
Fire History of Appalachian Forests of the Lower St-Lawrence Region (Southern Quebec)


1
Département de Biologie and Centre d’Études Nordiques, Université Laval, 1045 Avenue de la Médecine, Québec, QC G1V 0A6, Canada
2
Direction des Inventaires Forestiers, Ministère des Forêts, de la Faune et des Parcs, 5700 4e Avenue ouest, Québec, QC G1H 6R1, Canada
*
Author to whom correspondence should be addressed.
Forests 2017, 8(4), 120; https://doi.org/10.3390/f8040120
Submission received: 3 March 2017
/
Revised: 31 March 2017
/
Accepted: 5 April 2017
/
Published: 11 April 2017
Abstract
:Sugar maple (Acer saccharum) forests are among the main forest types of eastern North America. Sugar maple stands growing on Appalachian soils of the Lower St-Lawrence region are located at the northeastern limit of the northern hardwood forest zone. Given the biogeographical position of these forests at the edge of the boreal biome, we aimed to reconstruct the fire history and document the occurrence of temperate and boreal trees in sugar maple sites during the Holocene based on soil macrocharcoal analysis. Despite having experienced a different number of fire events, the fire history of the maple sites was broadly similar, with two main periods of fire activity, i.e., early- to mid-Holocene and late-Holocene. A long fire-free interval of at least 3500 years separated the two periods from the mid-Holocene to 2000 years ago. The maple sites differ with respect to fire frequency and synchronicity of the last millennia. According to the botanical composition of charcoal, forest vegetation remained relatively homogenous during the Holocene, except recently. Conifer and broadleaf species coexisted in mixed forests during the Holocene, in phase with fire events promoting the regeneration of boreal and temperate tree assemblages including balsam fir (Abies balsamea) and sugar maple.
1. Introduction
Deciduous and mixed forests of the temperate zone of eastern North America are ecosystems rarely impacted by fire [1]. Occasional fires caused by Euro-American settlers and aboriginal populations were reported for New England, southern Quebec and Ontario [2,3,4,5,6,7,8,9], whereas natural fires were seldom recorded over the last millennia and centuries [5,10,11].
Until recently, northern sugar maple (Acer saccharum Marsh.) forests were considered ecosystems maintained at equilibrium [12] under the regime of canopy-gap dynamics [13,14,15]. However, field evidence suggests that sugar maple is adapted to various sun-and-shade regeneration environments [16] associated with logging [17,18,19], abandoned fields [20], and fire [21,22,23]. The rise and dominance of sugar maple communities deduced from pollen data have been associated with reduced fire activities, particularly during the mid-Holocene [24,25,26,27]. Although realistic on biogeographical grounds, the fire/sugar maple connection during the Holocene as reported from pollen and microcharcoal data remains to be tested with direct evidence from botanically-identified macrofossil data including macrocharcoal. Given that macrocharcoal of sugar maple and other hardwood species as well as conifer species can be identified at the genus if not the species level, there is an opportunity to evaluate the relationship between past forest communities such as sugar maple forests and fire events using soil macrocharcoal analysis [28,29,30].
In this perspective, the main objective of this study is to reconstruct the Holocene fire history and tree species composition of two Appalachian sites dominated by sugar maple using soil macrocharcoal analysis. Sugar maple forests are widely distributed in southern Quebec, south of the St. Lawrence River, where they grow on well-drained podzolic soils. Along the climatic gradient running from the south-west to the north-east in this region, the cover of sugar maple forests decreases significantly to the point that only small patches of maple stands are found on the Appalachian uplands at the transition between the cold-temperate forest zone and the boreal forest zone. We have compared the long-term fire record of the two sugar maple sites to evaluate the frequency and synchronicity of fire events since initial postglacial forest establishment using 14C-dated charcoal of tree species. The selection of sugar maple forest sites for this analysis is part of a larger research program dedicated to the reconstruction of fire history of the temperate forest zone of southern Quebec based on local sugar maple sites. The study is also motivated by the hypotheses stating that the species is forming, in southern Quebec, the natural regional forest at equilibrium with the present climate [12]. Whether the maple forests in eastern North America are maintained at equilibrium, in the absence of allogenic disturbances, is at stake in the current context of the changing climate. This study is one step ahead for a better assessment of the dynamic biogeography of the cold-temperate deciduous forest of eastern North America.
2. Materials and Methods
2.1. Study Area
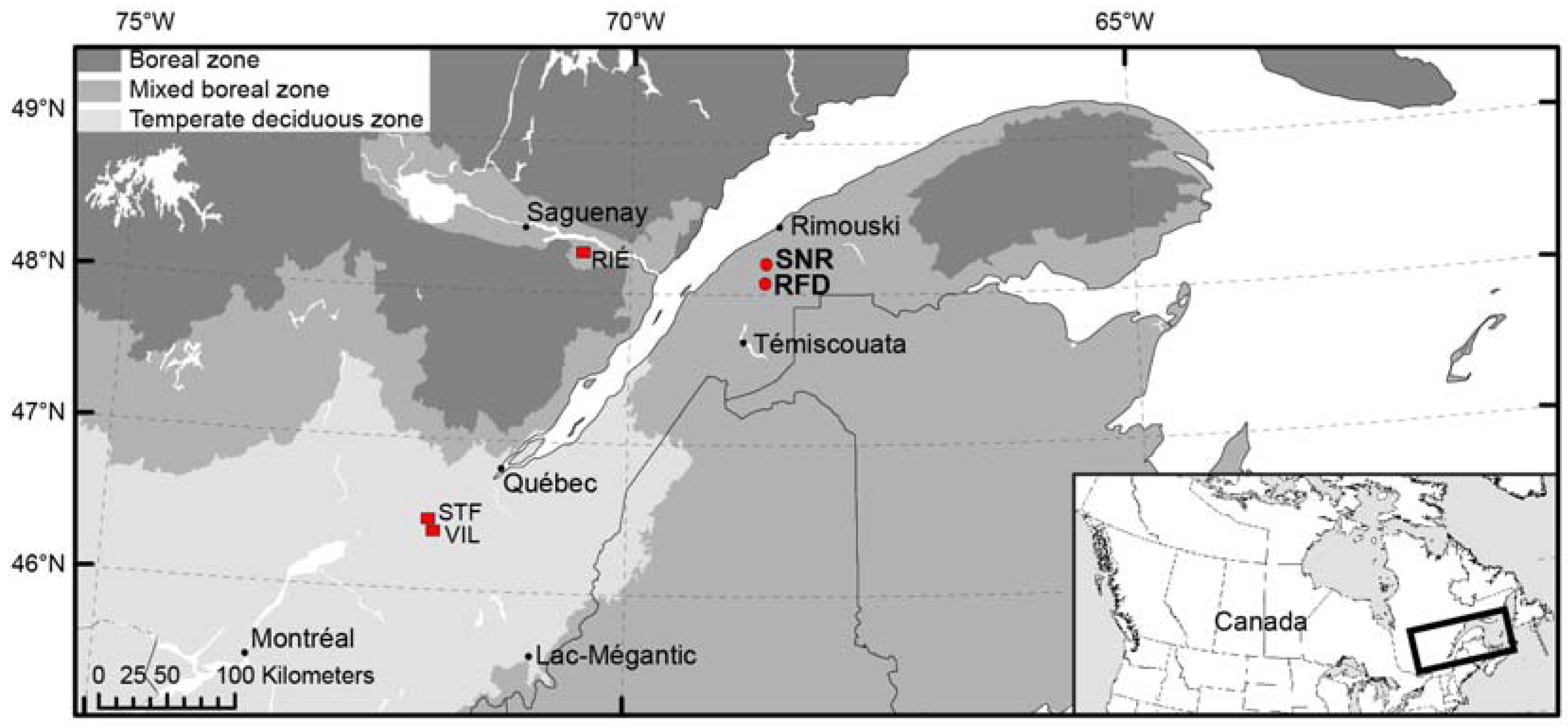
The Lower St-Lawrence region (Figure 1) is characterized by an Appalachian-type landscape composed of undulating, subparallel hills 50 to 500 m a.s.l. (above sea level). Conifer forests generally dominate in low altitude sites. Eastern white cedar (Thuja occidentalis L.) is abundant and is often found in association with balsam fir (Abies balsamea (L.) Mill.), red spruce (Picea rubens Sargent) and black spruce (Picea mariana (Mill.) B.S.P.). Mixed forests, the most abundant forest type in this region, are located in lowland and upland sites. Several tree species are growing in mixed forests, mostly balsam fir, trembling aspen (Populus tremuloides Michx.), paper birch (Betula papyrifera Marsh.), red maple (Acer rubrum L.) and yellow birch (Betula alleghaniensis Britton). Sugar maple forests are generally located uphill or on small well-drained summits above ~275 m a.s.l. Sugar maple often forms pure stands, although red maple, yellow birch, white spruce (Picea glauca (Moench) Voss) and balsam fir are present as companion species. American beech (Fagus grandifolia Ehrh.), white ash (Fraxinus americana L.), American basswood (Tilia americana L.), hemlock (Tsuga canadensis (L.) Carrière) and other mesophytic species associated with maple forests of the temperate zone are absent in this region.
Two sugar maple sites, an old-growth site and an even-aged site, were considered in this study. The sites are situated in the Appalachian uplands, south of the city of Rimouski, in the Lower St. Lawrence region, about 14 km from each other. The forest canopy and ground layer vegetation of the even-aged site are largely dominated by sugar maple, whereas the species is less prevalent in the old-growth site where conifers coexist both in the canopy and the ground layer.
The first site is a sugar maple stand of the Nicolas-Riou Seigneury (SNR, 48°12′23″ N, 68°39′25″ W) maintained in a natural state (no logging disturbances) and selected as the natural, old-growth stand of this study. The Nicolas-Riou site is dominated by a boreal flora of trees, shrubs and herbs, although sugar maple is the dominant tree species (Table 1). It is located on a south-facing hillside 310 m a.s.l. with 15° slope inclination. The organic horizon has an average depth of 7.2 (±4.0) cm, a pH of 4.6 and is composed of leaf litter, FH (mixed fibrous and humic) horizon, and well decomposed H horizon (≤1 cm in 15 out of the 25 microsites). The podzolic B horizon is a clay loam with a pH of 4.6.
The even-aged sugar maple stand of the Duchénier Faunal Reserve (RFD, 48°04′35″ N, 68°40′22″ W) is situated 300 m a.s.l., and located on a low northwest slope (5°). The organic horizon is relatively thick (9.8 ± 3.7 cm), with a pH of 4.5. The FH and H horizons (~3 cm) are present in 85% and 80% of the subplots, respectively. The podzolic B horizon is also a clay loam with a pH of 5.5. This stand is a secondary, anthropogenic forest (Table 1) formed after a clear-cut in 1942 and tree thinning in 1997. Data from the first land survey in 1870 indicates that the site was occupied by a mixed forest where balsam fir was the dominant tree species [31]. In 1930, foresters of the Price Brothers Company surveyed two plots at this site. Paper birch and balsam fir were identified as the dominant tree species in the two plots with maple (sugar maple and/or red maple) as companion species [32].
2.2. Field Sampling and Sample Treatment
The two sites were sampled in 2013. Charcoal sampling included 25 microsites distributed within a rectangular plot of 1000 m2 (20 m × 50 m), with 22 of the microsites positioned along the upper and lower lines and the other three microsites at the center of the plot. A 20 cm × 20 cm sample of the organic horizon (monolith), from the soil surface to the uppermost part of the mineral soil, was used for charcoal extraction at each microsite. After sampling the organic horizon at each microsite, two superposed mineral soil cores were taken with a soil corer, allowing the extraction of 750 cm3 of soil. The corresponding cores A and B were sampled at a depth of 0–15 cm and 15–30 cm from the mineral soil surface, respectively. The organic samples and the mineral cores (A and B samples of the 25 microsites) were immersed in a 2%–5% solution of NaOH for at least 12 h (or boiled for 15 min) to disperse the soil aggregates and to eliminate the organic matter during screening. Each sample was sieved under running water using 2 mm and 4 mm mesh screens. Charcoal pieces >2 mm, corresponding to local charcoal formed in situ and deposited in the sampling sites and not transported over long distances [33], were extracted manually under a binocular microscope and dried at room temperature. When charcoal fragments of the organic samples were particularly abundant, a maximum of 100 pieces were selected among which 30 were selected at random for botanical identification. The selected fragments were sectioned to examine the anatomical planes under an incident light microscope at 200× and 500×. The identification criteria based on wood anatomy [34,35,36] as well as the charcoal collection of the Centre d’études nordiques (Université Laval, Québec, QC, Canada) were used for the botanical identification of all charcoal pieces at the genus level, and the species level when possible. The largest parenchyma rays of charcoal fragments identified as Acer saccharum are seven to eight cells wide, whereas fragments identified as Acer sp. have narrower parenchyma rays. According to lab observations, sapling and small branch wood do not produce large rays as in mature sugar maple material. Therefore, small stems of sugar maple cannot be differentiated from soft maples. A number of charcoal fragments display a lamellar form. On these fragments, two of the three anatomical planes are too narrow for proper identification although spiral thickenings, a structure present in vessel elements of maple species, are usually visible. Unidentified shrubs of the Betulaceae family may include Corylus cornuta Marsh., Alnus sp., Ostrya virginiana (Mill.) K. Koch or Betula shrub.
2.3. Charcoal Dating and Fire History
The selection of charcoal fragments (minimal weight of 1.5 mg, ideally 5 mg) for 14C dating (accelerator mass spectrometry (AMS) method) was done according to the species or genus present in the sites and the location in the two plots. Only one charcoal fragment of a given species per core (i.e., cores A and B) was retained for dating. The selection of charcoal fragments was also based on equal representation of the main tree species distributed in the surface and mineral soil compartments. The selected fragments were pretreated to CO2 in the 14C laboratory of the Centre d’études nordiques and dated either at the Keck Carbon Cycle AMS Facility (University of California, Irvine, California, USA) or at the Center for Applied Isotope Studies (University of Georgia, Athens, GA, USA). Calib7.02 software [37] and the databases IntCal13 [38] were used for the calibration of the 14C dates in calendar years. For each 14C date (±2 σ), the distribution of the calendar ages and their associated probabilities were considered for calibration. Because of the changing concentration of 14C in the atmosphere caused by solar activity [39], the distribution of the calendar ages is often divided in multiple probabilistic states (age intervals). The mean year of the interval having the highest probability was determined as the calendar age of each radiocarbon-dated charcoal. Sixty-five years were added to the standard 0 year BP (1950 AD) to present the results in calendar years before 2015 AD. 14C dates with overlapping calendar years within the highest probability interval were considered as originating from the same fire. Tree-ring dating (based on the collar age of >10 trees in the RFD site; [40]) and 14C dates of charcoal were used concurrently to date the most recent fires (≤200 cal. BP).
Given the peculiar distribution of charcoal in the forest environment affected by blowdowns and other soil disturbance events that are altering the original soil stratigraphy, it is thus necessary to date the greatest number of fragments in order to record all the fires that occurred at the site [29]. It is common practice to use an indirect method to evaluate the total number of fires that occurred at a site, because it is not possible to date all the fragments (and the fires) accumulated in the site since initial postglacial establishment of forest trees. The random sampling of charcoal fragments does not guarantee that all fires are detected, so the construction of an accumulation curve is useful to evaluate whether the number of detected fires corresponds to the number of all the fires that occurred at the site. The accumulation curve indicates the number of fires detected according to the number of charcoal fragments dated. When reaching an asymptote, the accumulation curve then suggests that most of the local fires are theoretically detected. The accumulation curve of the two sites was calculated with the accumresult function using the random method of the BiodiversityR package in the R software [41,42]. This function plots 100 accumulation curves by random resampling of the 14C dates of each site. An evaluation of the total number of theoretical fires at each site was done using the nonlinear regression:
where F(n) is the number of fires and n the number of dated charcoal. F(max) is the factor controlling the flattening of the negative exponential curve that is considered as the estimate of the real number of fires (asymptote) [28,29,43]. k is the constant controlling the shape of the curve.
F(n) = F(max) (1−ekn)
3. Results
3.1. Charcoal Species of the Nicolas-Riou Seigneury Site
Charcoal fragments were recovered from the organic soil horizon of 16 out of the 25 microsites. About 25% of the pieces are conifers, mostly balsam fir, eastern white cedar and spruce (Picea sp.) (Table 2). More than half of the fragments are broadleaf species. Birch (Betula sp.) is the main taxon dominating the organic compartment with more than 60 pieces, but >50% of the pieces were found in one microsite. Thirty-three sugar maple pieces and three fragments of maple (either red maple, shrub maples or small branches of sugar maple) were extracted from one microsite. The distribution of broadleaf charcoal of the organic compartment is thus heterogeneous at the site scale. At least 20% of charcoal pieces of the organic compartment were not identified because of vitrified conditions, mineral coatings or amorphous structure (bark and pith pieces). Six out of the 10 monoliths contain charcoal predominantly located at the contact with the uppermost mineral horizon or in the lower part of the monoliths.
A total of 211 macrocharcoal pieces were recorded in the mineral soil, including 154 (73%) and 57 (27%) pieces extracted from A and B cores, respectively. About 50% of the macrocharcoal was located in one microsite, whereas the remaining 50% was in the other microsites. No sugar maple charcoal was found in the mineral soil. However, spiral thickenings observed in two pieces of unidentified broadleaf charcoal suggest the possible presence of sugar maple. Except for some shrub pieces (possibly Corylus cornuta) and maples, most charcoal fragments are birch wood. It is worth noting, however, that 27 out of the 35 birch charcoal pieces are located in one microsite, possibly coming from the same individual. Among the conifer charcoal, spruce, balsam fir and eastern white cedar were identified. Twenty-nine pieces of bark, or charcoal pieces without secondary xylem, also were recorded.
3.1.1. Fire Periods
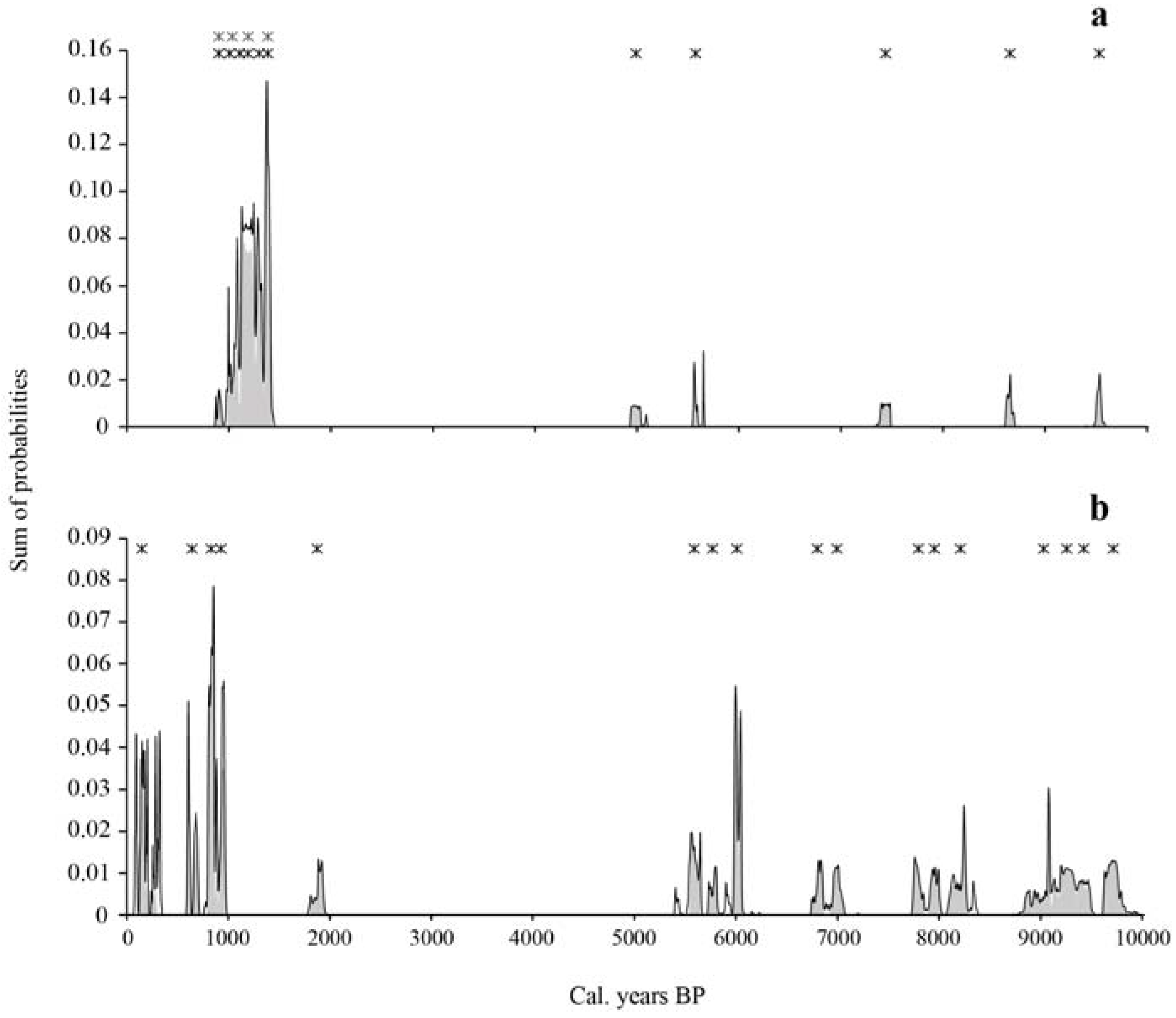
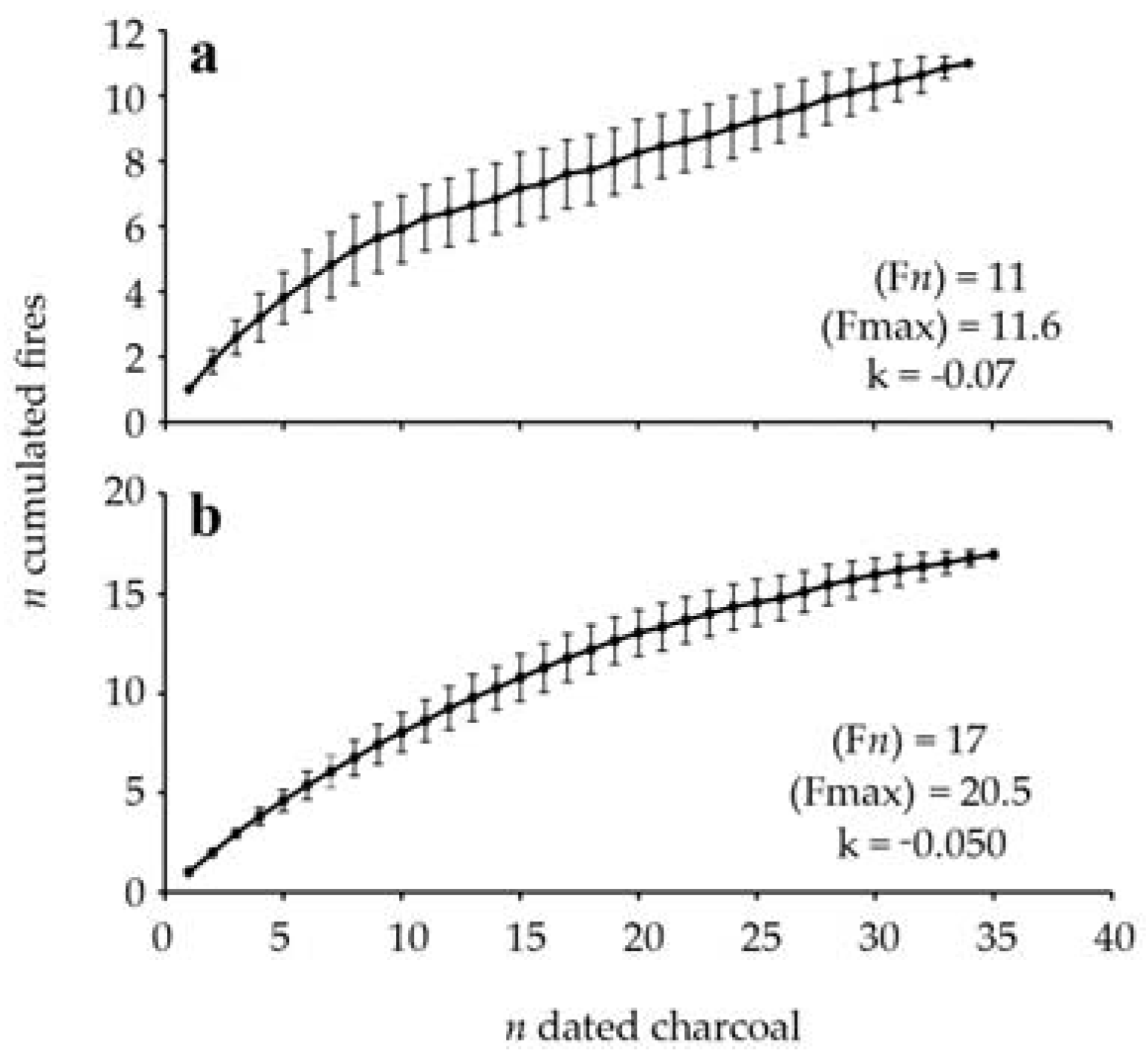
Eleven fires were identified from the charcoal assemblage, including five fires that occurred between 9500 and 5000 cal. BP, at an interval varying between 590 and 1860 cal. years (mean interval of 1135 ± 550 cal. years) (Table 3, Figure 2). A fire-free period of 3600 cal. years occurred between 5000 and 1400 cal. BP. According to the range of overlapping cal. years, at least four if not six fires were recorded in the late Holocene, between 1400 and 900 cal. BP, with a corresponding mean interval of 100–125 cal. years. No fires were detected since approximately 900 cal. BP; we can infer that the present maple forest site is growing without the influence of fire since that period. The oldest fires were recorded from macrocharcoal buried in the mineral compartment. According to the accumulation curve, the number of detected fires corresponds to the number of estimated fires (Figure 3).
3.1.2. Macrocharcoal Assemblage
The five fires of the oldest fire period are each represented by one charcoal fragment, i.e., spruce, broadleaf species, and unidentified conifer (Table 3). Macrocharcoal taxa of the recent fire period are birch, spruce and balsam fir. The only dated sugar maple charcoal is located at the interface between the organic horizon and the upper part of the mineral solum, and comes from a fire dated 1185 cal. BP. The species was part of a mixed forest along with Canada yew (Taxus canadensis), balsam fir, spruce and Corylus cornuta. Charcoal fragments produced by a 1095 cal. BP fire were part of a forest flora dominated by conifers (balsam fir and unidentified conifers) and broadleaf species including maple (shrub or sugar maple branch) and birch, whereas the two most recent fires (1000 and 900 cal. BP) burned conifer trees and possibly broadleaf trees.
3.2. Charcoal Species of the Duchénier Faunal Reserve Site
Twenty-three out of the 25 organic horizons contain charcoal. At least 56% of the charcoal from the organic compartment is conifers, 21% broadleaf pieces and 21% unidentified fragments or bark, pith or fungi (Table 2). Spruce, balsam fir and eastern white cedar are among the conifers identified. Half of the 62 birch pieces were located in one monolith, and 19 maple and two sugar maple charcoal pieces were also recorded in the organic compartment. Several charcoal pieces show a lamellar structure with only one anatomical plan, where it is however possible to observe spiral thickenings in the vessel elements possibly corresponding to maple charcoal. A small number of charcoal fragments corresponds to Corylus cornuta. The number of charcoal fragments in the 10 monoliths varies from 1 to more than 100. However, most fragments are located at the contact with the mineral solum and in the basal part of the organic horizon. Twenty-four and 15 charcoal pieces were extracted from the A and B cores, respectively. Two to six pieces were present in each mineral core, and despite their low representation, charcoal fragments of the mineral cores were well distributed in the plot. Pieces of broadleaf species include one birch, one sugar maple and five Betulaceae shrubs (Table 2). Nineteen pieces of spruce, balsam fir and eastern white cedar were also inventoried.
3.2.1. Fire Periods
Seventeen fires were identified over the last 9700 years. Twelve fires occurred between 9700 and 5600 cal. BP, at an interval varying between 160 and 800 cal. years (mean interval of 375 ± 280 cal. years) (Table 4, Figure 2). A fire-free period of 3700 cal. years followed the last mid-Holocene fire, from 5600 to 1900 cal. BP. Five fire events were recorded over the last 1900 cal. years, at a mean interval of 350 ± 370 cal. years. According to the accumulation curve of all the dated macrocharcoal fragments, a difference of three fires has been found between the number of detected fires and the number of estimated fires (Figure 3).
3.2.2. Macrocharcoal Assemblage
The oldest fires are represented by vitrified conifer charcoal (including spruce) and shrubs of the Betulaceae family (Table 4). Birch and balsam fir are recorded in the charcoal assemblage around 7000 and 6000 cal. BP. The sole buried sugar maple charcoal was dated 5600 cal. BP, and confirms the presence of the species during the mid-Holocene. Fires of the last 1900 cal. years are represented by birch, shrubs of the Betulaceae family and Corylus cornuta charcoal. Balsam fir, eastern white cedar and spruce are also present and relatively abundant. Lamellar charcoal fragments, indicative of broadleaf species, including sugar maple, and dated 650 cal. BP, suggest the presence of the latter species in the recent past.
4. Discussion
The charcoal data of the sugar maple sites allow us to reconstruct the fire history of a section of the Appalachian uplands of the Lower St. Lawrence region where 11 and 17 fire events were recorded in two sites during the Holocene, respectively. According to the accumulation curves, it is possible that our sampling missed a small number of fires. At both sites, a greater number of charcoal pieces was found in the organic compartment than in the mineral soil. The oldest charcoal pieces were located in the mineral compartment, but a 1435 cal. years old charcoal piece was found buried in the organic horizon of the Nicolas-Riou site (Old-growth site). Vitrified or altered charcoal fragments were among the oldest fragments dated. The internal structure of altered charcoal is probably associated with long-term chemical processes occurring in the mineral spodic horizons. Despite the difference in the number of fire events, the overall fire history of both sites is relatively similar. At the Nicolas-Riou site, fire events occurred during two well-bracketed periods of the Holocene, the oldest between 9500 and 5000 cal. BP and the most recent between 1400 and 900 cal. BP, the two periods being separated by a long quiescent, fire-free interval of 3600 cal. years. The pattern of fire frequency at the Duchénier site (even-aged site) is similar to that of the Nicolas-Riou site. The oldest fire period extended from the early Holocene to mid-Holocene, from 9700 to 5600 cal. BP, and the most recent fire period spanned the last 1900 cal. years. A long fire-free interval of 3700 cal. years also separated the two fire periods, which suggests that the millennia succeeding the mid-Holocene were rather favorable for the prevalence of fire-free forest conditions. The most important difference between the two sites is the contrasting fire history of the last 2000 cal. years. The Nicolas-Riou site experienced several fires between 1400 and 900 cal. BP and none during the last 900 years, whereas the Duchénier site was impacted by several fires over the last 1000 cal. years.
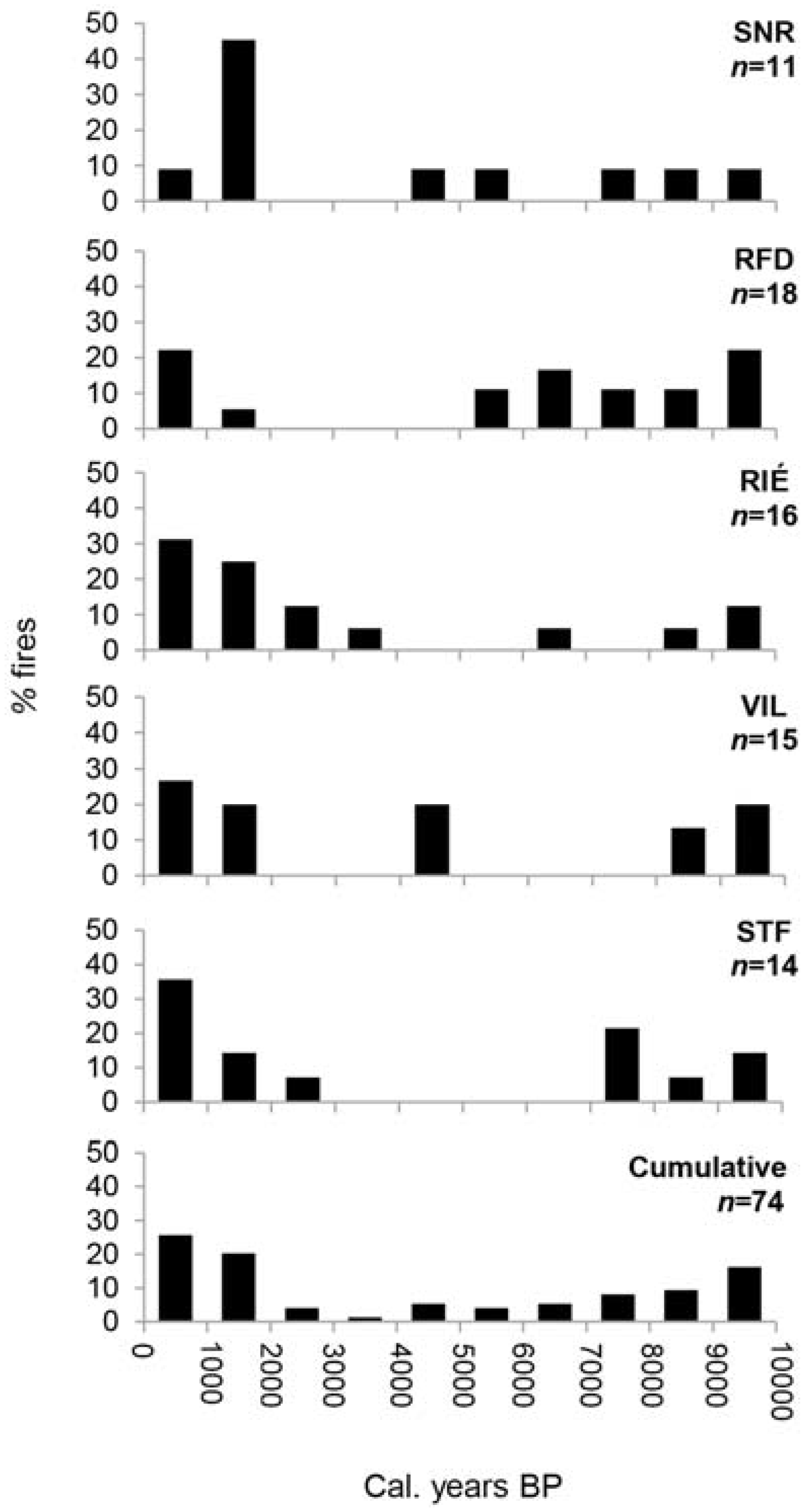
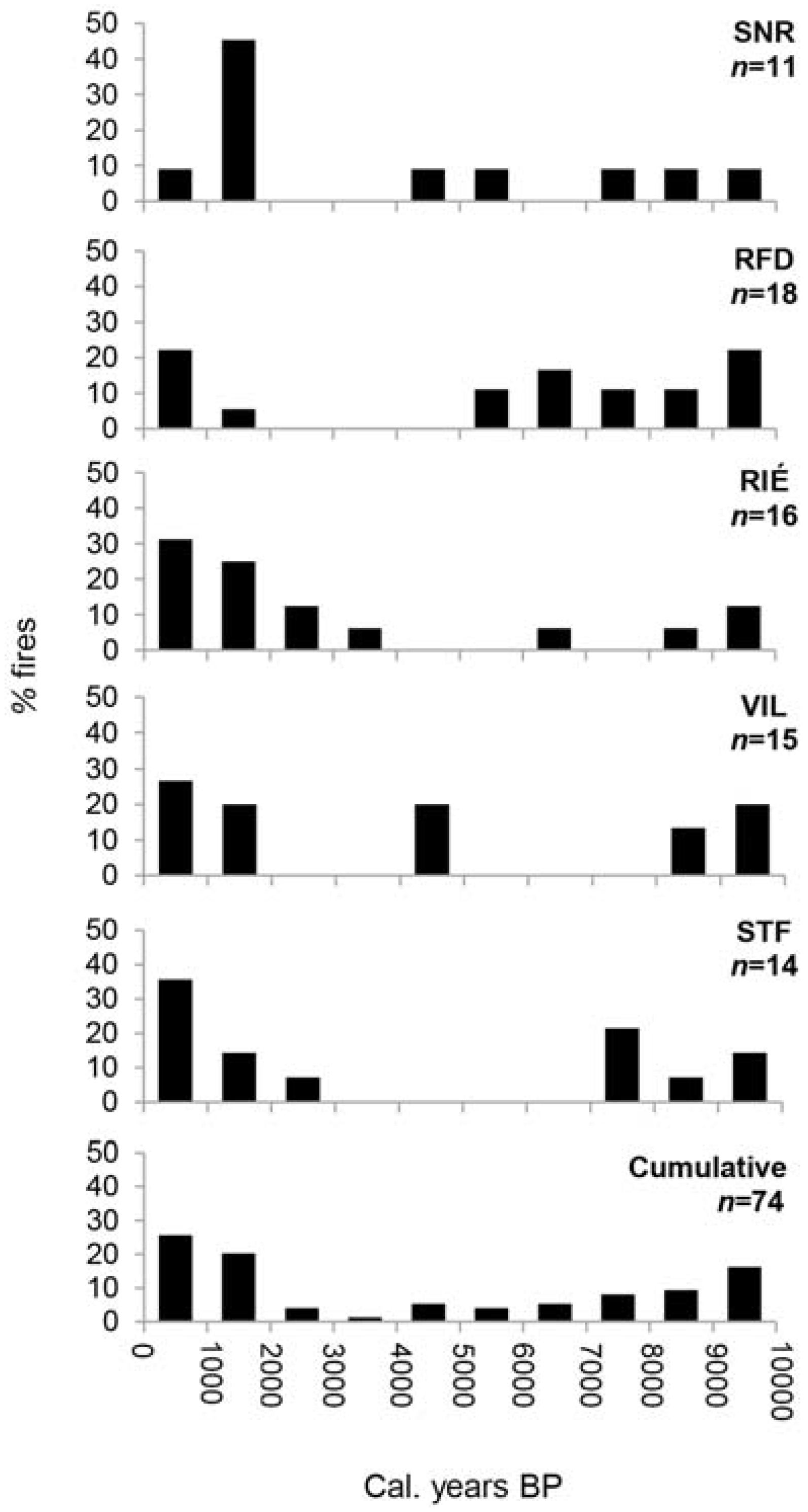
According to recently published reports and this study, fires were frequent in the early Holocene in the current maple sites distributed across southern Quebec according both to macrocharcoal [23,27,30] (Figure 4) and micro- and mesocharcoal [25] data. The mid-Holocene was a period less-prone to fire occurrence both in the study area and the areas close to our Appalachian sites in the Saguenay [23] and the Quebec City [30] areas. In southernmost Quebec, Elkadi [25] suggested declining fire occurrence at that time—as deduced from microcharcoal and pollen of Mégantic lake sediments—to be the result of the arrival and expansion of sugar maple forests which are thought to reduce fire propagation. Forests of broadleaf trees are generally less susceptible to burn and to propagate fire over large areas because of the stem architecture of dominant tree species that prevent fire from expanding vertically, the fuel type produced including litter and ground vegetation. Except for the influence of recent man-made disturbances, which were probably important locally, the early and late Holocene of the Appalachian uplands represent periods of relatively moderate fire activity before and after the long fire-free interval of the mid-Holocene. The fire events of the two Appalachian sites were not synchronized although they occurred mainly during the early and late Holocene which most likely were climatically fire-prone periods.
The fire regime of both sites has been characterized by mean fire intervals of 600 (RFD) and 800 (SNR) cal. years during the last 10,000 cal. years. These figures are means with large standard deviations which, no doubt, are less significant ecologically if one only considers the aggregation of fire events during the two main fire periods. Nevertheless, the mean fire intervals do suggest the development of fire-free forest communities that have been relatively stable and uniform during several thousand years, a situation less prone to the establishment of postfire pioneer species at the landscape scale. It is only when repetitive fires occur that the latter species invade forest landscapes, as has been the case in the St. Lawrence valley during the last 2000 years and the historic period of the last centuries [17,19,44]. Frequent fires (every 100–125 cal. years) occurred at the Nicolas-Riou site between 1400 and 900 cal. BP, inducing stand dynamics different to the fire-free situation that prevailed after, for about 900 cal. years. The vegetation cover of the Appalachian uplands was probably forming a mosaic of old-aged, postfire stands possibly impacted by occasional insect outbreaks and wind storms. The Nicolas-Riou site thus represents an example of old-growth forest likely maintained by the process of canopy-gap dynamics. In contrast, the Duchénier sugar maple forest has been impacted by several fires over the last centuries and clear cutting in the 20th century.
It is possible that most fires recorded in the Rimouski area were small in extent, but no paleoecological evidence exists to support this contention. Although the type of fire that occurred in the two sites is unknown, it is probable that surface fires were the main type of disturbance. Forests with hardwood species are less likely to propagate fire due to the greater humidity of the forest cover and the discontinuity of fuel between the soil and the crown. Furthermore, the rather scarce charcoal record of sugar maple and other hardwood species having similar architectural branching may be attributed, at least in part, to the impact of surface fires. Charcoal production in boreal conifers (spruce, fir, cedar) is facilitated by their architecture which includes the formation and long-term residence of branches along the stem, particularly near the ground.
With these reservations in mind, the charcoal assemblage suggests that a mixed forest composed of conifers and broadleaf species likely prevailed between 9500 and 5000 cal. BP in the Nicolas-Riou site. Much later, during the late Holocene, a conifer forest dominated by balsam fir and spruce, and accompanied by birch, a birch–balsam fir forest (it is not possible to distinguish yellow birch and paper birch with current anatomical characters), grew at the site around 1400 cal. BP. Sugar maple was also present at this site at 1300–1200 cal. BP. It is probable that a conifer forest was maintained at the site until at least 900 cal. BP. The present forest is a sugar maple–yellow birch–balsam fir stand that developed progressively from a boreal flora dominated by fir and spruce.
The Duchénier site was also occupied by a mixed forest dominated by fir, spruce and birch between 9700 and 5800 cal. BP. The arrival of sugar maple at the site at least 5600 cal. BP emphasizes the «warm» affinity of the forest. No data are available on the tree composition of the site during the following ca. 3600 cal. years, and until 2000 cal. BP when fire again burned the site. A mixed forest composed of conifers, including balsam fir and eastern white cedar, and broadleaf species, i.e., birch and sugar maple among others, grew at the site until at least 200 years ago. Both the conifer (and bark of unknown origin) charcoal fragments produced during the last fire and the unburned and uncut woody debris [40] suggest the presence of a conifer forest. Field relevés of the Price Brothers Company in 1930 [32] report the dominance of paper birch accompanied by maple and balsam fir. The composition of the present postfire/postcut forest differs from the prefire/precut forest given that it is dominated by sugar maple accompanied by yellow birch and white spruce.
When considering the floristic composition of both charcoal assemblages, mixed birch–balsam fir forests prevailed during the Holocene. Pollen data [45] also concluded on the persistence of the same tree species in the nearby Témiscouata (and Kamouraska) region where only minor vegetational changes occurred during the Holocene. The pollen curve of sugar maple west of the Rimouski area, in the Témiscouata uplands, shows the species occurrence early in the Holocene, at ca. 7200 cal. BP, if not earlier at 8500–9000 cal. BP [45]. It is probable that the species also arrived in the area south of Rimouski during the same period [46]. Were there any sugar maple forests in the study area during the long fire-free period between 5000 and 2000 cal. BP? Because of the lack of macrofossil data, it is not possible to assess the regional distribution of sugar maple forests despite favorable local and climatic conditions which most likely prevailed during this period. However, Richard et al. [45] suggest that sugar maple was more abundant during the mid-Holocene. Their conclusions on the development of the Appalachian vegetation, near our forest sites, are that forest communities were relatively stable during the Holocene, except during the initial phase of postglacial colonization. South of the study area, Elkadi [25] suggests a close relationship between the rise and expansion of sugar maple forests deduced from pollen and the reduced fire frequency (from microcharcoal) during the mid-Holocene.
According to the charcoal assemblages, sugar maple was a secondary, companion species, except at present. It is now the dominant species in the two study sites. At the Nicolas-Riou site, neither the conditions nor the moment of the shift of the millennial mixed birch–fir forest to the present sugar maple forest can be identified. Anthropogenic disturbances have not modified the course of forest succession in the past, explaining why the forest developed freely during the last 900 years. In this area where sugar maple forests are at their north-eastern range limit, the factors responsible for the balsam fir–sugar maple forest shift are unknown. However, demographic processes of the two tree species associated with canopy-gap dynamics may have played a structuring role in the alternating dominance of one species to the other with time. Both sugar maple and balsam fir are able to regenerate after fire and cutover in the Rimouski area according to surveys of stands regenerated after a 1932 fire [47]. Sugar maple and yellow birch colonize uphill sites after fire, whereas postfire balsam fir stands either accompanied by yellow birch or paper birch are distributed in mid-slope and down-slope sites [47]. In this study, however, there are no mentions of the tree composition of the forests burned in 1932, such that it is not possible to evaluate the resilience of the preburned stands, i.e., if the postfire stands are similar to the preburned stands.
The most important ecological changes at the Duchénier site also occurred during the last centuries, given that four repetitive fires over the last thousand years seem to have maintained the floristic composition of the site, a mixed balsam fir forest where sugar maple was a companion species. According to macrocharcoal produced by fire about 200 years ago (based on radiometric data of several charcoal fragments, and corresponding to the highest probability states, and tree-ring data [40]) the forest was dominated by conifers.
5. Conclusions
Soil macrocharcoal analysis at the stand scale [29,48,49,50] reveals local fire history from direct evidence. The results obtained from these in situ analyses allow the comparison of sites with different topographic and edaphic conditions at the Holocene time scale. Nonetheless, when multiple sites show a similar fire and tree composition history, a regional trend can then be extrapolated. Among the major findings of this study is the relatively similar fire history of the two sugar maple sites characterized by two main fire periods, in the early Holocene, from 9700 (9500) to 5000 cal. BP, and the late Holocene, and the long quiescent fire-free interval of more than 3500 cal. years separating the two fire periods. However, both sites differed in terms of fire occurrence during the last two thousand years. The Nicolas-Riou site was impacted by fire events about 900 years ago, which gives the current sugar maple forest an old-growth status maintained in the absence of fire disturbances over the last 900 cal. years. In contrast, the Duchénier site was affected by five fires during the last 2000 cal. years, and the current sugar maple stand is a young secondary forest established after fire and clear cut disturbances over the last 200 years. Based on scarce macrofossil evidence, both sugar maple sites were occupied by a relatively homogeneous forest during the course of the Holocene, except recently, with predominant mixed stands composed of conifer species, in particular balsam fir and spruce, and occasionally by eastern white cedar, and a few broadleaf species such as birch and sugar maple. The coexistence of conifer and broadleaf stands (balsam fir and sugar maple forests) was probably maintained in this area during a large part of the Holocene. The respective dominance of structuring species such as balsam fir and sugar maple throughout the Holocene of the Lower St. Lawrence region is attributable to natural and man-made disturbance events [17,19,44,45,47,51] which highlighted their life history traits as trans-successional tree species [22].
Acknowledgments
We would like to express our sincere thanks to Jocelyn Gosselin of the ‘Direction des inventaires forestiers’ of the Quebec Department of Forests, Fauna and Parks for initiating and funding this research on the Holocene fire history at the scale of southern Quebec. We are most grateful to Benoît Filion and Joanie Tremblay for field assistance. Special thanks to Francis St-Amour who participated in the long-lasting work of charcoal analysis. Dominique Arseneault from UQAR (Université du Québec à Rimouski) provided up-to-date information on recent forest surveys and forestry practices in the study area and the greater Rimouski area.
Author Contributions
S.P. conceived and designed the experiments; V.P., P.-L.C. and J.L. performed the experiments; V.P. and S.P. analyzed the data; S.P. and V.P. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Bonnicksen, T.M. America’s Ancient Forests: From the Ice Age to the Age of Discovery; John Wiley & Sons, Inc.: New York, NY, USA, 2000. [Google Scholar]
- Bureau, L. Les Solitudes de Lotbinière. Master’s Thesis, Université Laval, Québec, QC, Canada, 1968. [Google Scholar]
- Clark, J.S. Transformation of a northern hardwood forest by aboriginal (Iroquois) fire: Charcoal evidence from Crawford Lake, Ontario, Canada. Holocene 1995, 5, 1–9. [Google Scholar] [CrossRef]
- Fahey, T.J.; Reiners, W.A. Fire in the forests of Maine and New Hampshire. Bull. Torrey Bot. Club 1981, 108, 362–373. [Google Scholar] [CrossRef]
- Lorimer, C.G. The presettlement forest and natural disturbance cycle of northeastern Maine. Ecology 1977, 58, 139–148. [Google Scholar] [CrossRef]
- Lortie, M. Arbres, Forêts et Perturbations Naturelles au Québec; Les Presses de l’Université Laval: Québec, QC, Canada, 1979. [Google Scholar]
- Oliver, C.D.; Stephens, E.P. Reconstruction of a mixed-species forest in central New England. Ecology 1977, 58, 562–572. [Google Scholar] [CrossRef]
- Patterson, W.A.; Sassaman, K.E. Indian fires in the prehistory of New England. In Holocene Human Ecology in Northeastern North America; Nicholas, G., Ed.; Plenum: New York, NY, USA, 1988; pp. 107–135. [Google Scholar]
- Terrail, R. Influence de la Colonisation sur les Transformations du Paysage Forestier Depuis L’époque Préindustrielle dans l’Est du Québec. Ph.D. Thesis, UQAM, Montréal, QC, Canada, 2013. [Google Scholar]
- Clark, J.S.; Royall, P.D. Local and regional sediment charcoal evidence for fire regimes in presettlement north–eastern North America. J. Ecol. 1996, 84, 365–382. [Google Scholar] [CrossRef]
- Woods, K.D. Intermediate disturbance in a late-successional hemlock-northern hardwood forest. J. Ecol. 2004, 92, 464–476. [Google Scholar] [CrossRef]
- Grandtner, M.M. La Végétation Forestière du Québec Méridional; Les Presses de l’Université Laval: Québec, QC, Canada, 1966. [Google Scholar]
- Forcier, L.K. Reproductive strategies and the co-occurrence of climax tree species. Science 1975, 189, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Payette, S.; Filion, L.; Delwaide, A. Disturbance regime of a cold temperate forest as deduced from tree–ring patterns: The Tantaré Ecological Reserve, Quebec. Can. J. For. Res. 1990, 20, 1228–1241. [Google Scholar] [CrossRef]
- Runkle, J.R. Gap regeneration in some old-growth forest of the eastern United States. Ecology 1981, 62, 1041–1051. [Google Scholar] [CrossRef]
- Houle, G.; Payette, S. Seed dynamics of Abies balsamea and Acer saccharum in a deciduous forest of northeastern North America. Am. J. Bot. 1991, 78, 895–905. [Google Scholar] [CrossRef]
- Boucher, Y.; Arseneault, D.; Sirois, L. Logging-induced change (1930–2002) of a preindustrial landscape at the northern range limit of northern hardwoods, eastern Canada. Can. J. For. Res. 2006, 36, 505–517. [Google Scholar] [CrossRef]
- Brown, J.L. Les forêts du Témiscamingue, Québec: Ecologie et Photo-Interprétation; Laboratoire d’Écologie Forestière, Université Laval: Québec, QC, Canada, 1981. [Google Scholar]
- Dupuis, S.; Arseneault, D.; Sirois, L. Change from pre-settlement to present–day forest composition reconstructed from early land survey records in eastern Québec, Canada. J. Veg. Sci. 2011, 22, 564–575. [Google Scholar] [CrossRef]
- Brisson, J.; Bergeron, Y.; Bouchard, A. Les successions secondaires sur sites mésiques dans le Haut-Saint-Laurent, Québec, Canada. Can. J. Bot. 1988, 66, 1192–1203. [Google Scholar] [CrossRef]
- Bilodeau, J. Écologie des érablières à sucre (Acer saccharum Marsh.) à leur limite nord de distribution au Saguenay-Lac-Saint-Jean. Master’s Thesis, UQAC, Chicoutimi, QC, Canada, 1992. [Google Scholar]
- Nolet, P.; Delagrange, S.; Bouffard, D.; Doyon, F.; Forget, E. The successional status of sugar maple (Acer saccharum), revisited. Ann. For. Sci. 2008, 65, 208–217. [Google Scholar] [CrossRef]
- Pilon, V.; Payette, S. Sugar maple (Acer saccharum) forests at their northern distribution limit are recurrently impacted by fire. Can. J. For. Res. 2015, 45, 452–462. [Google Scholar] [CrossRef]
- Asnong, H.; Richard, P.J.H. La végétation et le climat postglaciaires du centre et de l’est de la Gaspésie, au Québec. Géogr. Phys. Quat. 2003, 57, 37–63. [Google Scholar] [CrossRef]
- Elkadi, T. L’histoire Postglaciaire de la Végétation et des Feux de la Région du Lac Mégantic. Master’s Thesis, Université de Montréal, Montréal, QC, Canada, 2013. [Google Scholar]
- Labelle, C.; Richard, P.J.H. Végétation tardiglaciaire et postglaciaire au sud-est du Parc des Laurentides. Géogr. Phys. Quat. 1981, 35, 345–359. [Google Scholar] [CrossRef]
- Talon, B.; Payette, S.; Filion, L.; Delwaide, A. Reconstruction of the long-term fire history of an old-growth deciduous forest in southern Québec, Canada, from charred wood in mineral soils. Quat. Res. 2005, 64, 36–43. [Google Scholar] [CrossRef]
- Frégeau, M.; Payette, S.; Grondin, P. Fire history of the central boreal forest in eastern North America reveals stability since the mid-Holocene. Holocene 2015, 25, 1912–1922. [Google Scholar] [CrossRef]
- Payette, S.; Delwaide, A.; Schaffhauser, A.; Magnan, G. Calculating long–term fire frequency at the stand scale from charcoal data. Ecosphere 2012, 3, 59. [Google Scholar] [CrossRef]
- Payette, S.; Pilon, V.; Couillard, P.L.; Frégeau, M. Long-term fire history of maple (Acer) forest sites in the central St. Lawrence Lowland, Quebec. Can. J. For. Res. 2016, 46, 822–831. [Google Scholar] [CrossRef]
- Arseneault, D.; (UQAR, Rimouski, Quebec, QC, Canada). Personal communication, 2013.
- Fonds de la compagnie Price Brothers, P666. Archives nationales du Québec à Saguenay: Saguenay, QC, Canada.
- Ohlson, M.; Tryterud, E. Interpretation of the charcoal record in forest soils: Forest fires and their production and deposition of macroscopic charcoal. Holocene 2000, 10, 519–525. [Google Scholar] [CrossRef]
- Hoadley, R.B. Identifying Wood: Accurate Results with Simple Tools; Taunton Press: Newtown, CT, USA, 1990. [Google Scholar]
- Inside Wood. Available online: http://insidewood.lib.ncsu.edu/search (accessed on 23 December 2016).
- Panshin, A.J.; de Zeeuw, C. Textbook of Wood Technology, 4th ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Stuiver, M.P.; Reimer, P.J.; Reimer, R.W. CALIB 7.02. (Computer Program and Documentation). 2013. Available online: http://radiocarbon.pa.qub.ac.uk/calib/ (accessed on 12 March 2014).
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Ramsey, C.B.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 radiocarbon age calibration curves 0–50000 years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef]
- Solanki, S.K.; Usoskin, I.G.; Kromer, B.; Schüssler, M.; Beer, J. Unusual activity of the sun during recent decades compared to the previous 11,000 years. Nature 2004, 431, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Pilon, V.; Payette, S.; Couillard, P.L.; Laflamme, J. Surface analysis as a tool to reconstruct the recent development of two contrasted sugar maple forests. 2017; Manuscript in preparation. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Soberon, J.; Llorente, J. The use of species accumulation functions for the prediction of species richness. Conserv. Biol. 1993, 7, 480–488. [Google Scholar] [CrossRef]
- Boucher, Y.; Arseneault, D.; Sirois, L. Logging-history (1820–2000) of a heavily exploited southern boreal forest landscape: Insights from sunken logs and forestry maps. For. Ecol. Manag. 2009, 258, 1359–1368. [Google Scholar] [CrossRef]
- Richard, P.J.H.; Larouche, A.C.; Lortie, G. Paléophytogéographie et paléoclimats postglaciaires dans l’ouest du Bas-Saint-Laurent, Québec. Géogr. Phys. Quat. 1992, 46, 151–172. [Google Scholar] [CrossRef]
- Richard, P.J.H.; Larouche, A.C. Histoire postglaciaire de la végétation et du climat dans la région de Rimouski, Québec. In Il y a 8000 ans à Rimouski, Paléoécologie et Archéologie d’un Site de la Culture Plano; Chapdelaine, C., Ed.; Collection Paléo-Québec No. 22; Recherches amérindiennes au Québec: Montréal, QC, Canada, 1994; pp. 49–89. [Google Scholar]
- Archambault, L.; Morissette, J.; Bernier-Cardou, M. Succession forestière après feu dans la sapinière à bouleau jaune du Bas-Saint-Laurent, Québec. For. Chron. 1997, 73, 702–710. [Google Scholar] [CrossRef]
- Fesenmyer, K.A.; Christensen, N.L., Jr. Reconstructing Holocene fire history in a southern Appalachian forest using soil charcoal. Ecology 2010, 91, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.L.; Horn, S.P.; Grissino-Mayer, H.D. Fire history from soil charcoal in a mixed hardwood forest in the Cumberland Plateau, Tennessee, USA. J. Torrey Bot. Soc. 2008, 135, 401–410. [Google Scholar] [CrossRef]
- Touflan, P.; Talon, B.; Walsh, K. Soil charcoal analysis: A reliable tool for spatially precise studies of past forest dynamics: A case study in the French Southern Alps. Holocene 2010, 20, 45–52. [Google Scholar] [CrossRef]
- Sorel, C. Impacts des Perturbations Anthropiques du XXe Siècle sur Deux Forêts du Bas-Saint-Laurent (Québec). Master’s Thesis, UQAR, Rimouski, QC, Canada, 2004. [Google Scholar]
Figure 1.
Location of SNR (Nicolas-Riou Seigneury) and RFD (Duchénier Faunal Reserve) sugar maple sites (red dots), and the other Rivière-Éternité (RIÉ) [23], Villeroy (VIL) and Ste–Françoise (STF) [30] sugar maple sites (red squares).
Figure 2.
Distribution of the sum of cumulated probabilities of calibrated 14C charcoal dates for the Nicolas-Riou Seigneury site (a) and the Duchénier Faunal Reserve site (b). The sum of probabilities in gray shading corresponds to the time intervals (with the highest probability) of fire dates. Each x symbol indicates the occurrence of a fire event centered on the mean of the greatest probability interval of one or several radiocarbon dates. The number of superposed x symbols between 1400 and 900 cal. BP in (a) correspond to minimum (4x symbols) and maximum (6x symbols) fires as deduced from the probability distribution of cal. years and range of overlapping cal. years.
Figure 2.
Distribution of the sum of cumulated probabilities of calibrated 14C charcoal dates for the Nicolas-Riou Seigneury site (a) and the Duchénier Faunal Reserve site (b). The sum of probabilities in gray shading corresponds to the time intervals (with the highest probability) of fire dates. Each x symbol indicates the occurrence of a fire event centered on the mean of the greatest probability interval of one or several radiocarbon dates. The number of superposed x symbols between 1400 and 900 cal. BP in (a) correspond to minimum (4x symbols) and maximum (6x symbols) fires as deduced from the probability distribution of cal. years and range of overlapping cal. years.
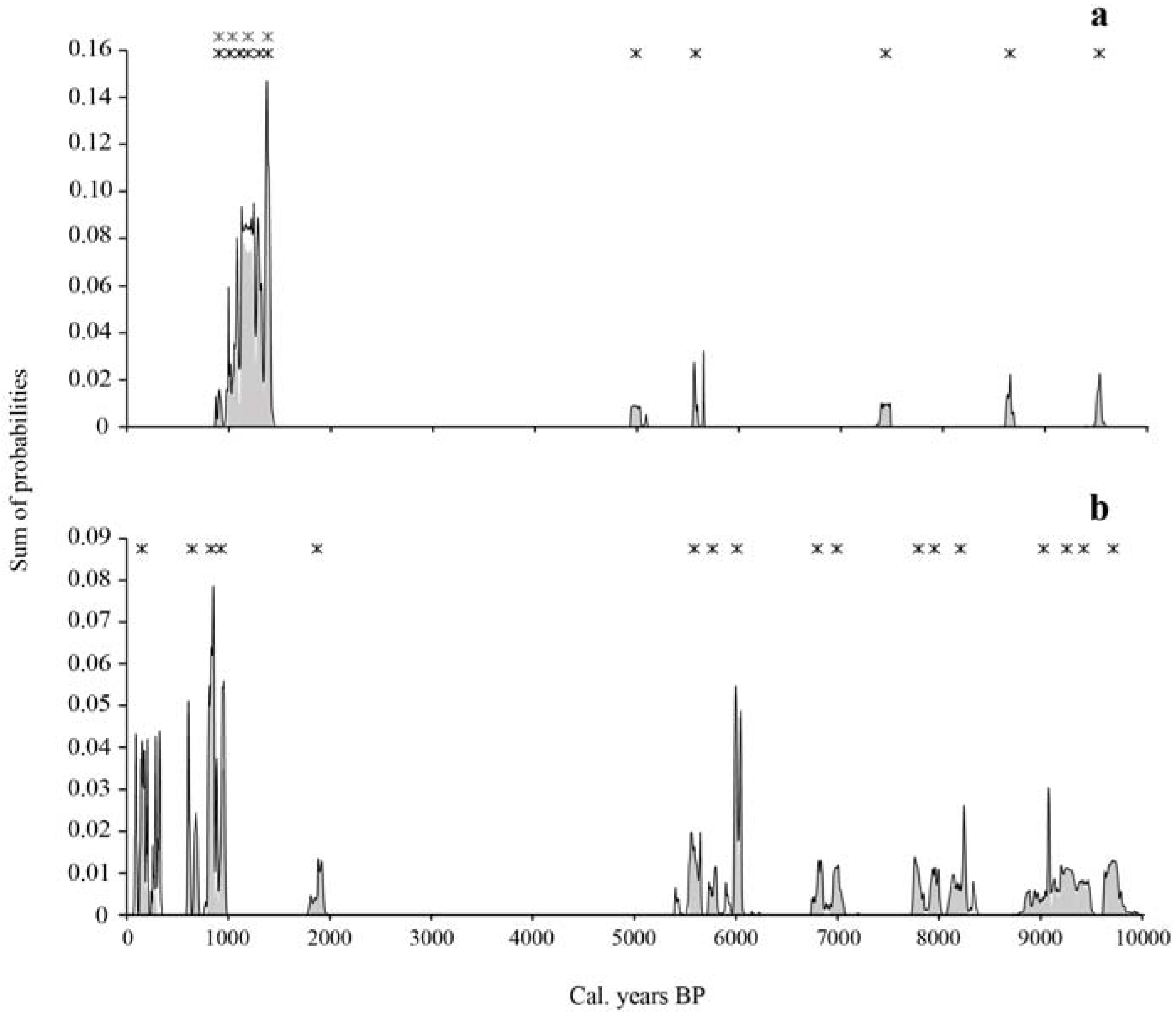
Figure 3.
Average accumulation curve of the number of macrocharcoal pieces dated and fires detected (mean of 100 iterations ± σ) for the SNR (a) and RFD (b) sites. The number of fires detected F(n), the estimated number of fires F(max) and the constant k are also shown.
Figure 3.
Average accumulation curve of the number of macrocharcoal pieces dated and fires detected (mean of 100 iterations ± σ) for the SNR (a) and RFD (b) sites. The number of fires detected F(n), the estimated number of fires F(max) and the constant k are also shown.
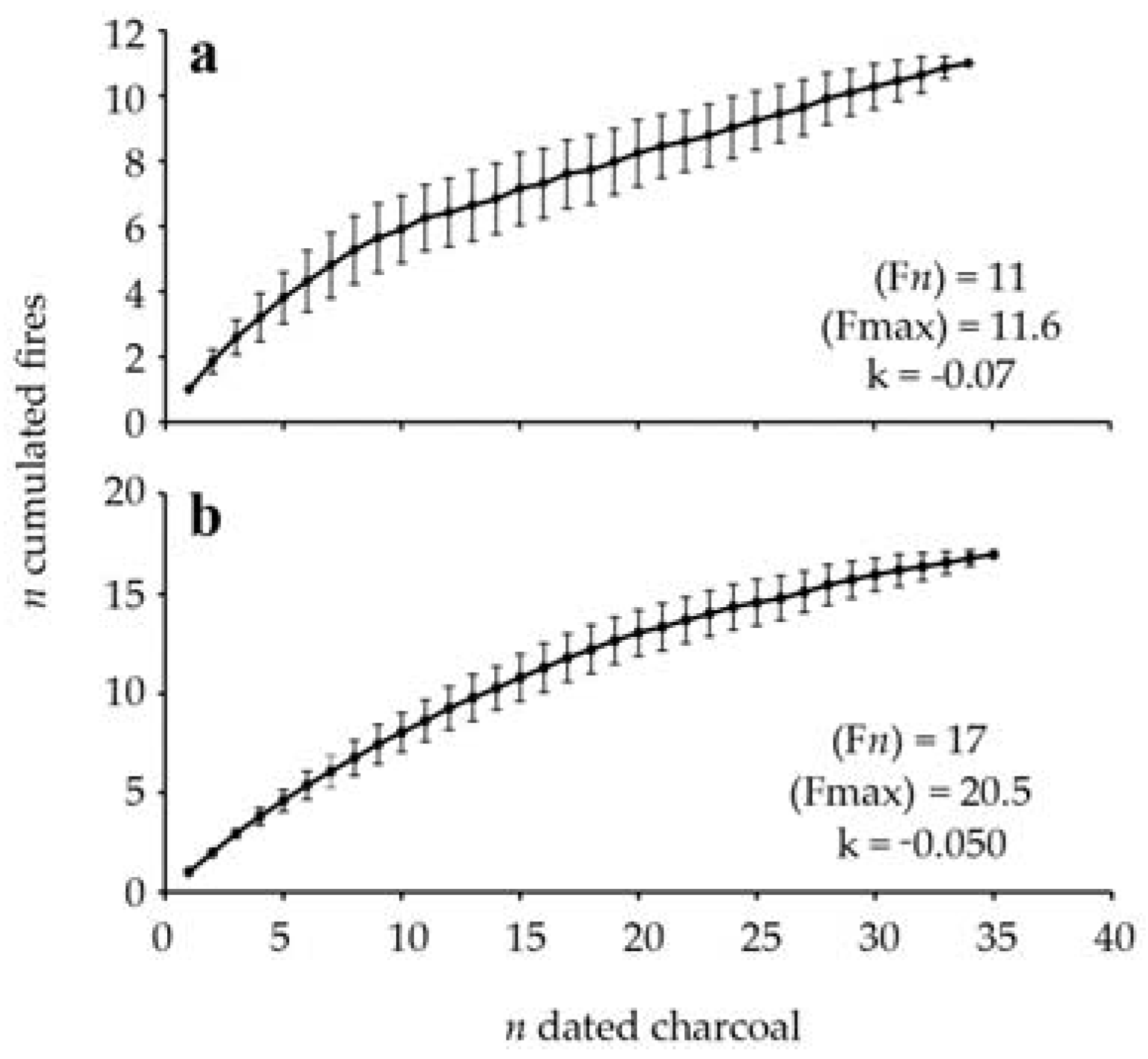
Figure 4.
Occurrence of fires during the Holocene at Villeroy (VIL), Ste-Françoise (STF), Rivière-Éternité (RIÉ), Nicolas-Riou (SNR) and Duchénier (RFD) sites, and cumulated fires (%) of all sites. The total number of fires detected is indicated under each site name abbreviation. Results are presented in percentages of fire that occurred per 1000 cal.-year period. Sources: VIL and STF, [30]; RIÉ, [23].
Figure 4.
Occurrence of fires during the Holocene at Villeroy (VIL), Ste-Françoise (STF), Rivière-Éternité (RIÉ), Nicolas-Riou (SNR) and Duchénier (RFD) sites, and cumulated fires (%) of all sites. The total number of fires detected is indicated under each site name abbreviation. Results are presented in percentages of fire that occurred per 1000 cal.-year period. Sources: VIL and STF, [30]; RIÉ, [23].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Principal species growing in the current sites of the Nicolas-Riou Seigneury (SNR) and the Duchénier Faunal Reserve (RFD) sugar maple forests. The “x” symbol indicates the presence of the species. The entire surface of both 0.1 ha plots was surveyed. Species that were noticed outside the plots but in the same stand were also recorded as present.
Table 1.
Principal species growing in the current sites of the Nicolas-Riou Seigneury (SNR) and the Duchénier Faunal Reserve (RFD) sugar maple forests. The “x” symbol indicates the presence of the species. The entire surface of both 0.1 ha plots was surveyed. Species that were noticed outside the plots but in the same stand were also recorded as present.
| Layer | Species | SNR | RFD |
|---|---|---|---|
| Tree | Abies balsamea (L.) Mill, | x | x |
| Acer rubrum L. | 1 tree | ||
| Acer saccharum Marsh. | x | x | |
| Betula alleghaniensis Britton | x | x | |
| Betula papyrifera Marsh. | x | ||
| Picea glauca (Moench) Voss | x | x | |
| Pinus strobus L. | 1 seedling | ||
| Thuja occidentalis L. | x | x | |
| Shrub | Acer pensylvanicum L. | x | x |
| Acer spicatum Lam. | x | ||
| Corylus cornuta Marsh. | x | x | |
| Lonicera canadensis Bartr. ex Marsh. | x | ||
| Rubus idaeus L. | x | ||
| Sorbus sp. | seedlings | ||
| Taxus canadensis Marsh, | x | ||
| Herb | Actaea rubra (Aiton) Willd. | x | |
| Aralia nudicaulis L. | x | x | |
| Carex sp. | 1 individual | ||
| Circaea alpina L. | x | ||
| Dryopteris carthusiana (Vill.) H.P. Fuchs | x | x | |
| Galium sp. | x | ||
| Gymnocarpium disjunctum (Rupr.) Ching | x | ||
| Huperzia lucidula (Michaux) Trevisan | x | ||
| Hypopitys monotropa Crantz | x | x | |
| Lysimachia borealis (Raf.) U. Manns & Anderb. | x | x | |
| Mitella nuda L. | x | ||
| Osmundastrum sp. | x | ||
| Oxalis montana Raf. | x | ||
| Phegopteris connectilis (Michx.) Watt | x | x | |
| Rubus pubescens Raf. | x | ||
| Streptopus lanceolatus (Ait.) Reveal | x | x | |
| Trillium erectum L. | x | x |
Table 2.
Number, average weight, and identification of the charcoal pieces of the Nicolas-Riou (SNR) and the Duchénier (RFD) sugar maple forests. The number of microsites that contained charcoal of a given group either in the mineral core and/or in the organic monolith, is indicated in the n microsite column. Sixteen and 24 of the 25 microsites sampled contained charcoal at SNR and RFD, respectively.
Table 2.
Number, average weight, and identification of the charcoal pieces of the Nicolas-Riou (SNR) and the Duchénier (RFD) sugar maple forests. The number of microsites that contained charcoal of a given group either in the mineral core and/or in the organic monolith, is indicated in the n microsite column. Sixteen and 24 of the 25 microsites sampled contained charcoal at SNR and RFD, respectively.
| Charcoal Pieces | SNR | RFD | ||||
|---|---|---|---|---|---|---|
| Organic | Mineral a | n Microsite | Organic | Mineral | n Microsite | |
| Average weight (±σ) mg | 9.2 (±15.3) | 8.8 (±10.9) | ||||
| n extracted charcoal pieces | >800 | 211 | >1000 | 39 | ||
| Unidentified conifer | 48 | 27 | 14 | 133 | 9 | 17 |
| Picea sp. and Picea sp./Larix laricina | 2 | 10 | 5 | 75 | 4 | 8 |
| Abies balsamea/Thuja occidentalis b | 18 | 10 | 4 | 87 | 3 | 9 |
| Abies balsamea c | 13 | 2 | 8 | 45 | 2 | 7 |
| Thuja occidentalis | 6 | 1 | 2 | |||
| Taxus canadensis | 3 | 2 | ||||
| Unidentified broadleaf | 52 | 35 | 11 | 22 | 11 | |
| Betula sp. | 60 | 35 | 5 | 62 | 1 | 11 |
| Betulaceae | 13 | 4 | 3 | 3 | 2 | |
| Betulaceae shrub | 8 | 1 | 1 | 8 | 5 | 5 |
| Other shrub | 2 | 1 | ||||
| Broadleaf d | 15 | 2 | 3 | 13 | 2 | |
| Acer sp. | 3 | 3 | 1 | 19 | 1 | |
| Acer saccharum | 33 | 1 | 2 | 1 | 2 | |
| Populus sp./Salix sp. | 2 | 1 | ||||
| Unidentified pieces | 16 | 26 | 12 | 12 | 10 | 12 |
| Fungi | 10 | 3 | 33 | 11 | ||
| Leaf-bud | 2 | 2 | ||||
| Bark-pith | 40 | 29 | 9 | 79 | 3 | 15 |
a Including 21 unidentified pieces <2.5 mg; b Including Abies-Thuja-Tsuga; c Including Abies-Tsuga; d With spiral thickenings in vessel elements, including lamellar.
Table 3.
14C dates, calibrated years (cal. years before 2015), botanical identification of charcoal fragments and number of fire events recorded in the Nicolas-Riou site.
Table 3.
14C dates, calibrated years (cal. years before 2015), botanical identification of charcoal fragments and number of fire events recorded in the Nicolas-Riou site.
| Microsite | Location | Sample Number a | Taxon or Piece | 14C Age (±σ) | Cal. Age | Probability | n Fires Cumulated |
|---|---|---|---|---|---|---|---|
| 14 | Mineral/organic contact | C-157758 | Picea sp./Larix laricina | 955 ± 15 | 865–935 | 0.69 | 11 |
| 6 | Mineral A | C-147457 | Conifer | 1015 ± 15 | 985–1025 | 1.00 | 10 |
| 14 | Organic | G-21386 | Abies balsamea/Thuja occidentalis | 1085 ± 20 | 1000–1075 | 0.70 | 9/10 |
| 17 | Mineral A | C-147888 | Acer sp. | 1135 ± 20 | 1035–1145 | 0.98 | 9 |
| 23 | Mineral A | C-147456 | Conifer | 1140 ± 15 | 1045–1140 | 0.99 | |
| 24 | Mineral/organic contact | C-157757 | Bark | 1155 ± 15 | 1110–1160 | 0.43 | 8/9 |
| 20 | Mineral A | C-147918 | Betulaceae, shrub | 1155 ± 20 | 1110–1215 | 0.58 | |
| 6 | Mineral B | C-147920 | Taxus canadensis | 1160 ± 20 | 1115–1240 | 0.79 | 8 |
| 4 | Mineral B | G-18880 | Conifer, vitrified | 1170 ± 20 | 1120–1240 | 0.93 | |
| 16 | Mineral/organic contact | C-157769 | cf. Acer saccharum | 1220 ± 15 | 1135–1230 | 0.73 | |
| 20 | Mineral/organic contact | C-157770 | cf. Corylus cornuta | 1190 ± 20 | 1130–1240 | 1.00 | |
| 24 | Mineral B | C-147919 | Structure without xylem /bark | 1190 ± 20 | 1130–1240 | 1.00 | |
| 24 | Mineral/organic contact | G-21403 | Picea sp./Larix laricina | 1200 ± 20 | 1130–1245 | 1.00 | |
| 16 | Organic | G-21405 | Abies balsamea (Tsuga) b | 1215 ± 20 | 1130–1250 | 0.92 | |
| 23 | Mineral B | C-147453 | Picea sp./Larix laricina | 1200 ± 15 | 1135–1240 | 1.00 | |
| 15 | Mineral A | C-147922 | Abies balsamea (Tsuga) b | 1210 ± 20 | 1130–1245 | 0.96 | |
| 13 | Mineral A | C-147887 | Abies balsamea | 1245 ± 20 | 1235–1330 | 0.87 | |
| 14 | Mineral A | G-18884 | Abies balsamea/Thuja occidentalis | 1250 ± 20 | 1240–1335 | 0.94 | 7 |
| 10 | Mineral B | C-147455 | Picea sp./Larix laricina | 1265 ± 15 | 1245–1330 | 1.00 | |
| 24 | Mineral A | C-147917 | Betula sp. | 1275 ± 20 | 1245–1340 | 1.00 | |
| 4 | Mineral A | C-147466 | Vitrified | 1235 ± 15 | 1270–1325 | 0.53 | |
| 24 | Mineral/organic contact | C-157771 | Betula sp. | 1275 ± 15 | 1285–1335 | 0.59 | |
| 11 | Mineral A | C-147916 | Abies balsamea (Tsuga) b | 1355 ± 20 | 1335–1370 | 1.00 | 6/7 |
| 18 | Organic | C-157761 | Betula sp. | 1385 ± 20 | 1350–1395 | 1.00 | |
| 18 | Organic | C-157760 | Betula sp. | 1410 ± 20 | 1355–1410 | 1.00 | 6 |
| 18 | Mineral A | G-18882 | Vitrified | 1420 ± 20 | 1360–1415 | 1.00 | |
| 18 | Mineral B | C-147454 | Betula sp. | 1425 ± 20 | 1360–1415 | 1.00 | |
| 11 | Mineral A | C-147465 | Vitrified | 1430 ± 15 | 1365–1415 | 1.00 | |
| 18 | Mineral/organic contact | G-21402 | Betula sp. | 1435 ± 20 | 1365–1425 | 1.00 | |
| 2 | Mineral A | C-147459 | Broadleaf | 4380 ± 20 | 4930–5040 | 0.92 | 5 |
| 20 | Mineral A | C-147458 | Conifer | 4810 ± 20 | 5545–5600 | 0.71 | 4 |
| 14 | Mineral A | C-147461 | Broadleaf (not Acer sp.) | 6430 ± 20 | 7375–7485 | 1.00 | 3 |
| 12 | Mineral B | C-147923 | Picea sp. | 7810 ± 25 | 8610–8695 | 1.00 | 2 |
| 12 | Mineral A | C-147460 | Conifer, vitrified | 8425 ± 25 | 9490–9570 | 0.99 | 1 |
a C:UCIAMS; G:UGAMS. b Piece with anatomical traits corresponding to fir and hemlock (Tsuga canadensis). Hemlock is absent from the Lower St. Lawrence region.
Table 4.
14C dates, calibrated years (cal. years before 2015), botanical identification of charcoal fragments and number of fire events recorded in the Duchénier site.
Table 4.
14C dates, calibrated years (cal. years before 2015), botanical identification of charcoal fragments and number of fire events recorded in the Duchénier site.
| Microsite | Location | Sample Number a | Taxon or Piece | 14C Age (±σ) | Cal. Age | Probability | n Fires Cumulated |
|---|---|---|---|---|---|---|---|
| 3 | Mineral/organic contact | C-157767 | Picea sp. | 130 ± 15 | 125–185 | 0.39 | 17 |
| 3 | Mineral/organic contact | C-157751 | Bark | 130 ± 15 | 125–185 | 0.39 | |
| 3 | Organic | C-157768 | cf. Thuja occidentalis | 130 ± 15 | 125–185 | 0.39 | |
| 3 | Organic | C-157755 | Conifer | 115 ± 15 | 125–205 | 0.61 | |
| 3 | Organic | G-21385 | Bark | 140 ± 20 | 235–295 | 0.29 | |
| 3 | Mineral/organic contact | C-157754 | Broadleaf with spiral thickenings, lamellar | 545 ± 15 | 590–620 | 0.83 | 16 |
| 3 | Organic | C-157756 | Acer saccharum, lamellar | 595 ± 15 | 650–710 | 0.77 | |
| 14 | Mineral B | C-147925 | Thuja occidentalis | 855 ± 20 | 790–860 | 0.98 | 15 |
| 17 | Mineral A | C-148767 | Bark | 860 ± 20 | 790–860 | 0.98 | |
| 18 | Mineral B | C-148224 | Abies balsamea | 880 ± 20 | 800–865 | 0.77 | |
| 15 | Organic | C-157751 | cf. Abies balsamea | 890 ± 15 | 805–865 | 0.60 | |
| 21 | Organic | C-157766 | Betula sp., branch | 910 ± 20 | 845–980 | 0.99 | 14 |
| 15 | Mineral/organic contact | C-157750 | cf. Corylus cornuta | 905 ± 20 | 905–975 | 0.55 | |
| 23 | Mineral A | G-18883 | cf. Betulaceae, shrub | 905 ± 20 | 905–975 | 0.55 | |
| 11 | Mineral A | C-148765 | Conifer, altered | 1880 ± 25 | 1800–1945 | 1.00 | 13 |
| 5 | Mineral A | G-18876 | Acer saccharum | 4800 ± 25 | 5540–5610 | 0.80 | 12 |
| 4 | Mineral A | C-148770 | Vitrified, orange deposit | 4740 ± 25 | 5565–5650 | 0.61 | |
| 19 | Mineral A | C-148768 | Structure without xylem | 5005 ± 25 | 5720–5820 | 0.75 | 11 |
| 12 | Mineral B | C-148766 | Conifer, altered | 5185 ± 25 | 5975–6020 | 0.68 | 10 |
| 9 | Mineral A | C-148772 | Vitrified | 5200 ± 30 | 5975–6060 | 1.00 | |
| 2 | Mineral A | G-18885 | Abies balsamea (Tsuga) b | 5200 ± 25 | 5980–6055 | 1.00 | |
| 15 | Mineral A | C-148771 | Vitrified, orange deposit | 5935 ± 25 | 6740–6865 | 0.95 | 9 |
| 9 | Mineral A | G-18877 | Betula sp. | 6070 ± 25 | 6925–7065 | 1.00 | 8 |
| 23 | Mineral B | C-147924 | cf. Betulaceae shrub | 6895 ± 25 | 7740–7850 | 1.00 | 7 |
| 8 | Mineral B | C-148730 | Conifer, altered | 7050 ± 30 | 7895–8020 | 1.00 | 6 |
| 3 | Mineral B | C-148769 | Vitrified, orange deposit | 7310 ± 40 | 8085–8250 | 1.00 | 5 |
| 19 | Mineral A | G-18879 | Conifer, reaction wood | 7370 ± 25 | 8220–8290 | 0.66 | |
| 5 | Mineral A | G-18878 | Picea sp. | 7995 ± 25 | 8835–9065 | 1.00 | 4 |
| 13 | Mineral A | G-18881 | Vitrified, orange deposit | 8100 ± 25 | 9060–9155 | 1.00 | 3/4 |
| 22 | Mineral A | C-147926 | cf. Betulaceae shrub | 8210 ± 25 | 9145–9340 | 0.96 | 3 |
| 4 | Mineral A | C-148728 | Picea sp. | 8240 ± 35 | 9150–9380 | 0.94 | |
| 5 | Mineral B | C-147928 | Picea sp. | 8330 ± 25 | 9350–9500 | 1.00 | 2 |
| 6 | Mineral B | C-148729 | Conifer | 8690 ± 35 | 9610–9775 | 0.97 | 1 |
| 22 | Mineral A | G-18875 | Vitrified | 8720 ± 30 | 9615–9775 | 0.99 |
a C:UCIAMS; G:UGAMS; b Charcoal whose anatomical characters correspond to balsam fir or hemlock (Tsuga canadensis). Hemlock is absent from the Lower St. Lawrence region.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Payette, S.; Pilon, V.; Couillard, P.-L.; Laflamme, J. Fire History of Appalachian Forests of the Lower St-Lawrence Region (Southern Quebec). Forests 2017, 8, 120. https://doi.org/10.3390/f8040120
AMA Style
Payette S, Pilon V, Couillard P-L, Laflamme J. Fire History of Appalachian Forests of the Lower St-Lawrence Region (Southern Quebec). Forests. 2017; 8(4):120. https://doi.org/10.3390/f8040120
Chicago/Turabian StylePayette, Serge, Vanessa Pilon, Pierre-Luc Couillard, and Jason Laflamme. 2017. "Fire History of Appalachian Forests of the Lower St-Lawrence Region (Southern Quebec)" Forests 8, no. 4: 120. https://doi.org/10.3390/f8040120
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.