Comparative Histopathology of Host Reaction Types in Slash Pine Resistant to Cronartium quercuum f. sp. fusiforme

Abstract
:1. Introduction
2. Experimental Section
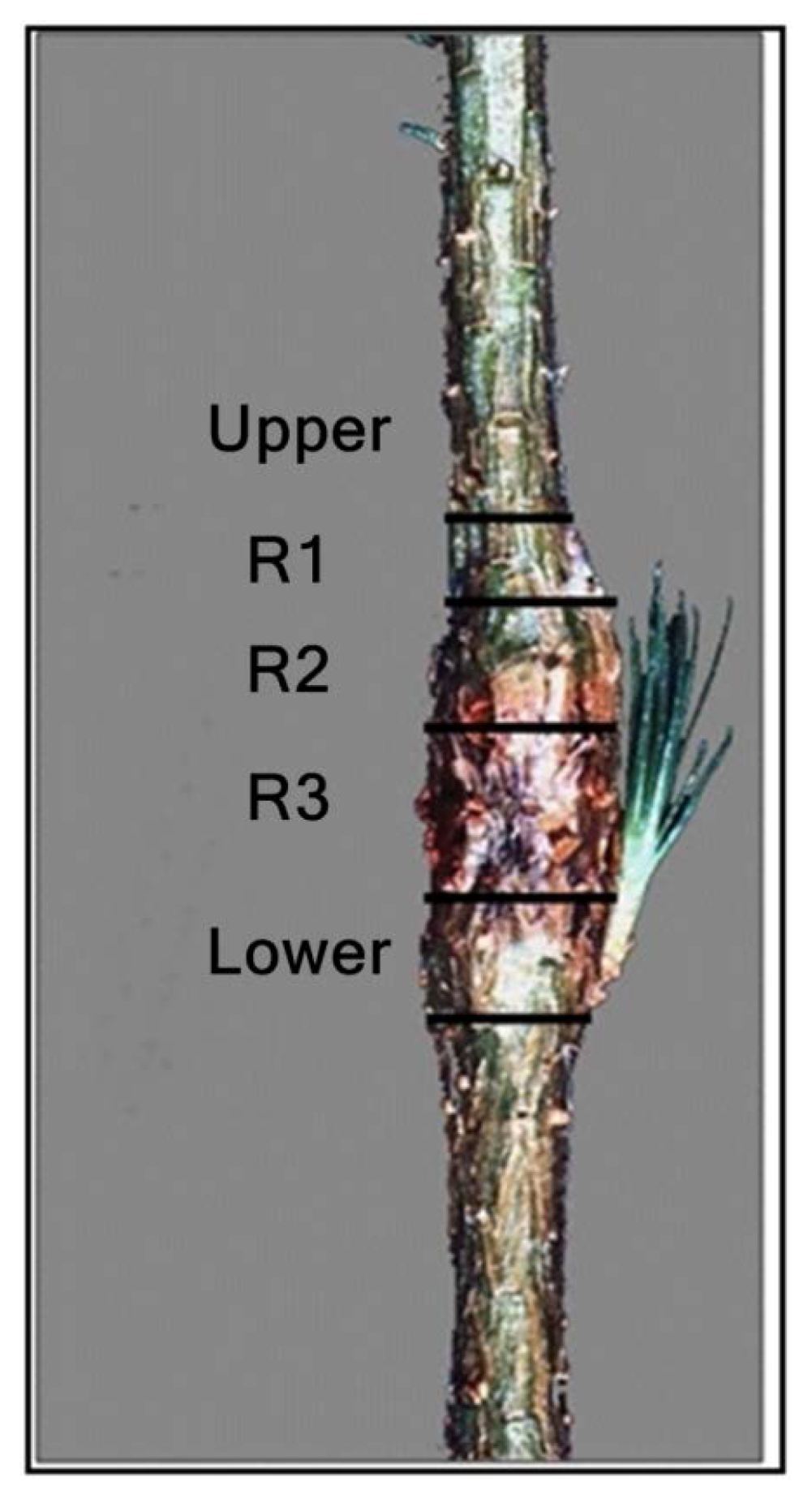
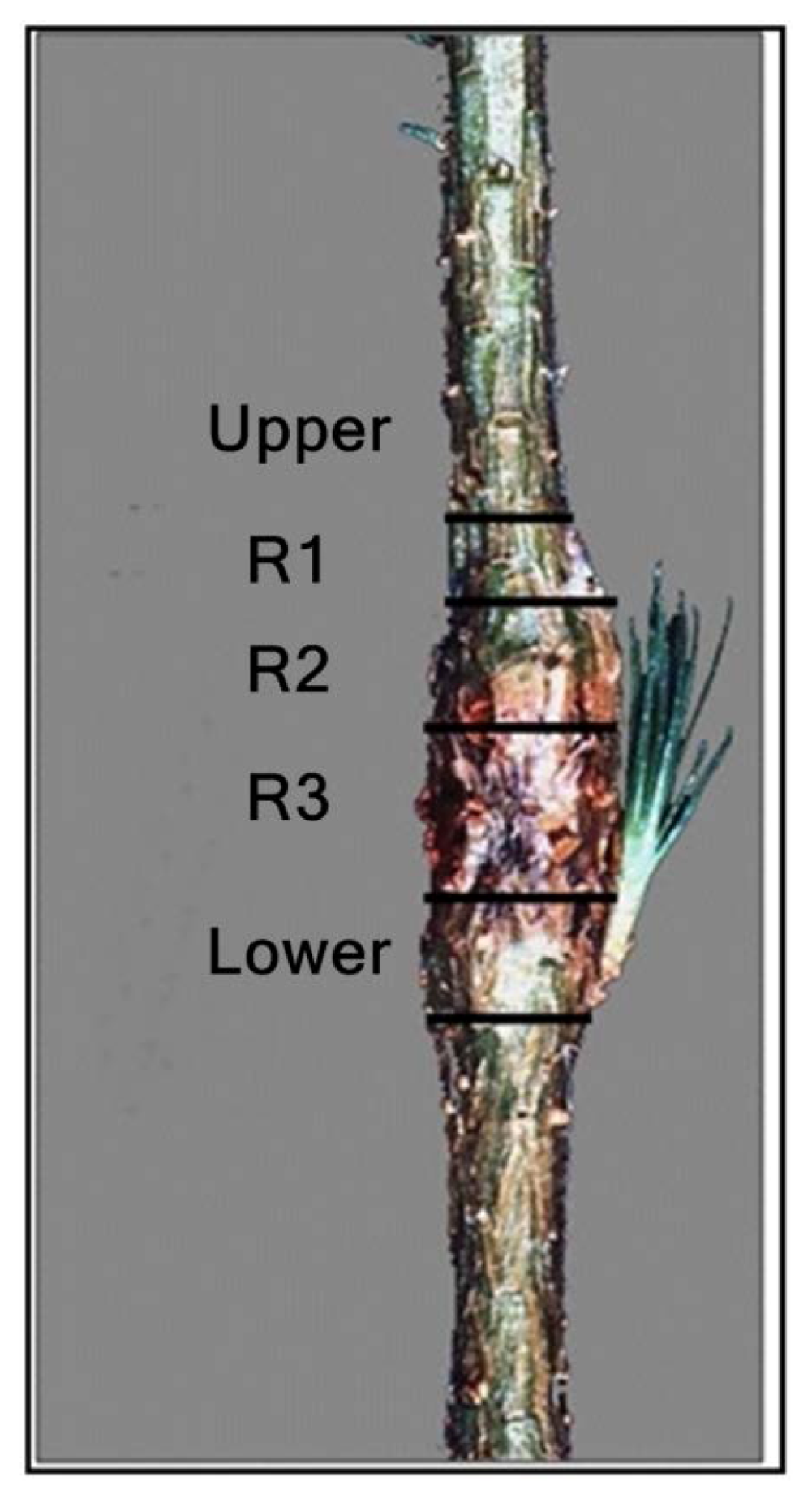
3. Results

{kind=link}
{kind=link}
{kind=link}
| Characteristics | Short galls | Rough galls | Smooth galls |
|---|---|---|---|
| Reaction zone(s) | |||
| Location | Innermost portion of the cortex | In the cortex near the epidermis | Radial to epidermis |
| Pattern | Widest in radial width and continuous around the circumference of the cambial area | Numerous small and narrow (radially) developing around the stem circumference with areas typical of susceptible reaction between reaction zones | Small and narrow conforming to the stem circumference, interconnected with areas typical of a susceptible reaction |
| Staining | Uniformly dark green | Very dark green | Light green |
| Periderm | Well-developed, encircling the RZ | Absent | Narrow and poorly developed along the innermost portion of the RZ |
| Fungal colonization in the RZ | Limited hyphae, both functional and nonfunctional based on the staining reaction. Haustoria few or absent | Absent | Limited hyphae present |
| Pattern | Haustoria and thick-walled hyphae abundant only in and adjacent to phloem ray cells | Distorted and limited haustoria radially inward from margins of the RZ | Dense colonization radially inward from RZ, hyphae multiseptate. |
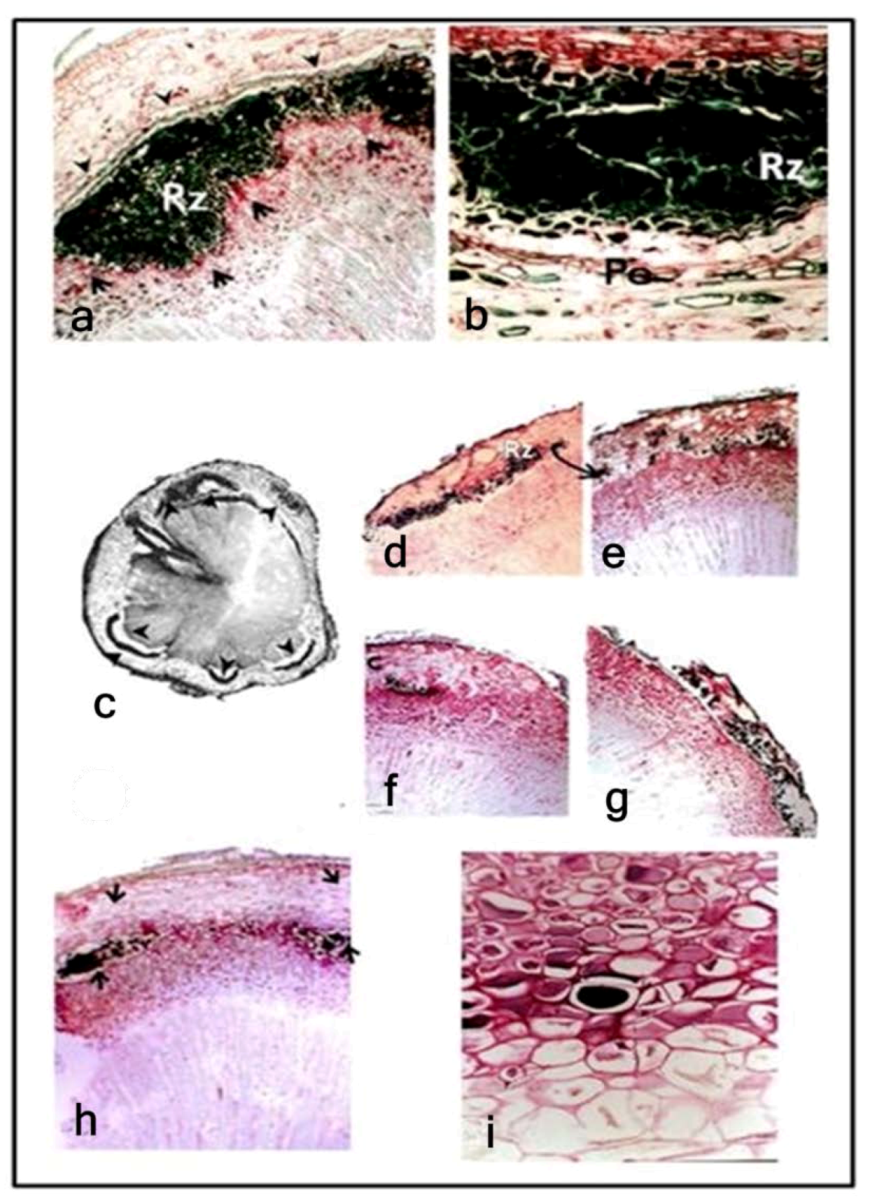
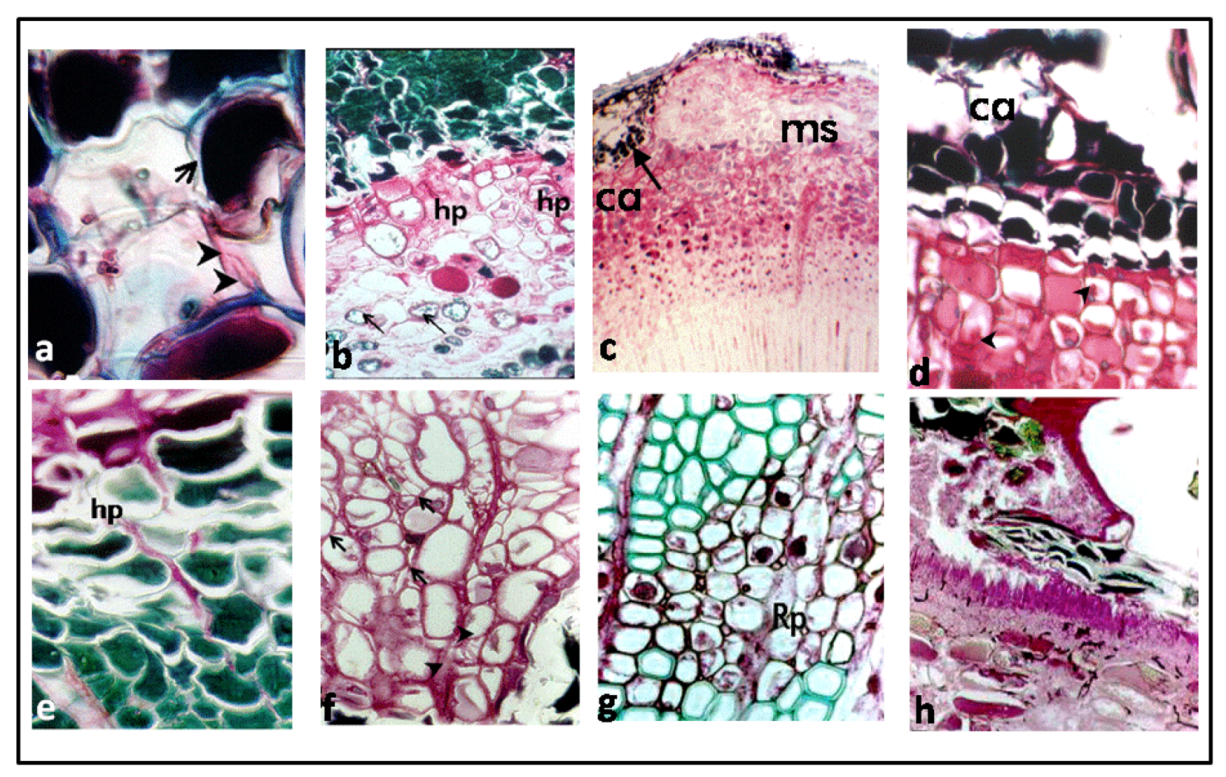
3.1. Reaction Zones (RZs)

3.2. Fungal Colonization
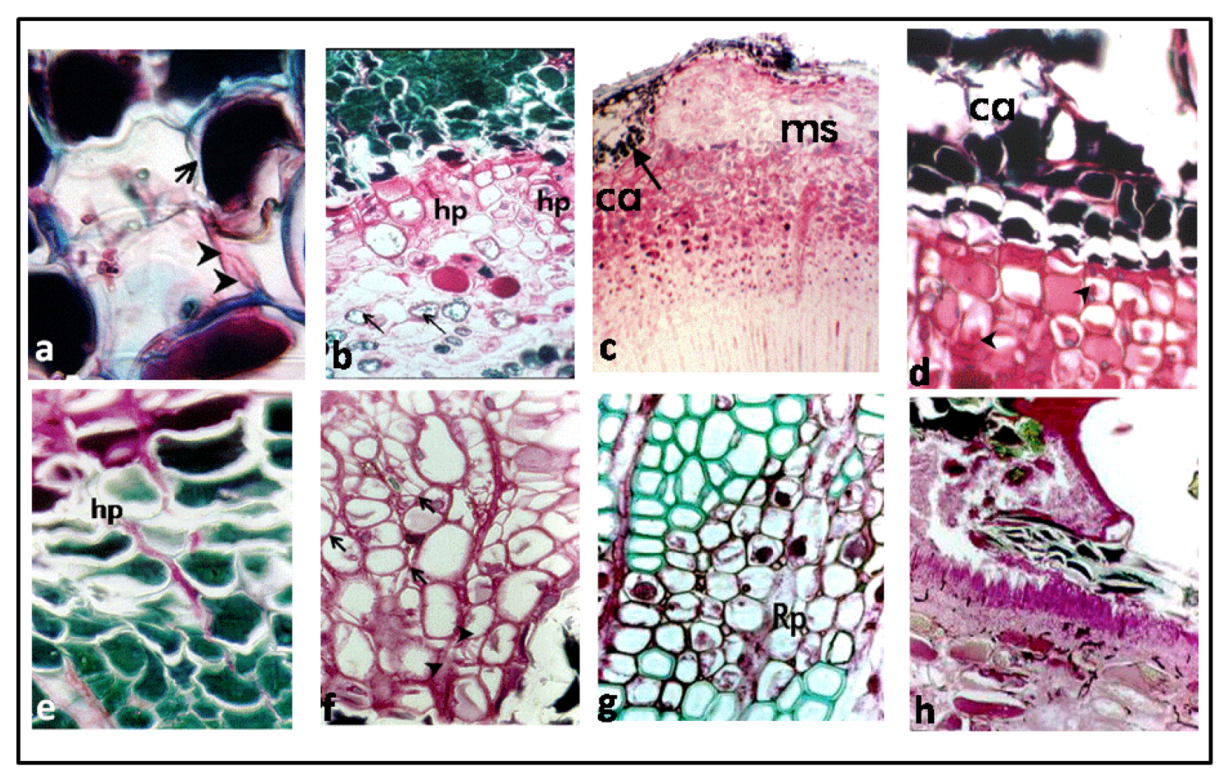
4. Discussion
5. Conclusions
Acknowledgments
References
- Powers, H.R., Jr.; McClure, J.P.; Knight, H.A.; Dutrow, G.F. Fusiform Rust: Forest Survey Incidence Data and Financial Impact in the South; Forest Service Research Paper SE 127; Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1975. [Google Scholar]
- Anderson, R.L.; McClure, J.P.; Cost, N.C.; Uhler, R.J. Estimating fusiform rust losses in five southeastern states. South. J. Appl. For. 1986, 10, 237–240. [Google Scholar]
- Hodge, G.R.; Schmidt, R.A.; White, T.L. Substantial realized gains from mass selection of fusiform rust-free trees in highly infected stands of slash pine. South. J. Appl. For. 1990, 14, 143–146. [Google Scholar]
- Powers, H.R.; Miller, T.; Belanger, R.P. Management strategies to reduce losses from fusiform rust. South. J. Appl. For. 1993, 17, 146–149. [Google Scholar]
- Pye, J.M.; Wagner, J.E.; Holmes, T.P.; Cubbage, F.W. Evaluation of the Effects of Fusiform Rust Research on Southern Pine Timber Investment Returns; Research Paper Forest Experiment Station; Research Work Unit SE 4851 Southeast: Research Triangle Park, NC, USA, 1994. [Google Scholar]
- Schmidt, R.A.; Powers, H.R.; Snow, G.A. Application of genetic disease resistance for the control of fusiform rust in intensively managed southern pine. Phytopathology 1981, 71, 993–997. [Google Scholar] [CrossRef]
- Schmidt, R.A.; Holley, R.C.; Klapproth, M.C. Results from operational plantings of fusiform rust resistant slash loblolly pines in high rust incidence areas in Florida Georgia. In Proceedings of the Rusts of Hard Pines Working Party Conference (S.2.06-10), Athens, GA, USA, 16–21 September 1985; pp. 33–41.
- Barber, J.C.; Dorman, K.W.; Bauer, E. Slash Pine Progeny Tests Indicate Genetic Variation in Resistance to Rust; Research Note 104; Southeastern Forest Experiment Station: Asheville, NC, USA, 1957. [Google Scholar]
- Schmidt, R.A.; Goddard, R.E. Preliminary Results of Fusiform Rust Resistance from Field Progeny Tests of Selected Slash Pines. In Proceedings of the 11th Southern Forest Tree Improvement Conference, Atlanta, GA, USA, 15–16 June 1971; pp. 37–44.
- Goddard, R.E.; Schmidt, R.A. Early identification of fusiform rust resistant slash pine families through controlled inoculation. In Proceedings of the 11th Southern Forest Tree Improvement Conference, Atlanta, GA, USA, 15–16 June 1971; pp. 31–36.
- Jewell, F.F. Inoculation of slash pine seedlings with Cronartium fusiforme. Phytopathology 1960, 50, 48–51. [Google Scholar]
- Screening for Disease Resistance as a Service for Tree Improvement Programs; Protection Report R8-PR8; Forest Health Protection, Southern Region, USDA Forest Service: Atlanta, GA, USA, 2004.
- Walkinshaw, C.H.; Dell, T.R.; Hubbard, S.D. Predicting Field Performance of Slash Pine Families from Inoculated Greenhouse Seedlings; Forest Experiment Station Research Paper SO-160U; Southern Forest Experiment Station: New Orleans, LA, USA, 1980. [Google Scholar]
- Miller, T.; Cowing, E.B.; Powers, H.R.; Blalock, T.E. Types of resistance and compatibility in slash pine seedlings infected by Cronartium fusiforme. Phytopathology 1976, 66, 1229–1235. [Google Scholar]
- Jackson, L.W.; Parker, J.N. Anatomy of fusiform rust galls on loblolly pine. Phytopathology 1958, 48, 637–650. [Google Scholar]
- Jewell, F.F.; Jewell, D.C.; Walkinshaw, C.H. Histopathology of the initiation of resistance-zones in juvenile slash pine to Cronartium quercuum f. sp. fusiforme. Phytopath. Med. 1980, 19, 8–12. [Google Scholar]
- Jewell, F.F.; Speirs, D.C. Histopathology of one- and two-year-old resisted infections by Cronartium fusiforme in slash pine. Phytopathology 1976, 66, 741–748. [Google Scholar]
- Schmidt, R.A.; Gramacho, K.P.; Miller, T.; Young, C.H. Components of Partial Resistance in the Slash Pine-Fusiform Rust Pathosystem. Phytopathology 2000, 90, 1005–1010. [Google Scholar] [CrossRef]
- Sohn, S.I.; Goddard, R.E.; Schmidt, R.A. Comparative Performances of Slash Pine for fusiform Rust Resistance in High Rust Hazard Locations. In Proceedings of the 13th Southern Forest Tree Improvement Conference, Raleigh, NC, USA, 10–11 June 1975; pp. 204–211.
- Knighten, J.L.; Young, C.H.; McCartney, T.C.; Anderson, R.L. Resistance Screening Center Procedures Manual: A Step-by-Step Guide to Materials and Methods Used in Operational Screening of Southern Pines for Resistance to Fusiform Rust; USDA Forest Service, Forest Health Protection: Asheville, NC, USA, 1988. [Google Scholar]
- Vaughan, R.E. A method for the differential staining of fungus and host cells. Ann. Mo. Bot. Gard. 1914, 1, 241–242. [Google Scholar] [CrossRef]
- Bao, J.R.; Lazarovits, G. Differential colonization of tomato roots by nonpathogenic and pathogenic Fusarium oxysporum may influence Fusarium wilt control. Phytopathology 2001, 91, 449–456. [Google Scholar] [CrossRef]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods; Lewis Publishers: Boca Raton, FL, USA, 1995. [Google Scholar]
- Lundquist, J.E.; Miller, T. Development of stem lesions on slash pine seedlings infected by Cronartium quercuum f. sp. fusiforme. Phytopathology 1984, 74, 514–518. [Google Scholar] [CrossRef]
- Miller, T.; Cowling, E.B. Infection and colonization of different organs of slash pine seedling by Cronartium fusiforme. Phytopathology 1977, 67, 179–186. [Google Scholar] [CrossRef]
- Struckmeyer, B.E.; Riker, A.J. Wound periderm formation in white-pine trees resistant to blister rust. Phytopathology 1951, 41, 276–281. [Google Scholar]
- Mullik, D.B. The non-specific nature of defense in bark and wood during wounding, insect and pathogen attack. Recent Adv. Phytochem. 1977, 11, 395–442. [Google Scholar]
- Kinloch, B.B.; Littlefield, J.L. White pine blister rust: Hypersensitive resistance in sugar pine. Can. J. Bot. 1977, 55, 1148–1155. [Google Scholar]
- Nelson, C.D.; Doudrick, R.L.; Nance, W.L.; Hamaker, J.M.; Capo, B. Specificity of host: Pathogen genetic interaction for fusiform rust disease on slash pine. In Proceedings of the 22nd Southern Forest Tree Improvement Conference, Atlanta, GA, USA, 14–17 June 1993; pp. 403–410.
- Wilcox, P.L.; Amerson, H.V.; Kuhlman, E.G.; Liu, B.H.; O’Malley, D.M.; Sederoff, R.R. Detection of a major gene for resistance to fusiform rust disease in loblolly pine by genomic mapping. Proc. Natl. Acad. Sci. USA 1996, 93, 3859–3864. [Google Scholar] [CrossRef]
- Kong, X. RAPD Mapping and Its Application to Slash Pine Breeding. Ph.D. Thesis, Texas A & M University, College Station, TX, USA, 1996; p. 124. [Google Scholar]
- Nelson, C.D.; Kubisiak, T.L.; Amerson, H.V. Unraveling and managing fusiform rust disease: A model 31 approach for coevolved forest tree pathosystems. For. Pathol. 2010, 40, 67–72. [Google Scholar]
- Kubisiak, T.L.; Anderson, C.L.; Amerson, H.V.; Smith, J.A.; Davis, J.M.; Nelson, C.D. A genomic amp enriched for markers linked to Avr1 in Cornartium quercuum f. sp. fusiforme. Fungal Gen. Biol. 2011, 48, 266–274. [Google Scholar] [CrossRef]
- Snow, G.A.; Jewell, F.F.; Eleuterius, L.N. Apparent recovery of slash and loblolly pine seedlings from fusiform rust infection. Plant Dis. Rep. 1963, 47, 318–319. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gramacho, K.P.; Miller, T.; Schmidt, R.A. Comparative Histopathology of Host Reaction Types in Slash Pine Resistant to Cronartium quercuum f. sp. fusiforme. Forests 2013, 4, 319-328. https://doi.org/10.3390/f4020319
Gramacho KP, Miller T, Schmidt RA. Comparative Histopathology of Host Reaction Types in Slash Pine Resistant to Cronartium quercuum f. sp. fusiforme. Forests. 2013; 4(2):319-328. https://doi.org/10.3390/f4020319
Chicago/Turabian StyleGramacho, Karina P., Thomas Miller, and Robert A. Schmidt. 2013. "Comparative Histopathology of Host Reaction Types in Slash Pine Resistant to Cronartium quercuum f. sp. fusiforme" Forests 4, no. 2: 319-328. https://doi.org/10.3390/f4020319