the overall biomass than the biomass specific to a certain grass type as crops mature and senesce later in summer. This could be an important consideration for scaling the perennial bioenergy grass biomass yield estimation for a large number of sites. Examining relationships between the recorded yields and index values by perennial bioenergy grass types from early summer to right before harvest would allow the testing of this hypothesis.
- We should expect an earlier harvest for sites with a short growing season or for growing perennial grasses with earlier peak biomass yields than other sites. The optimal date may overlap with, or occur after, harvest for those sites. The sites with early harvest may require a separate model because of their potentially distinct yield–index relationships. This would be a critical limitation for scaling the current method for estimating perennial bioenergy grass biomass yields over large areas.
- Regardless of the approach (i.e., all-site, cross-site, or site-specific estimations), estimating the biomass yields for a site with heterogeneous perennial bioenergy grasses is more difficult than for sites with a monoculture or uniform mixed grass for the fields within a site. Stratifying the fields by grass types would allow type-specific index–yield relationships to be compared for fields with comparable crop types. This comparison could determine the optimal date for yield estimation by grass type. It could also identify grasses that may hinder the remotely sensed biomass yield estimation using a single model due to their unique spectral-response characteristics.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L.; et al. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.A.; Niemi, J.; Helmers, M.J.; Liebman, M.; Arbuckle, J.G.; James, D.E.; Kolka, R.K.; O’neal, M.E.; Tomer, M.D.; Tyndall, J.C.; et al. Prairie strips improve biodiversity and the delivery of multiple ecosystem services from corn–soybean croplands. Proc. Natl. Acad. Sci. USA 2017, 114, 11247–11252. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.S.; Blanco-Canqui, H. Lignocellulosic-based bioenergy and water quality parameters: A review. GCB Bioenergy 2018, 10, 504–533. [Google Scholar] [CrossRef]
- Wang, E.; Cruse, R.M.; Sharma-Acharya, B.; Herzmann, D.E.; Gelder, B.K.; James, D.E.; Flanagan, D.C.; Blanco-Canqui, H.; Mitchell, R.B.; Laird, D.A. Strategic switchgrass (Panicum virgatum) production within row cropping systems: Regional-scale assessment of soil erosion loss and water runoff impacts. GCB Bioenergy 2020, 12, 955–967. [Google Scholar] [CrossRef]
- Zaimes, G.N.; Schultz, R.C. Assessing Riparian Conservation Land Management Practice Impacts on Gully Erosion in Iowa. Environ. Manag. 2012, 49, 1009–1021. [Google Scholar] [CrossRef]
- Yimam, Y.T.; Ochsner, T.E.; Fox, G.A. Hydrologic cost-effectiveness ratio favors switchgrass production on marginal croplands over existing grasslands. PLoS ONE 2017, 12, e0181924. [Google Scholar] [CrossRef]
- Blanco-Canqui, H. Energy Crops and Their Implications on Soil and Environment. Agron. J. 2010, 102, 403–419. [Google Scholar] [CrossRef]
- Zhou, X.; Helmers, M.J.; Asbjornsen, H.; Kolka, R.; Tomer, M.D.; Cruse, R.M. Nutrient removal by prairie filter strips in agricultural landscapes. J. Soil Water Conserv. 2014, 69, 54–64. [Google Scholar] [CrossRef]
- de Freitas, E.N.; Khatri, V.; Contin, D.R.; de Oliveira, T.B.; Contato, A.G.; Peralta, R.M.; dos Santos, W.D.; Martinez, C.A.; Saddler, J.N.; Polizeli, M.d.L.T. Climate change affects cell-wall structure and hydrolytic performance of a perennial grass as an energy crop. Biofuels Bioprod. Biorefining 2022, 16, 471–487. [Google Scholar] [CrossRef]
- Jager, H.I.; Hilliard, M.R.; Langholtz, M.H.; Efroymson, R.A.; Brandt, C.C.; Nair, S.S.; Kreig, J.A. Ecosystem service benefits to water users from perennial biomass production. Sci. Total. Environ. 2022, 834, 155255. [Google Scholar] [CrossRef]
- Brandes, E.; McNunn, G.S.; A Schulte, L.; Bonner, I.J.; Muth, D.J.; A Babcock, B.; Sharma, B.; A Heaton, E. Subfield profitability analysis reveals an economic case for cropland diversification. Environ. Res. Lett. 2016, 11, 014009. [Google Scholar] [CrossRef]
- Ssegane, H.; Negri, M.C.; Quinn, J.; Urgun-Demirtas, M. Multifunctional landscapes: Site characterization and field-scale design to incorporate biomass production into an agricultural system. Biomass-Bioenergy 2015, 80, 179–190. [Google Scholar] [CrossRef]
- Ssegane, H.; Negri, M.C. An Integrated Landscape Designed for Commodity and Bioenergy Crops for a Tile-Drained Agricultural Watershed. J. Environ. Qual. 2016, 45, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, T.; Tian, L.; Zhang, Y.; Ting, K. A review of remote sensing methods for biomass feedstock production. Biomass-Bioenergy 2011, 35, 2455–2469. [Google Scholar] [CrossRef]
- Schmer, M.R.; Mitchell, R.B.; Vogel, K.P.; Schacht, W.H.; Marx, D.B. Spatial and Temporal Effects on Switchgrass Stands and Yield in the Great Plains. BioEnergy Res. 2010, 3, 159–171. [Google Scholar] [CrossRef]
- Cacho, J.F.; Feinstein, J.; Zumpf, C.R.; Hamada, Y.; Lee, D.J.; Namoi, N.L.; Lee, D.; Boersma, N.N.; Heaton, E.A.; Quinn, J.J.; et al. Predicting Biomass Yields of Advanced Switchgrass Cultivars for Bioenergy and Ecosystem Services Using Machine Learning. Energies 2023, 16, 416. [Google Scholar] [CrossRef]
- Chen, Y.; Guerschman, J.; Shendryk, Y.; Henry, D.; Harrison, M.T. Estimating Pasture Biomass Using Sentinel-2 Imagery and Machine Learning. Remote Sens. 2021, 13, 603. [Google Scholar] [CrossRef]
- Reinermann, S.; Asam, S.; Kuenzer, C. Remote Sensing of Grassland Production and Management—A Review. Remote Sens. 2020, 12, 1949. [Google Scholar] [CrossRef]
- Ahmad, A.; Gilani, H.; Ahmad, S.R. Forest Aboveground Biomass Estimation and Mapping through High-Resolution Optical Satellite Imagery—A Literature Review. Forests 2021, 12, 914. [Google Scholar] [CrossRef]
- Chao, Z.; Liu, N.; Zhang, P.; Ying, T.; Song, K. Estimation methods developing with remote sensing information for energy crop biomass: A comparative review. Biomass-Bioenergy 2019, 122, 414–425. [Google Scholar] [CrossRef]
- Giovos, R.; Tassopoulos, D.; Kalivas, D.; Lougkos, N.; Priovolou, A. Remote Sensing Vegetation Indices in Viticulture: A Critical Review. Agriculture 2021, 11, 457. [Google Scholar] [CrossRef]
- Jensen, J.R. Remote Sensing of the Environment: An Earth Resource Perspective; Prentice Hall: Upper Saddle River, NJ, USA, 2003. [Google Scholar]
- Kanemasu, E.T.; Demetriades-Shah, T.H.; Su, H.; Lang, A.R.G. Estimating Grassland Biomass Using Remotely Sensed Data. Appl. Remote Sens. Agric. 1990, 185–199. [Google Scholar] [CrossRef]
- Hamada, Y.; Zumpf, C.R.; Cacho, J.F.; Lee, D.; Lin, C.-H.; Boe, A.; Heaton, E.; Mitchell, R.; Negri, M.C. Remote Sensing-Based Estimation of Advanced Perennial Grass Biomass Yields for Bioenergy. Land 2021, 10, 1221. [Google Scholar] [CrossRef]
- Namoi, N.; Jang, C.; Robins, Z.; Lin, C.-H.; Lim, S.-H.; Voigt, T.; Lee, D. Aerial Imagery Can Detect Nitrogen Fertilizer Effects on Biomass and Stand Health of Miscanthus × giganteus. Remote Sens. 2022, 14, 1435. [Google Scholar] [CrossRef]
- Foster, A.J.; Kakani, V.G.; Mosali, J. Estimation of bioenergy crop yield and N status by hyperspectral canopy reflectance and partial least square regression. Precis. Agric. 2016, 18, 192–209. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.; Schjoerring, J. Reflectance measurement of canopy biomass and nitrogen status in wheat crops using normalized difference vegetation indices and partial least squares regression. Remote Sens. Environ. 2003, 86, 542–553. [Google Scholar] [CrossRef]
- Naguib, N.S.; Daliman, S. Analysis of NDVI and NDRE Indices Using Satellite Images for Crop Identification at Kelantan. IOP Conf. Ser. Earth Environ. Sci. 2022, 1102, 012054. [Google Scholar] [CrossRef]
- Chang, J.; Shoshany, M. Red-edge ratio normalized vegetation index for remote estimation of green biomass. In Proceedings of the 2016 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Beijing, China, 10–15 July 2016; pp. 1337–1339. [Google Scholar] [CrossRef]
- Hamada, Y.; Zumpf, C.R.; Quinn, J.J. Predicting Switchgrass Biomass Yields Using a Spectral Vegetation Index Derived from Multispectral Satellite Imagery; Technical Report, ANL/EVS-23/20; Argonne National Laboratory (ANL): Argonne, IL, USA, 2023; 44p. [Google Scholar]
- Datt, B. A New Reflectance Index for Remote Sensing of Chlorophyll Content in Higher Plants: Tests using Eucalyptus Leaves. J. Plant Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S.A. Comparison of time series tasseled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]
- Klemas, V.; Smart, R. The Influence of Soil Salinity, Growth Form and Leaf Moisture on the Spectral Radiance of Spartina alterniflora Canopies. Photogramm. Eng. Remote Sens. 1983, 49, 77–83. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site A | Site B | Site C | Site D | |
|---|---|---|---|---|
| Elevation (m) | 305 | 116 | 168 | 244 |
| Total field count | 13 | 18 | 15 | 24 |
| Field count available for modeling | 13 | 16 | 14 | 20 |
| Total area 1 (ha) | 243.4 | 134.3 | 63.8 | 100.2 |
| Average field size (ha) [standard deviation] | 18.7 [20.6] | 8.4 [7.9] | 4.3 [3.4] | 5.0 [2.6] |
| Average field-level perennial bioenergy grass yield 2 (Mg/ha) [standard deviation] | 7.9 [2.4] | 5.7 [1.8] | 2.7 [0.6] | 7.2 [2.0] |
| Perennial bioenergy grass variety | Switchgrass ‘Alamo’ | Switchgrass ‘Kanlow’, Miscanthus, mixed grass (big bluestem, Indiangrass, switchgrass) | Mixed grass (big bluestem, Indiangrass, switchgrass) | Switchgrass ‘Blackwell’ |
| Harvest activities for the 2019 season | ||||
| Mowing period | 9/18–10/6 | 8/13–8/29 | 9/3–9/5 | 7/30–8/4 |
| Raking period | 9/19–10/7 | 8/25–8/31 | 9/4–9/6 | 7/30–8/12 |
| Baling begins | 9/19–10/7 | 8/27–8/31 | 9/4–9/6 | 8/4–8/21 |
| Dates of Sentinel-2 image availability for the 2019 season | ||||
| April | — | 11, 16 | 4 | 4 |
| May | 24 | 19, 21, 24 | 14, 19, 24 | 4, 19, 24 |
| June | 3, 23, 28 | 3, 15 | 3, 23 | 3, 13, 28 |
| July | 3, 13, 28 | 10, 13, 20, 30 | 3, 8, 18, 28 | 3, 13, 28 |
| August | 12 | 9, 19, 29 | 22 | 12, 22 |
| September | 11, 16, 21 | 8, 21 | 1, 6, 11, 16, 21 | 1, 11, 16, 21, 26 |
| October | 1, 11, 21 | 1, 18, 28 | 1, 11 | 1, 11 |
| Total images considered | 14 | 19 | 18 | 19 |
| Site | R2 | RMSE (Mg/ha) | MAE (Mg/ha) |
|---|---|---|---|
| All | 0.76 | 1.5 | 1.2 |
| A | 0.69 | 1.8 | 1.4 |
| B | 0.44 | 1.9 | 1.5 |
| C | 0.33 | 1.6 | 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, Y.; Zumpf, C.R.; Quinn, J.J.; Negri, M.C. Estimating Field-Level Perennial Bioenergy Grass Biomass Yields Using the Normalized Difference Red-Edge Index and Linear Regression Analysis for Central Virginia, USA. Energies 2023, 16, 7397. https://doi.org/10.3390/en16217397
Hamada Y, Zumpf CR, Quinn JJ, Negri MC. Estimating Field-Level Perennial Bioenergy Grass Biomass Yields Using the Normalized Difference Red-Edge Index and Linear Regression Analysis for Central Virginia, USA. Energies. 2023; 16(21):7397. https://doi.org/10.3390/en16217397
Chicago/Turabian StyleHamada, Yuki, Colleen R. Zumpf, John J. Quinn, and Maria Cristina Negri. 2023. "Estimating Field-Level Perennial Bioenergy Grass Biomass Yields Using the Normalized Difference Red-Edge Index and Linear Regression Analysis for Central Virginia, USA" Energies 16, no. 21: 7397. https://doi.org/10.3390/en16217397