Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA


1
Department of Natural Resource Ecology and Management, Oklahoma State University, 008 Agricultural Hall, Stillwater, OK 74078, USA
2
Bureau of Indian Affairs, Southern Plains Region, P.O. Box 368, Anadarko, OK 73005, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(2), 75; https://doi.org/10.3390/f9020075
Submission received: 8 January 2018
/
Revised: 29 January 2018
/
Accepted: 1 February 2018
/
Published: 3 February 2018
(This article belongs to the Special Issue Ecology and Management of Invasive Species in Forest Ecosystems)
Abstract
:Cross Timbers forests, typically dominated by Quercus stellata Wangenh. and Q. marilandica Muenchh., are the transition zone between eastern deciduous forest and prairie in the southern Great Plains. Fire exclusion beginning in the mid-1900s has led to increasing stand density and encroachment of fire-intolerant Juniperus virginiana L. and mesic hardwood. We measured current forest structure and tree ages of 25 stands (130 plots) in north-central Oklahoma to characterize the extent and dynamics of encroachment. The respective basal area and stand density of the overstory (diameter at breast height; dbh > 10 cm) were 19.0 m2 ha−1 and 407 trees ha−1 with Q. stellata comprising 43% of basal area and 42% of stand density. Quercus marilandica represented only 3% of basal area and 4% of overstory density. Juniperus virginiana represented 7% of basal area and 14% of stand density while mesic hardwoods, e.g., Celtis spp., Ulmus spp., Carya spp., 33% of basal area and stand density. The sapling layer was dominated by mesic hardwoods (68%) and J. virginiana (25%) while the seedling layer was dominated by mesic hardwoods (74%). The majority of Quercus recruited into the overstory between 1910–1970, while recruitment of J. virginiana and mesic hardwoods began more recently (post 1950s). Growth rate, based on the relationship between age and dbh, was faster for mesic hardwoods than for J. virginiana and Q. stellata. These results indicate that removal of recurrent surface fire as a disturbance agent has significantly altered forest composition in the Cross Timbers region by allowing encroachment of J. virginiana and fire-intolerant, mesic hardwoods. This increases wildfire risk because J. virginiana is very flammable and will alter how these forests respond to future drought and other disturbance events.
1. Introduction
The Cross Timbers region of the United States is the ecotone between the eastern deciduous forest and the southern Great Plains, stretching from Texas north through Oklahoma into southern Kansas. This region is a mosaic comprising oak forest, oak savanna, and tallgrass prairie. The Cross Timbers region historically covered approximately 8 million hectares, about 4.5 million in Oklahoma [1]. The forest areas in the Cross Timbers are typically dominated by an overstory of Quercus stellata (post oak) and Q. marilandica (blackjack oak) that reach approximately 15 m in height [2].
The species composition and structure of Cross Timbers forests were maintained by frequent surface fire [3]. Measurements of the historical fire return interval pre-European American settlement ranged from 2.9–6.7 years [4,5,6,7,8]. Due to a combination of fire exclusion and favorable recruitment following canopy gaps that developed as a result of widespread drought in the 1950s, the Cross Timbers forests have seen an increase in stand basal area and an increase in fire-intolerant species such as Juniperus virginiana (eastern redcedar), Celtis spp., and Ulmus spp. [9]. This change in forest composition and structure in the Cross Timbers parallels the “mesophication” of much of the rest of the eastern deciduous forest, where fire-tolerant oak species are replaced with less fire-tolerant species due to fire exclusion [10]. In particular, the encroachment of highly flammable, evergreen J. virginiana into Cross Timbers forests is of serious concern. Re-measurement of stands initially sampled from 1953–1957 by Rice and Penfound [11] indicated that J. virginiana increased from 0.05 to 2.71 m2 ha−1 in terms of basal area and from 0.73 to 23.85 stems ha−1, while the overall stand basal area and stand density roughly doubled [9,12].
Juniperus virginiana is typically considered a shade-intolerant species; however, within the forest interior of the Cross Timbers, J. virginiana finds enough light availability in canopy gaps and is physiologically active during the winter/spring oak leaf-off period [13]. Once J. virginiana establishes, it shades out herbaceous vegetation [14] and adds litter to the forest floor that decomposes differently from oak leaves [15], alters fungal communities and litter chemistry [16], and burns much less readily than either oak litter or herbaceous vegetation. This reduces the available fine fuel load around J. virginiana trees and saplings and results in a forest that is incapable of maintaining a consistent, low-intensity surface fire. These low-intensity surface fires are necessary to purge J. virginiana seedlings (and other fire intolerant species), which become difficult to kill with prescribed fire once the trees reach a height of approximately 2 m [17,18].
In contrast to its effects on surface fire, J. virginiana encroachment increases the risk of wildfire. Nowacki and Abrams [10] proposed an alternative stable state for the oak forests of the eastern United States where fire exclusion leads to a more mesic forest that is resistant to all types of fire due to reduced fine fuel loading. However, the encroaching J. virginiana is more xeric, resinous, and burns extremely well under drought/wildfire conditions [19]. Hoff et al. [20] found that J. virginiana encroachment in north-central Oklahoma increased available fuel loading for wildfires by 38% through the inclusion of J. virginiana foliage and small branches. This reduction in ability for fine fuels to support a surface fire combined with the increased risk of crown fire from J. virginiana in the midstory sets the stage for wildfires that will be harder to suppress and more dangerous to human health and property. Increased risk and severity of wildfires also may change the regeneration pattern of Cross Timbers forests by replacing surface fires with stand-replacing fires [21].
Understanding the extent of encroachment of fire-intolerant species into Cross Timbers forests and the timeline of the invasion process is relevant to understanding regional forest dynamics and is important information for land managers. Previous work by Stambaugh et al. [21] and DeSantis et al. [9], based on tree aging, documented an increase in J. virginiana within Cross Timbers forests beginning in the mid 1900s. However, neither study looked at the timeline for invasion of more mesic species such as Celtis occidentalis L., Celtis laevigta Willdenow, or Ulmus americana L. In addition, the majority of studies on Cross Timbers forests have occurred on government controlled and managed lands (e.g., [4,5,7,8,22]) that do not reflect the typical disturbance pattern for land owned by small private landowners, which represents the vast majority of land holdings in the region. By knowing the dynamics of J. virginiana and mesic species invasion into Cross Timbers forests, land managers can prioritize and schedule management activities based on current conditions and projected rates of encroachment to reduce J. virginiana, reduce wildfire risk, and/or restore historical ecosystem composition or structure.
To address the issue of J. virginiana and mesic species encroachment into Cross Timbers forests, we measured current forest structure and condition during the summer of 2016 on Bureau of Indian Affairs (BIA) properties in north-central Oklahoma. In addition, we determined the age structure of the various species and related that to the dynamics of J. virginiana and mesic hardwood encroachment. Our objectives were to (1) quantify the current forest overstory, sapling, and seedling composition, (2) determine the extent of encroachment by fire-intolerant species, (3) establish a sub-decadal timeline of encroachment, and (4) compare growth rates of major species groups.
2. Materials and Methods
2.1. Study Area
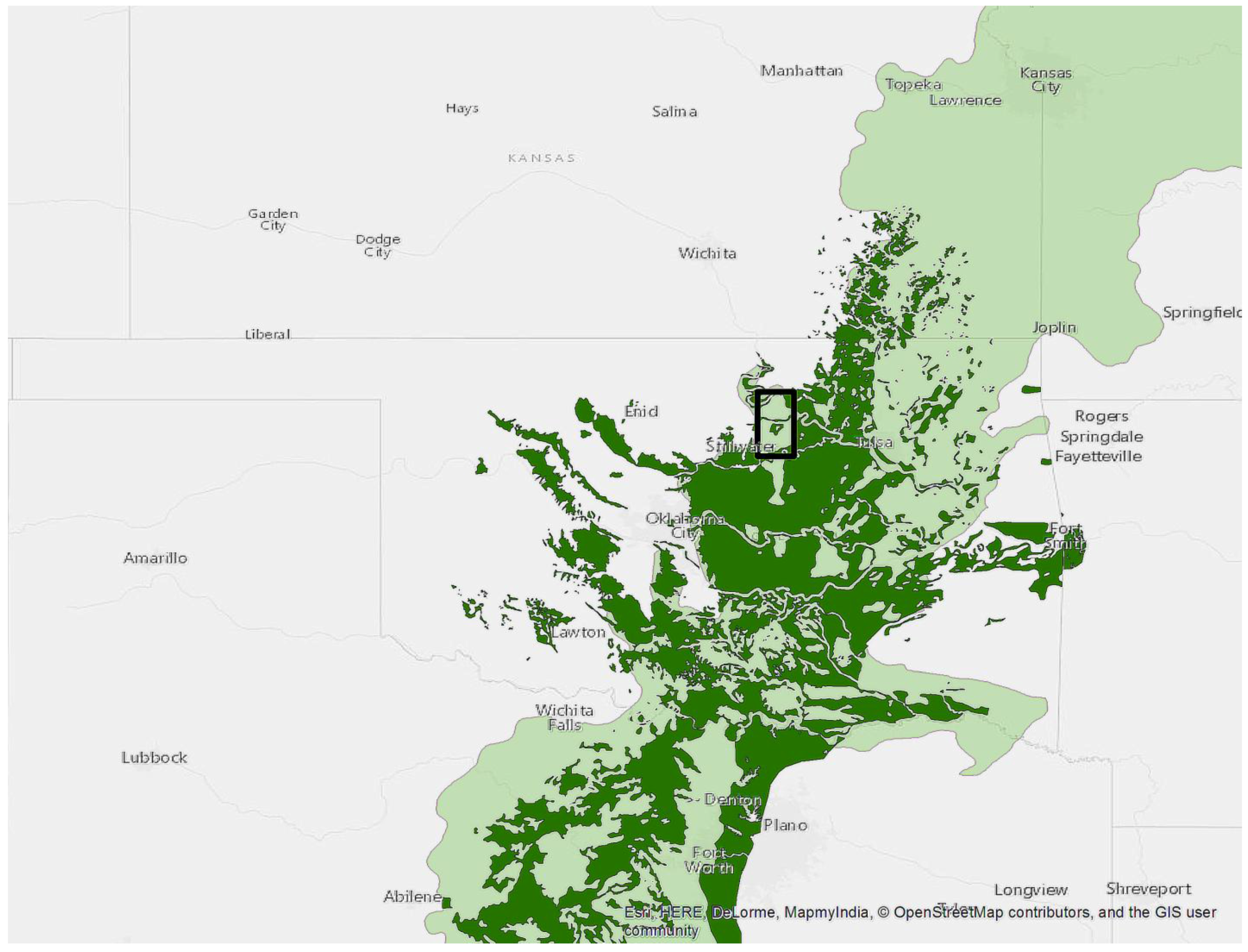
We used 25 properties managed in trust for the Pawnee Tribe by the BIA, Southern Plains Region (Pawnee and Payne Counties Oklahoma, USA). These are typically landholdings ~65 ha in size for which the federal government holds legal title but the beneficial interest and management remain with the individual tribal member. We did not identify recent fires on any of the 25 properties. This subset of the BIA managed properties in the region (Figure 1) was randomly selected and had to meet two criteria; they had to be in excess of 16 ha and have approximately 50% oak forest canopy cover during leaf-on conditions. These 25 properties fell on a north-south swath from the Arkansas River in the north to the Cimarron River in the south (Figure 2). The eastern extent was limited by the availability of BIA properties in the Southern Plains region while the western extent was limited by the 50% oak forest requirement as Cross Timbers forests transitions into prairie. This area in Oklahoma averages 100 cm of annual precipitation. The average daytime high temperature is 15.5 °C, ranging from 34.0 °C in July to 8.8 °C in January. The growing season averages 197 days. Winds from the south and southeast dominate, averaging 14.5 km hr−1 (OK Climatological Survey 2016). Soils are predominantly silt loam complexes and feature prominent interspersed rock outcroppings [23].
Once the forest areas were identified on each of the 25 properties [20], five field measurement plots per property (one larger property had ten field measurement plots) were randomly located within the forest interior using ArcMAP (Esri, Redlands, CA, USA). Plot locations were found in the field using the onboard GPS capability of an iPad Air (Apple, Cupertino, CA, USA) with an accuracy of roughly 5 m. Field plot centers were marked with a metal stake and tagged. Plots were visited from May to August 2016, and data were collected using modified FIREMON protocol [25] designed for long-term fire effects monitoring.
2.2. Data Collection
Overstory hardwood tree data were collected using a circular fixed radius plot of 11.3 m (0.04 ha). All overstory trees, defined as having a diameter at breast height (dbh) greater than 10 cm, were measured 1.37 m above the ground to the nearest 0.5 cm and identified by species. On every 5th tree, starting with the most dominant tree (determined by relative size and crown dominance) on the plot and moving clockwise, we measured tree height to the nearest 0.1 m using a laser hypsometer (Truepulse 200b, Laser Technology Inc., Centennial, CO, USA) or a telescopic measuring rod (Hastings, Hastings, MI, USA). Cores (5.15 mm diameter) were taken at breast height using an increment corer (Haglof, Långsele, Sweden) on this subset of trees. We cored a minimum of three overstory hardwood trees per plot. If a plot had an insufficient number of mature stems to sample three stems by choosing every fifth, an alternative sampling strategy was used. For example, a plot with nine trees would have every third tree sampled and for a plot with only three trees, all were sampled.
Hardwood sapling data were collected within a smaller fixed radius subplot of 3.57 m (0.004 ha). Saplings were defined by a dbh between 0.1 and 10 cm. We identified the species of each stem, measured height to the nearest 0.1 m and recorded dbh as one of five size classes, i.e., 0–2, 2–4, 4–6, 6–8, 8–10 cm. We quantified seedling density and size within a subplot with radius of 1.78 m (0.001 ha) by recording a count by species and height class. Height class intervals were 0–0.25, 0.25–0.5, 0.5–0.75, 0.75–1.0 and 1.0–1.37 m.
All J. virginiana overstory trees, saplings, and seedlings were sampled on the same 11.3 m (0.04 ha) fixed radius plot used for mature hardwood trees. All J. virginiana seedlings were counted into the same height class intervals as hardwood seedlings. Saplings over 1.37 m and under 3 m tall were recorded by count, and all J. virginiana larger than 3 m tall were measured for dbh to the nearest 0.5 cm and height to the nearest 0.1 m. Every fifth J. virginiana with dbh > 0.1 cm beginning with the most dominant and moving clockwise was selected for ageing. A core was taken if the tree was of sufficient size (approximately 10 cm dbh); otherwise, a stem disk was taken approximately 15 cm above ground level using a bow saw. An alternative numerical sampling strategy was used as described above if necessary to age a minimum of three J. virginiana stems per plot; however, because not all plots had J. virginiana present, some plots had less than three trees sampled.
We used two different methods to record standing dead tree (snag) density. We initially assumed snags would be rare and recorded snags by count and species on a circular plot with a radius of 25 m (0.19 ha). Because snag density was higher than expected, we began to collect snag data within the overstory tree plot (radius 11.3 m) and included dbh measurements from each snag and species when possible (n = 53 plots). In this manuscript, snag relative densities are reported from this subset of plots with more rigorous data collection.
2.3. Age
Juniperus virginiana will produce false rings. Because of this and the extremely narrow growth rings of some slower growing hardwoods, all cores were aged using a variable magnification sliding stage scope after progressive sanding to 400 grit using a mechanical sander. Additional hand sanding was undertaken if necessary using 3000 grit sandpaper. Cores usually included the pith. When the pith was not included in the core, the core was either discarded or the age to the pith was estimated based on growth trends for that core. This was only done when the core was judged to be close to the center (~3 rings). Stem disks were sanded and aged using a 12× hand lens or microscope. Linear regression analysis for the relationship between age (ground level) and height of J. virginiana between 1.37 and 3 m tall in our data set determined that it took approximately 9 years (8.87 year) for J. virginiana to reach a height of 1.37 m. Accordingly, we adjusted the J. virginiana ages for those measured at breast height by adding 9 years. When comparing to hardwoods, unadjusted ages of cored J. virginiana were used for comparison to hardwood ages because those were based on cores taken at breast height.
2.4. Analyses
To simplify the analyses of species relative density, we grouped similar species together. Quercus stellata, Q. marilandica and J. virginiana were analyzed as stand-alone species. The two Carya species, C. texana Buckley and C. illinoinensis (Wangenh.) K. Koch, were grouped together due to the small sample size of C. illinoinensis and similar fire tolerance. Celtis leavigata Willdenow and C. occidentalis L., which frequently hybridize and share high fire sensitivity, were grouped together. We encountered one Ulmus rubra Muhl. and included it with U. americana. We encountered both Q. velutina Lam. and Q. shumardii Buckland and grouped them together in a Red Oak group. Species that typically never reach the 10 cm dbh threshold were classified as Shrubs and were analyzed separately for relative frequency. All remaining tree species were considered Other, and had a relative density of no more than 2% in the overstory and sapling categories or 5% in the seedling category. Five-year precipitation averages were calculated using data available from the National Oceanic and Atmospheric Administration, National Centers for Environmental Information for the north central region of Oklahoma [26].
Slopes and least squares means (LS means) for the relationship between age and dbh were tested to determine if there were differences between J. virginiana, Q. stellata, and a combined Mesic species group comprising Carya spp., Celtis spp. and Ulmus spp. using Analysis of Covariance (ANCOVA) using mixed model analyses (PROC MIXED) of SAS version 9.3 (SAS Institute, Cary, NC, USA). The age vs. height and age vs. dbh relationships using ages corrected to ground level were determined using regression analysis (PROC REG) in SAS version 9.3 to calculate the growth rate of J. virginiana.
3. Results
3.1. Forest Composition
Overstory Q. stellata averaged approximately 23 cm dbh and 12.0 m in height (Table 1). It was the most frequently encountered tree species representing 171 trees ha−1 and 8.2 m2 ha−1 of basal area per plot and occurred on 92% of properties and 78% of plots. In contrast, Q. marilandica represented only 16 trees ha−1 and 0.7 m2 ha−1 of basal area (Table 2) and occurred on 52% of properties and 21% of plots (Table 3). Quercus marilandica were slightly smaller than Q. stellata, averaging 21 cm dbh and 10.7 m in height (Table 1). Quercus velutina were relatively large, i.e., 32 cm dbh and 16.3 m tall (Table 1), but only represented 2.6 m2 ha−1 basal area and 26 stems ha−1 (Table 2) occurring on 72% of properties and 35% of plots (Table 3). Carya texana (52% of properties, 22% of plots), C. occidentalis (68% of properties, 35% of plots), C. leavigata (60% of properties, 22% of plots), and U. americana (88% of properties, 38% of plots) made up the remainder of the commonly encountered hardwood tree species in the overstory (Table 3). Juniperus virginiana comprised 57 trees ha−1 and 1.3 m2 ha−1 basal area (Table 2) and occurred on 96% of properties and 53% of plots (Table 3) but were smaller than hardwood trees with an average dbh of 16 cm and average height of 8.6 m (Table 1). The average number of snags was 126.2 ha−1 with an average dbh of 23 cm.
Juniperus virginiana was common in the sapling layer (100% of properties and 84% of plots; Table 3) with an average density of 232 saplings ha−1 (Table 2). Carya texana (67 stems ha−1, 40% of properties and 16% of plots), C. occidentalis (271 stems ha−1, 56% of properties and 30% of plots) and Sapindus drummondii (67 stems ha−1, 16% of properties and 4% of plots) were also tree species that were major contributors to the sapling layer (Table 2 and Table 3). The shrub species Cercis canadensis and Cornus drummondii were frequently encountered sapling-sized stems with densities of 205 and 325 stems ha−1, respectively (Table 2). Conspicuously absent from the sapling size class were the traditionally dominant overstory tree species, Q. marilandica and Q. stellata, which together accounted for only 62 stems ha−1. Ulmus americana was also present in relatively small quantity of 52 stems ha−1. In the seedling category, U. americana accounted for 3331 ha−1 (100% of properties and 61% of plots) more than any other species. For the seedling size class, Celtis spp. were all classified as C. occidentalis due to difficulties in identification. This group averaged 2600 seedlings ha−1 (92% of properties and 61% of plots). Juniperus virginiana only had 171 stems ha−1 in the seedling layer.
3.2. Relative Dominance and Density
Most tree species had relative densities similar in magnitude to relative basal area. Quercus stellata was the dominant overstory species comprising 43% and 42% of the basal area and stem density distribution, respectively (Figure 3). Quercus marilandica, a historically abundant overstory tree in Cross Timbers forests, represented only 3% of the basal area and 4% of the relative stem density of overstory trees (Figure 3). The Red Oak group, dominated by Q. velutina and commonly associated with stream courses in the Cross Timbers, represented an outsized proportion of basal area (14%) compared to its relative density of 7% (Figure 3) due to large average dbh. Juniperus virginiana accounted for 14% of the relative density in the overstory tree size class but only 7% of the relative basal area of (Figure 3) due to smaller than average dbh. The more mesic species, i.e., Cetlis spp., Ulmus spp., and Carya spp. (we lumped C. texana with the ‘mesic’ species as it has a similar fire sensitivity and litter dynamics as the more mesic species) combined, accounted for 22% of the relative basal area and 23% of relative stem density in the overstory (Figure 3). The species distribution of snags closely mirrored the mature tree stem distribution (Figure 3). The exception was Q. marilandica, which had relative density in the snag distribution almost twice as great as its relative density in the overstory. While occurrence of Biscogniauxia spp. (formerly Hypoxylon) canker was not formally recorded, almost every Q. marilandica snag displayed signs of infection, as did some live stems. No other species were noticed to have signs of Biscogniauxia spp. canker on live trees. Quercus stellata snags frequently displayed some signs of Biscogniauxia spp., but it was not as prevalent as on Q. marilandica snags.
Juniperus virginiana stems represented 25% of the sapling relative density and the more mesic species, i.e., Cetlis spp., Ulmus spp., and Carya spp., made up an additional 48% of the saplings (Figure 3). In contrast, all Quercus species combined made up only 13% of tree saplings (Figure 3). Combined, Cetlis spp., Ulmus spp., and Carya spp. made up 60% of the seedlings (Figure 3). Juniperus virginiana’s relative density in the seedling size class was 2%, which was much lower than for the sapling and overstory size classes. Combined, the Quercus species comprised 24% of seedlings.
Cornus drumundii (53%) and C. canadensis (34%) comprised the majority of shrubs in the sapling size class. Prunus mexicana composed 8% and the ‘other’ category an additional 4%. Rhus copallina is an early successional species that was commonly encountered on forest edges, but rare within the forest interior (1%) where inventory plots were located. In total, all shrubs made up 623 stems ha−1 of 1542 total stems ha−1 in the sapling layer (40.4%). Within the seedling layer, shrub species comprised 2323 stems ha−1 out of 13,202 stems ha−1 total (17.5%) (Table 2).
3.3. Age Distribution
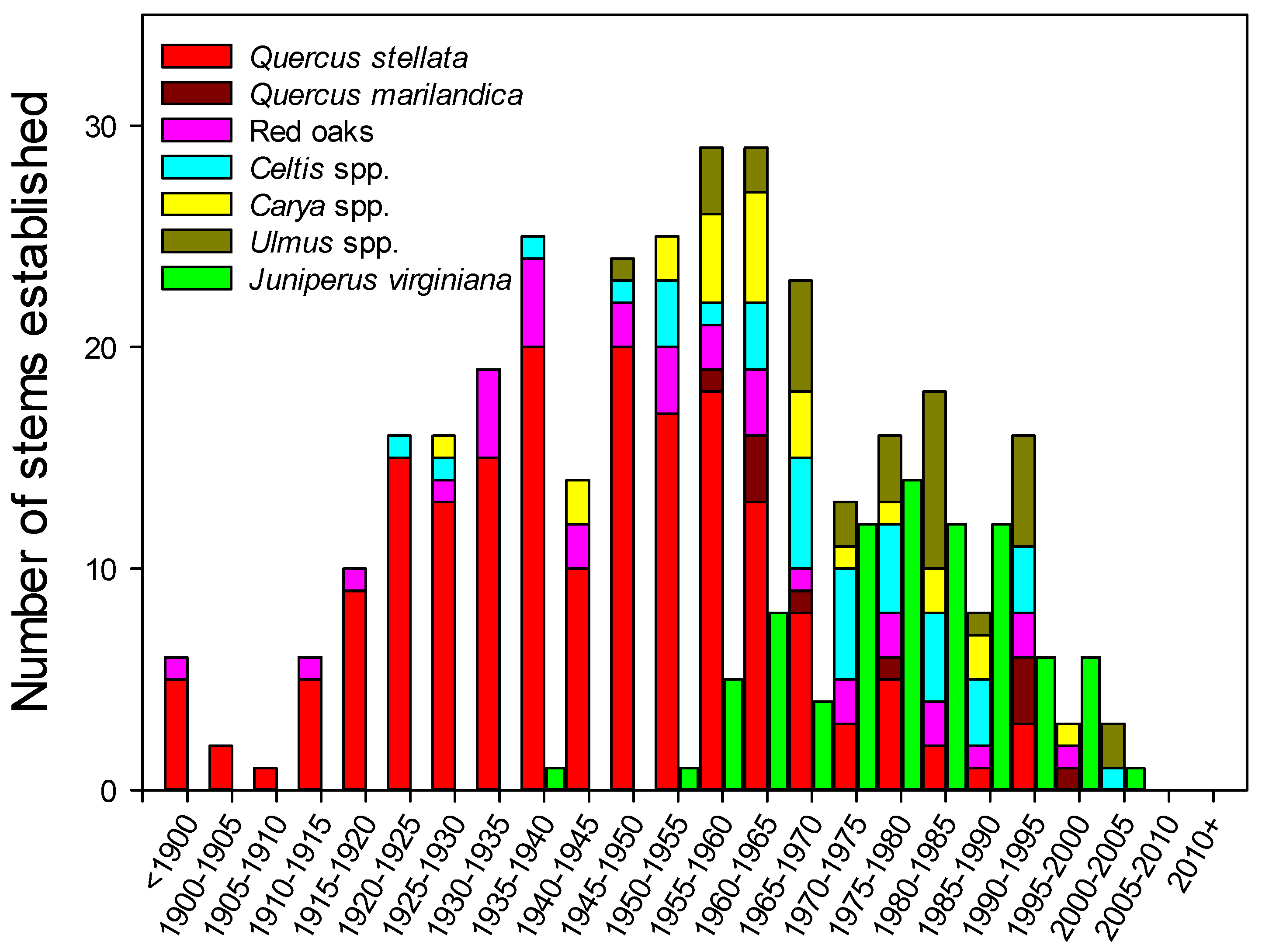
The species distribution for hardwood tree age reported in bi-decadal increments based on pith age at breast height was dominated by Q. stellata, particularly at older ages (Figure 4). The oldest tree sampled was a 233-year-old Q. stellata, which reached a height of 1.37 m between 1780 and 1785. The average Q. stellata sampled for age was 75 years old, with 83% of stems sampled between age 50 and 100 years. Quercus marilandica and Red Oaks, with a mean age of 63 and 73% stems between 40 and 90 years of age, were on average slightly younger than Q. stellata, but older than the more mesic species. The Cetlis spp., Ulmus spp., and Carya spp. stems were younger and began to reach the sapling size class after the 1950s with only six out of 92 stems sampled over 70 years of age. This group averaged 44 years of age, and 76% of stems sampled were between 20 and 60 years old. This parallels the results from the relative frequency data that seem to indicate recent and ongoing recruitment of the mesic species given their abundance in the sapling and seedling size classes. Measured at breast height, J. virginiania began to reach the sapling size class beginning in the 1950s.
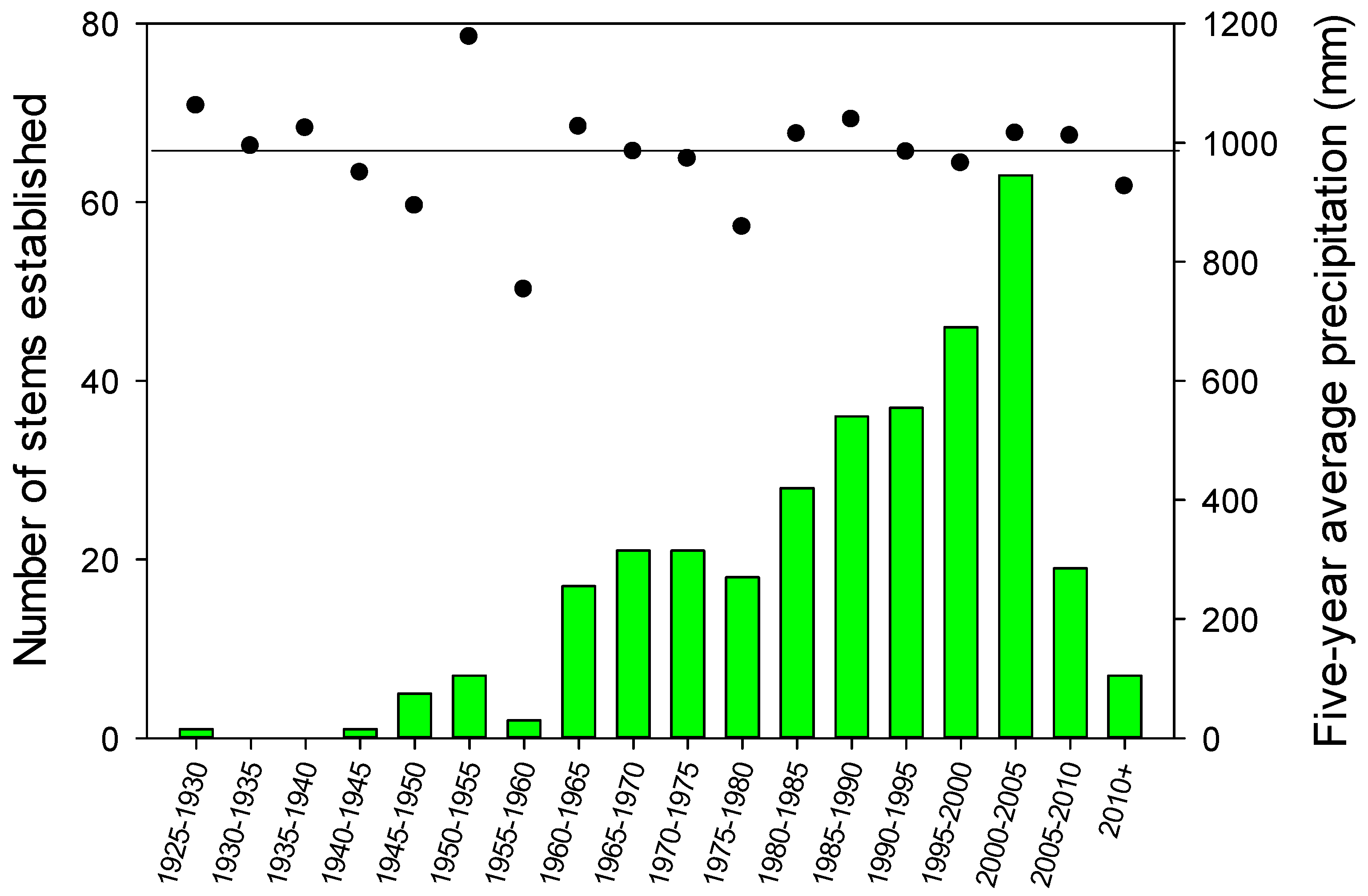
The J. virginiana age distribution of all measured trees based on pith age at ground level is presented in bi-decadal increments (Figure 5). The oldest stem dated to the last years of the 1920s. The majority of J. virginiana recruitment began in the 1950s, and the rate of recruitment increased through the early 2000s. The rate of increase between 1940 and 2005 was mostly linear with some variation associated with differences in 5-year precipitation averages. For instance, less recruitment during the 1955–1960 and 1975–1980 periods corresponded to periods of below average precipitation. Recruitment during the 2005–2010 and 2010+ intervals was lower because we did not age trees shorter than 1.4 m and many trees that recently established were thus not sampled as it takes approximately 9 years for J. virginiana to reach a height of 1.4 m.
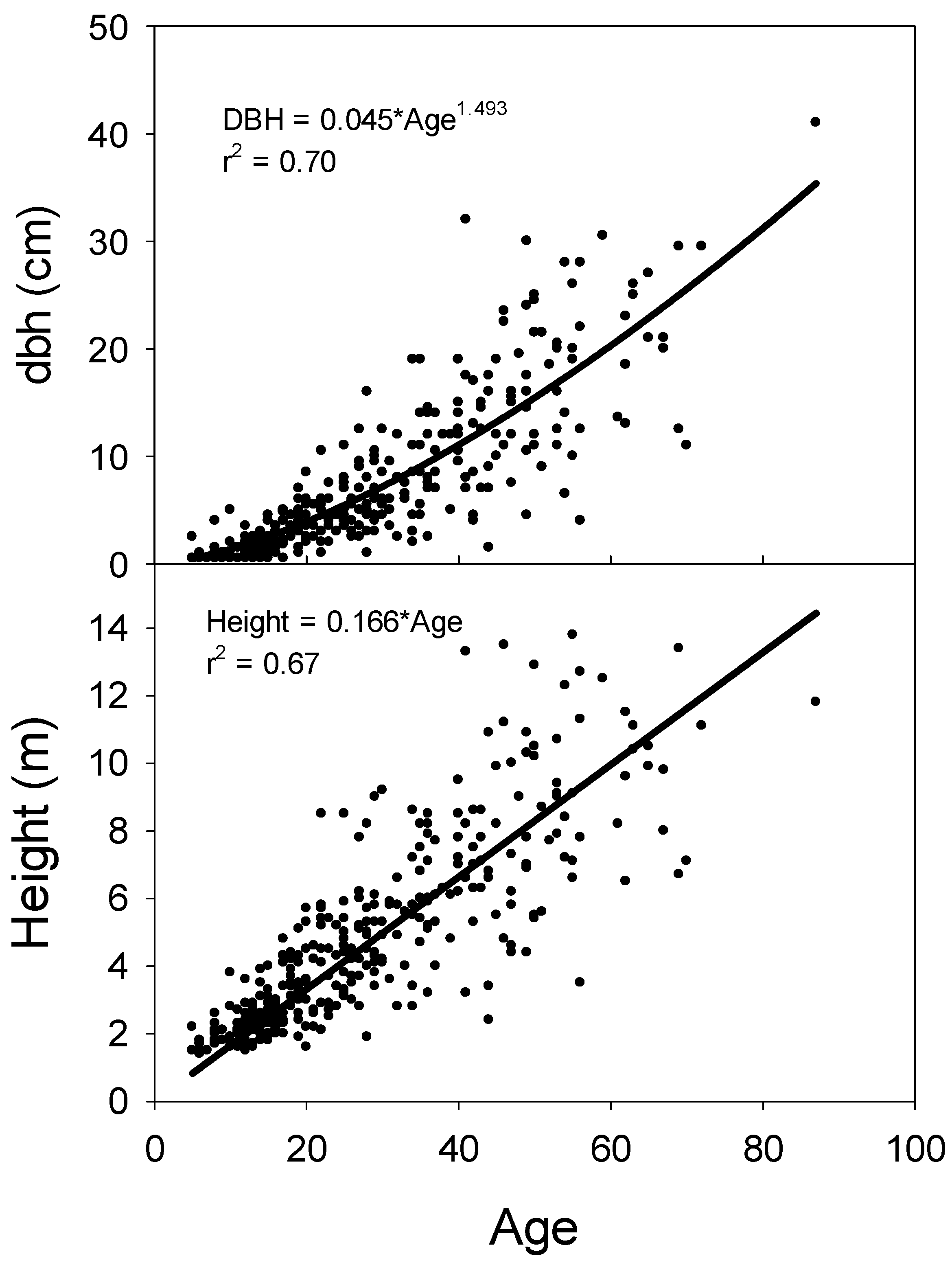
The relationship between age at ground-level and dbh for J. virginiana was best characterized by a power function (Figure 6). This equation form was superior to a linear relationship because it provided a better fit for stems with small dbh. A stem at 20 years of age added approximately 0.30 cm dbh per year, while a stem age 50 added approximately 0.45 cm dbh annually. The relationship between age and height was adequately modeled with a linear regression forced through the origin. Juniperus virginiana grew approximately 16 cm annually in height.
3.4. Age Size Relationships between Groups
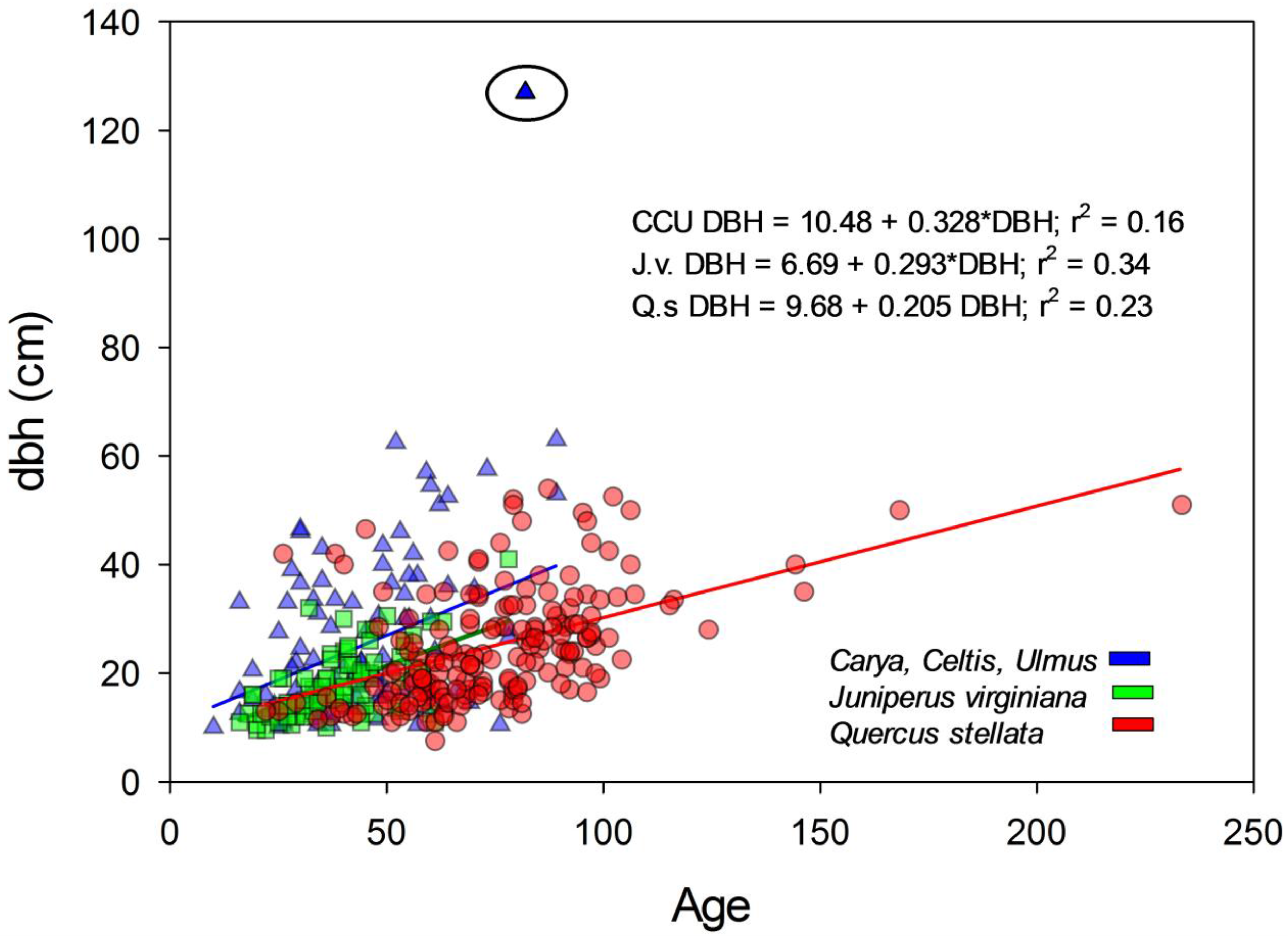
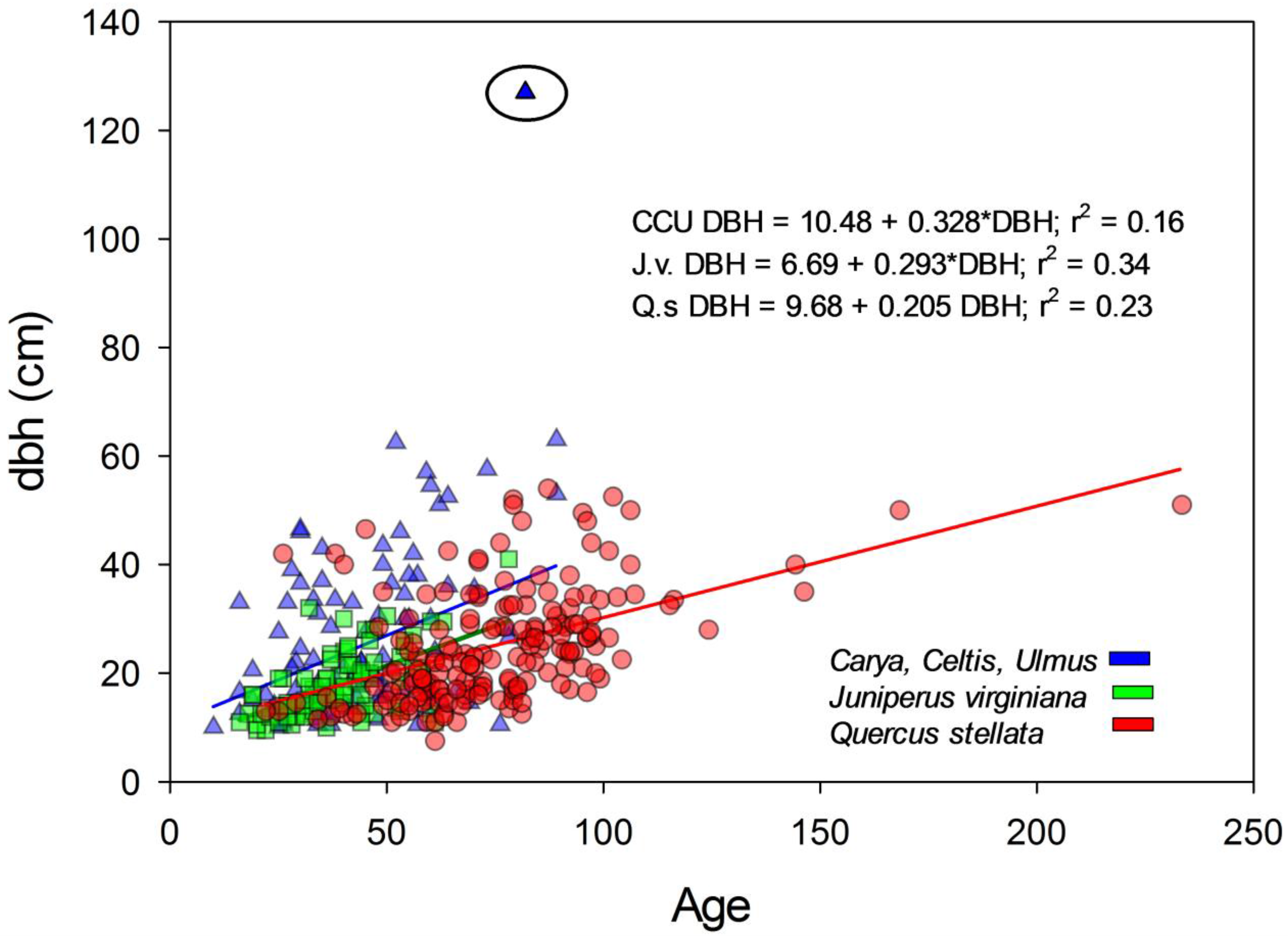
The relationship between dbh and age measured at breast height for J. virginiana, Q. stellata, and a combined grouping of Cetlis spp., Ulmus spp., and Carya spp. is presented in Figure 7. There was a C. occidentalis tree (confirmed) with a dbh of 127 cm that was 60 cm greater than the next largest tree measured for age. If the ‘outlier’ tree was removed from the analysis, the slopes were not significantly different from one another (p = 0.12). However, analysis of LS Means indicated the dbh of the mesic species grouping (28.28 ± 1.16 s.e.) was greater than Q. stellata (21.12 ± 0.82 s.e.) and J. virginiana (22.47 ± 1.16 s.e.) (p < 0.0001), indicating greater dbh for the mesic species at a given age (Figure 7). When including this ‘outlier’ tree, the slopes of the relationship between dbh and age were significantly greater for the mesic species as were the LS Means. Pairwise comparisons indicated that J. virginiana and Q. stellata slopes between age and dbh were not different (p = 0.66), but the slope of the Cetlis spp., Ulmus spp., and Carya spp. grouping was significantly greater than Q. stellata (p = 0.0001) and J. virginiana (p = 0.001). Analysis of LS Means indicated that Q. stellata (21.26 ± 0.89 s.e.) and J. virginiana (21.13 ± 1.23 s.e.) were not significantly different (p = 0.21), but that both were lower than the mesic species grouping (30.02 ± 1.13 s.e.) (p < 0.0001).
4. Discussion
The areas of privately managed Cross Timbers forests in north-central Oklahoma we measured are undergoing encroachment by J. virginiana, densification (increase in tree density and basal area), and mesophication (increase in Celtis spp., Ulmus spp., and other mesic, fire-intolerant species). The age structure as well as relative abundance of species in the overstory, sapling, and seedlings size classes indicate that this is a fairly recent and ongoing transformation. These changes have important implications for the future forest condition and successional processes related to drought and fire as well as management options for applying prescribed fire and restoration activities.
Our overstory age data agree with previous studies from Cross Timbers forests [9,21] reporting that J. virginiana recruitment accelerated after the 1950s. This coincided with fire suppression in the post WWII era and droughts during the 1950s, which opened canopy gaps due to tree mortality. Likewise, the relative density distributions among the overstory, sapling, and seedling size classes from our study support a mesophication and densification narrative, as the fire-intolerant tree species are most abundant in the sapling and seedling size classes. Three stands in north-central Oklahoma first measured in the middle of 1950s [11] and later remeasured in the late 2000s [9] exhibited an increase in basal area from 10.4 to 22.4 m2 ha−1, an increase in overstory density from 531 to 754 trees ha−1, and an increase in overstory J. virginiana basal area from 0 to 0.7 m2 ha−1 and density from 0 to 82 trees ha−1 over a period of 50 years. The stand conditions we measured were similar to those measured in the 2000s by DeSantis et al. [9] with the exception that we had approximately twice the basal area of J. virginiana, which could be related to an additional decade for J. virginana growth between the two studies.
While Q. stellata dominated the overstory, the most abundant species in the sapling size class were fire-intolerant trees and shrubs, e.g., C. drummondii, C. occidentallis, J. virginiana, C. canadensis, S. drummondii, and U. americana which indicates a potential transition away from a Quercus-dominated overstory if fire continues to be excluded in future. Similarly, the most abundant seedling species were C. occidentalis and U. americana. Although scarce in the sapling size class, Quercus spp. likely will become relatively more abundant if fire is reintroduced. Quercus stellata and Q. marilandica seedlings can survive in the understory for decades, allocating much of their acquired carbon to growing root systems which allows them to resprout multiple times following topkill from fire [27].
The faster diameter development of the fire-intolerant, mesic grouping of the Celtis spp., Ulmus spp., Carya spp. further indicates that once established and free from disturbance, the rate of meshophication may accelerate. In contrast, the dbh of more xeric J. virginiana grew approximately at the same rate as did Q. stellata. In regards to wildfire fuels, the growth of J. virginiana is particularly important. On these same sites, Hoff et al. [20] measured that J. virginiana composed 18.4% of canopy cover and added 6.3 Mg ha−1 to available wildfire fuel. Given the large number of J. virginiana in the sapling size class, these values are in the process of increasing further. Height and dbh growth rates of J. virginiana from our study were similar to another report of J. virginiana growing in a forest [28], but 20% less in height growth rate and 56% less in diameter growth rate than those reported for J. virginiana in prairie environments [18]. The relatively slow growth of J. virginiana growing in the forest interior indicates that it would take approximately 12 years to reach 2 m tall at which time they are more difficult to kill with prescribed fire. This is helpful for planning prescribed fire and indicates it may be possible to burn less frequently than the historical fire return interval (3–6 years) to accomplish management objectives related to J. virginiana reduction.
There were fewer J. virginiana seedlings than might have been expected from their high relative density in the sapling size class. The dips in J. virginiana recruitment associated with below average 5-year precipitation suggests that drought may reduce seedling/sapling establishment and survival. The recent period ending in 2015 was the driest since 1975–1980, which may help explain the low numbers of J. virginiana in the current seedling size class. Juniperus virginiana transpires water all year in the Cross Timbers [13]. While extremely drought-tolerant, J. virginiana seedlings may be more susceptible to drought than larger J. virginiana trees because seedlings have smaller, shallower root systems. Cones of J. virginina typically disperse February to March and seeds germinate the following spring such that an increase in seedlings germination from seed stored in the seedbed should not be expected post drought. Alternative explanations for the relatively low frequency of J. virginiana seedlings could be that survival of J. virginiana seedlings might be greater such that fewer seedlings are needed to achieve successful recruitment into the sapling size class or, the rate of new J. virginiana recruitment may be beginning to wane if forest density has increased the point of hindering seedling establishment.
Cross Timbers forests are considered to be dominated by Q. stellata and Q. marilandica, yet we encountered relatively few Q. marilandica in the overstory, sapling, or seedling classes. In the 1950s Rice and Penfound [11] measured that Q. marilandica had 55% and 73% as much basal area stem density as did Q. stellata. When remeasured in the 2000s by DeSantis et al. [12], Q. marilandica had only 26% and 41% as much basal area and stem density, respectively, as did Q. stellata. The decline of the relative dominance and density of Q. marilandica was due to both a decrease in Q. marilandica as well as an increase in Q. stellata [12]. While no historical data are available for the stands measured in our study, greater relative snag density for Q. marilandica probably indicates greater mortality of mature trees for this species, which reduces dominance. Drought is a predisposing factor for many pathogens including Biscogniuxia spp. infections on Quercus spp. [29]. In Cross Timbers forests, Q. marilandica seemed predisposed to infection. The years of 2011 and 2012 were among the driest years in north-central Oklahoma since the late 1970s [30]. Our field observations agree with DeSantis et al. [12] and Masters and Waymire [31], who reported high occurrence of canker on Q. marilandica in response to drought and competition. Without significant disturbance events, particularly fire to stimulate regeneration and reduce competition, Q. marilandica are disappearing and will likely continue to decline.
The Cross Timbers region frequently undergoes moderate to severe droughts, which may be exacerbated by increased competition for water within current, denser forest conditions. The increasingly more common mesic species may experience greater drought mortality during extreme events as they lack the drought tolerance of the formerly dominant oak species. This potential for increased drought mortality may further increase encroachment of J. virginiana, as it has a high physiological drought tolerance [32,33]. Therefore, J. virginiana will likely survive drought better than the mesic species [34] and fill any canopy gaps resulting from forest dieback. Future fire regimes also will be affected by ongoing changes within the Cross Timbers forests, particularly by J. virginiana encroachment. Increased fuel loading for wildfires results from this encroachment [20] and has the potential to cause stand replacing wildfires or widespread damage to overstory oaks [21]. Stand replacing fires do not have an analog in the known fire history of the Cross Timbers forests. Given the high flammability of J. virginiana, the mesophication process of Cross Timbers forests produces a different potential outcome than further east. In the Eastern Deciduous Forest, mesophication causes a loss of fire as a potential disturbance altogether. However, the encroachment of J. virginiana into Cross Timbers forests may reduce land manager’s ability to use prescribed fire, but increase the likelihood of stand replacing, crown fires.
5. Conclusions
Our data provide important insight into mesophication related to fire exclusion of Cross Timbers forests. The encroachment of J. virginiana in the tree and sapling size classes and the increase of mesic, fire-intolerant hardwood species such as Celtis spp., Carya spp. and Ulmus spp. in the sapling and seedling classes provide evidence that the composition of Cross Timbers forests is changing. The linear increase in J. virginiana recruitment into the overstory indicates that J. virginiana likely will continue to increase in dominance as the younger trees become older and larger. In addition, the presence of mesic, fire-intolerant species is expected to increase in the absence of fire due to their abundance in the sapling and seedling size classes as well as their faster growth relative to Quercus spp. or J. virginiana. These data and other reports [9,12,21] provide compelling evidence that removal of recurrent surface fire as a disturbance agent is significantly altering the trajectory of forest composition in the Cross Timbers. This has significant implications for wildfire risk and how these forests will respond to future disturbance events.
Acknowledgments
Special thanks to: OSU interns Giovanni Serreau and Caren Mendonica, Bureau of Indian Affairs Pathways students Ryan Morrison and Joe Rule for their hard work in the field, and Chris Clay for facilitating access to BIA trust lands. This work was supported by the Bureau of Indian Affairs project A16AC00016, the Oklahoma Agricultural Experiment Station, and McIntire Stennis project OKL0 2929.
Author Contributions
D.L.H., R.E.W., C.B.Z., and N.D.L. conceived and designed the study; D.L.H. collected the data; D.L.H. and R.E.W. analyzed the data; D.L.H, R.E.W., C.B.Z., and N.D.L. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kuchler, A.W. Potential Natural Vegetation of the Conterminous United States; Special Publication No. 36; American Geographical Society: New York, NY, USA, 1964. [Google Scholar]
- Therrell, M.D.; Stahle, D.W. A predictive model to locate ancient forests in the Cross Timbers of Osage County, Oklahoma. J. Biogeogr. 1998, 25, 847–854. [Google Scholar] [CrossRef]
- Hallgren, S.W.; DeSantis, R.D.; Burton, J.A. Fire and vegetation dynamics in the Cross Timbers forests of South-Central North America. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011; Dey, D.C., Stambaugh, M.C., Clark, S.L., Schweitzer, C.J., Eds.; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012. [Google Scholar]
- Clark, S.L.; Hallgren, S.W.; Stahle, D.W.; Lynch, T.B. Characteristics of the Keystone Ancient Forest Preserve, an old-growth forest in the Cross Timbers of Oklahoma. Nat. Area J. 2005, 25, 165–175. [Google Scholar]
- Stambaugh, M.C.; Guyette, R.P.; Godfrey, R.; McMurry, E.R.; Marschall, J.M. Fire, drought, and human history near the Western terminus of the Cross Timbers, Wichita Mountains, Oklahoma, USA. Fire Ecol. 2009, 5, 51–65. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Stahle, D.W. Historic fire regime of an upland oak forest in south-central North America. Fire Ecol. 2010, 6, 45–61. [Google Scholar] [CrossRef]
- Allen, M.S.; Palmer, M.W. Fire history of a prairie/forest boundary: More than 250 years of frequent fire in a North American tallgrass prairie. J. Veg. Sci. 2011, 22, 436–444. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Sparks, J.C.; Guyette, R.P.; Willson, G. Fire history of a relict oak woodland in northeast Texas. Rangel. Ecol. Manag. 2011, 64, 419–423. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Stahle, D.W. Drought and fire suppression lead to rapid forest composition change in a forest-prairie ecotone. For. Ecol. Manag. 2011, 261, 1183–1840. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the Eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Rice, E.L.; Penfound, W.T. The upland forests of Oklahoma. Ecology 1959, 40, 593–608. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Lynch, T.B.; Burton, J.A.; Palmer, M.W. Long-term directional changes in upland Quercus forests throughout Oklahoma, USA. J. Veg. Sci. 2010, 21, 606–615. [Google Scholar] [CrossRef]
- Caterina, G.L.; Will, R.E.; Turton, D.J.; Wilson, D.S.; Zou, C.B. Water use of Juniperus virginiana trees encroached into mesic prairies in Oklahoma, USA. Ecohydrology 2014, 7, 1124–1134. [Google Scholar]
- Van Els, P.; Will, R.E.; Palmer, M.W.; Hickman, K.R. Changes in forest understory associated with Juniperus encroachment in Oklahoma, USA. Appl. Veg. Sci. 2010, 13, 356–368. [Google Scholar]
- Nunes-Biral, V.C. Impacts of Tree Encroachment into a Grassland on Soil Carbon and Inputs in Northcentral Oklahoma. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2017. [Google Scholar]
- Williams, R.J.; Hallgren, S.W.; Wilson, G.W.T.; Palmer, M.W. Juniperus virginiana encroachment into upland oak forests alters arbuscular mycorrhizal abundance and litter chemistry. Appl. Soil Ecol. 2013, 65, 23–30. [Google Scholar] [CrossRef]
- Ansley, R.J.; Wiedemann, H.T. Reversing the woodland steady state: Vegetation responses during restoration of Juniperus-dominated grasslands with chaining and fire. In Western North American Juniperus Communities: A Dynamic Vegetation Type; Van Auken, O.W., Ed.; Springer: New York, NY, USA, 2008; pp. 272–290. [Google Scholar]
- Owensby, C.E.; Blan, K.R.; Eaton, B.J.; Russ, O.G. Evaluation of eastern redcedar infestations in the Northern Kansas Flint Hills. J. Range Manag. 1973, 26, 256–260. [Google Scholar] [CrossRef]
- Twidwell, D.; Fuhlendorf, S.D.; Taylor, C.A., Jr.; Rogers, W.E. Refining thresholds in coupled fire-vegetation models to improve management of encroaching woody plants in grasslands. J. Appl. Ecol. 2013, 50, 603–613. [Google Scholar] [CrossRef]
- Hoff, D.; Will, R.E.; Zou, C.B.; Weir, J.; Gregory, M.; Lillie, N. Estimating increased fuel loading within the Cross Timbers forest matrix of Oklahoma, USA due to an encroaching conifer, Juniperus virginiana, using leaf-off imagery. For. Ecol. Manag. 2018, 409, 215–224. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Marschall, J.M.; Guyette, R.P. Linking fire history to successional changes of xeric oak woodlands. For. Ecol. Manag. 2014, 320, 83–95. [Google Scholar] [CrossRef]
- Bragg, D.C.; Stahle, D.W.; Cerny, K.C. Structural attributes of two old-growth Cross Timbers stands in western Arkansas. Am. Midl. Nat. 2012, 167, 40–55. [Google Scholar] [CrossRef]
- Scott, G.F.; Boyett, J.B.; Dennis, J.; White, J.D. Supplement to the Soil Survey of Pawnee County, Oklahoma; USDA Natural Resource Conservation Service: Washington, DC, USA, 2006. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the world: A new map of life on earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Lutes, D.C.; Keane, R.E.; Caratti, J.F.; Key, C.H.; Benson, N.C.; Sutherland, S.; Gangi, L.J. FIREMON: Fire Effects Monitoring and Inventory System; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006. [Google Scholar]
- Southern Climate Impacts Planning Program. Climate Trends for Oklahoma North-Central Region. Available online: http://charts.srcc.lsu.edu/trends/ (accessed on 2 October 2017).
- Clark, S.L.; Hallgren, S.W. Dynamics of oak (Quercus marilandica and Q. stellata) reproduction in an old-growth Cross Timbers forest. Southeast. Nat. 2003, 2, 559–574. [Google Scholar]
- Lykin, R.W. Estimation of Aboveground Eastern Redcedar Biomass. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 1995. [Google Scholar]
- Despres-Loustau, M.L.; Marcais, B.; Nageleisen, L.M.; Piou, D.; Vannini, A. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef]
- Oklahoma Climatological Survey. The Climate of Pawnee County. Available online: http://climate.mesonet.org/county_climate/Products/County_Climatologies/county_climate_pawnee.pdf (accessed on 26 May 2017).
- Masters, R.E.; Waymire, J.R. Oak savanna restoration: Oak response to fire and thinning through 28 years. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011; Dey, D.C., Stambaugh, M.C., Clark, S.L., Schweitzer, C.J., Eds.; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012. [Google Scholar]
- Sperry, J.S.; Tyree, M.T. Water-stress-induced xylem embolism in three species of conifers. Plant Cell Environ. 1990, 13, 427–436. [Google Scholar] [CrossRef]
- Willson, C.J.; Manos, P.S.; Jackson, R.B. Hydraulic traits are influenced by phylogenetic history in the drought-resistant, invasive genus Juniperus (Cupressaceae). Am. J. Bot. 2008, 95, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Volder, A.; Tjoelker, M.G.; Briske, D.D. Contrasting physiological responsiveness of establishing trees and a C4 grass to rainfall events, intensified summer drought, and warming in oak savanna. Glob. Chang. Biol. 2010, 16, 3349–3362. [Google Scholar] [CrossRef]
Figure 1.
Study area represented by the black rectangle. The lightly shaded area is the Terrestrial Ecoregions of the World [24] designation for the transition between central forest and grasslands. The dark area represents the Cross Timbers from the Potential Natural Vegetation Type for the Continental U.S. [1]. This ecotone is commonly referred to as the Cross Timbers in Texas, Oklahoma and Kansas.
Figure 1.
Study area represented by the black rectangle. The lightly shaded area is the Terrestrial Ecoregions of the World [24] designation for the transition between central forest and grasslands. The dark area represents the Cross Timbers from the Potential Natural Vegetation Type for the Continental U.S. [1]. This ecotone is commonly referred to as the Cross Timbers in Texas, Oklahoma and Kansas.
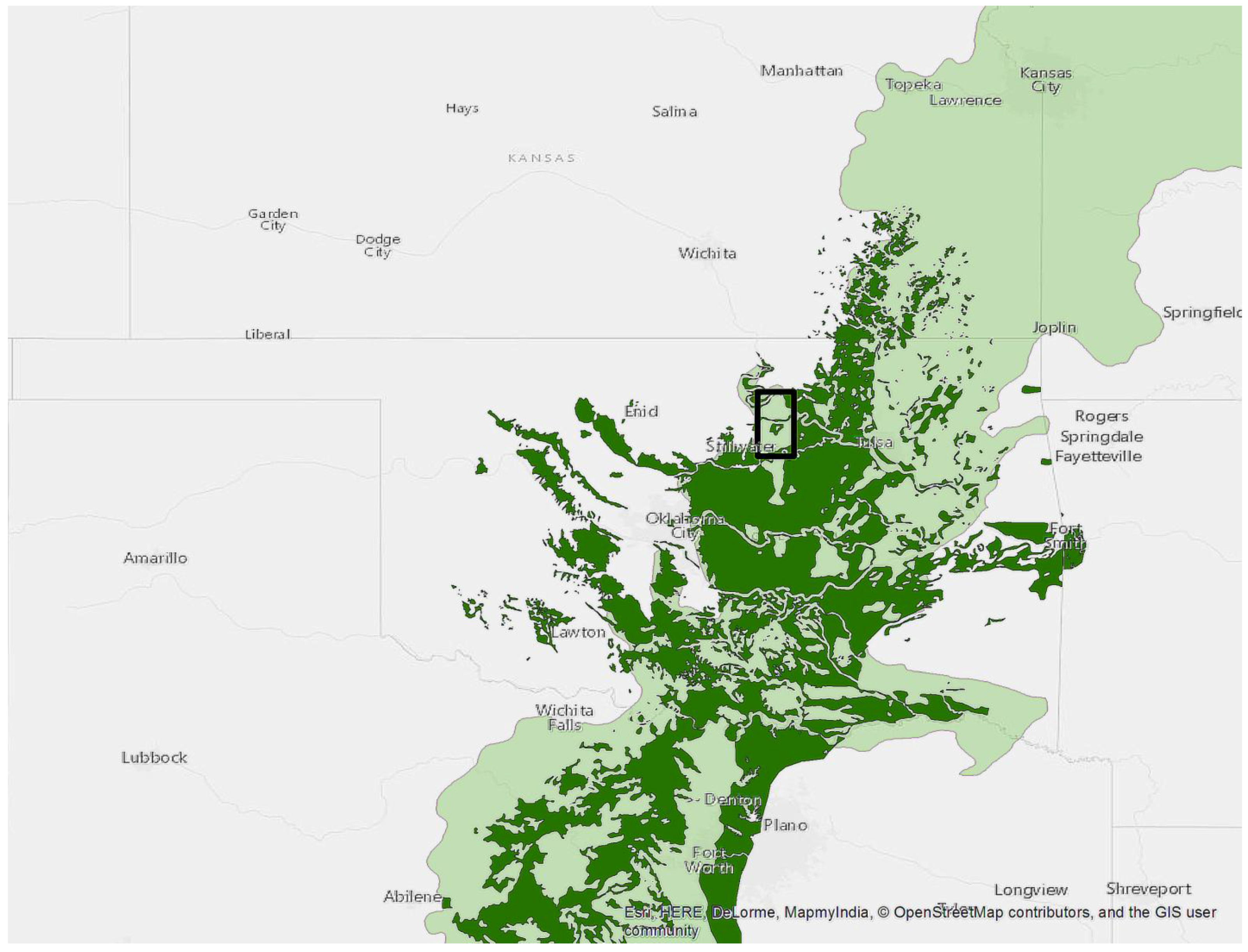
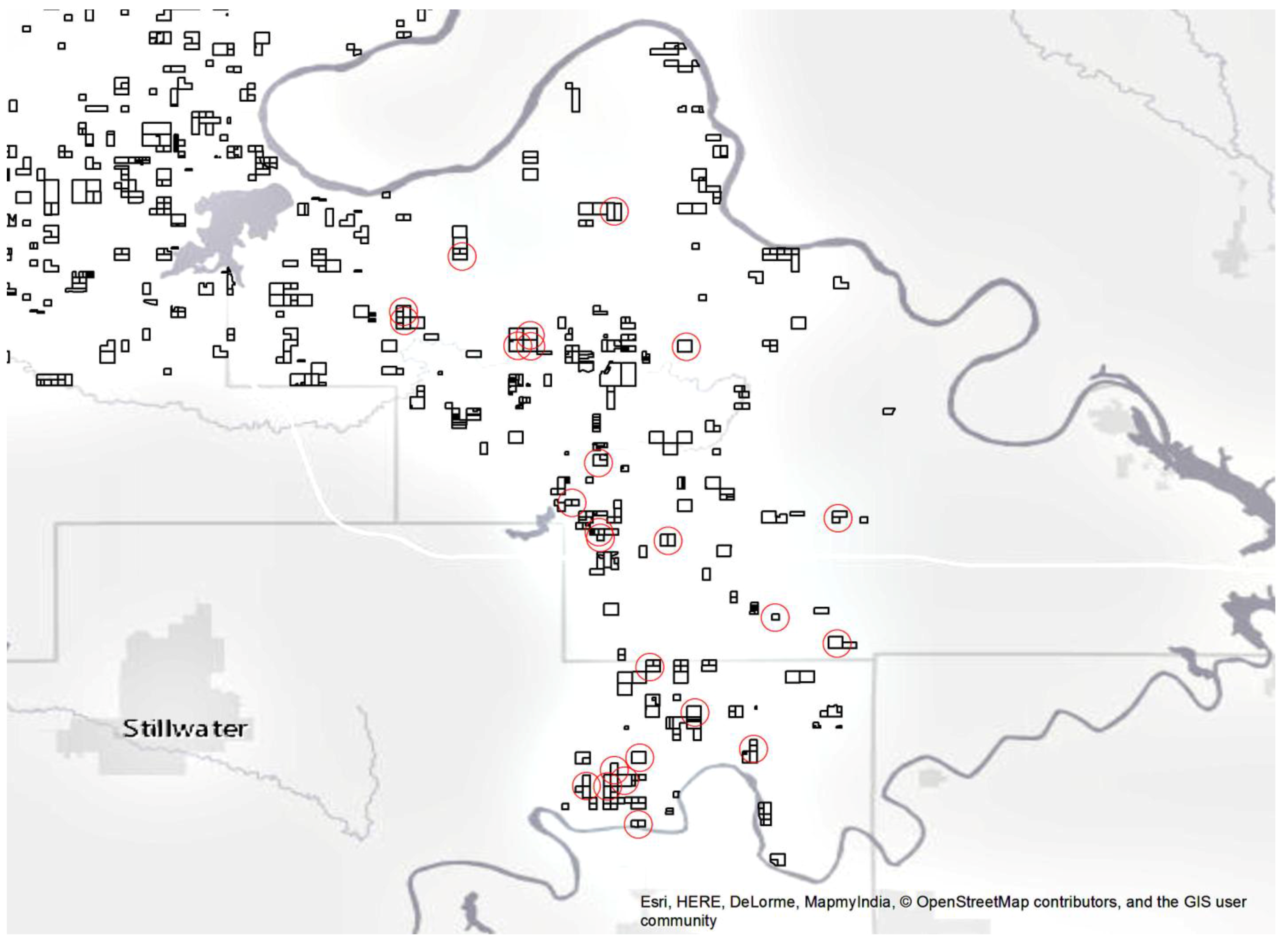
Figure 2.
BIA-managed properties in this area of the Southern Plains region are outlined in black. The 25 properties randomly selected for field measurements in this study are circled in red.
Figure 2.
BIA-managed properties in this area of the Southern Plains region are outlined in black. The 25 properties randomly selected for field measurements in this study are circled in red.
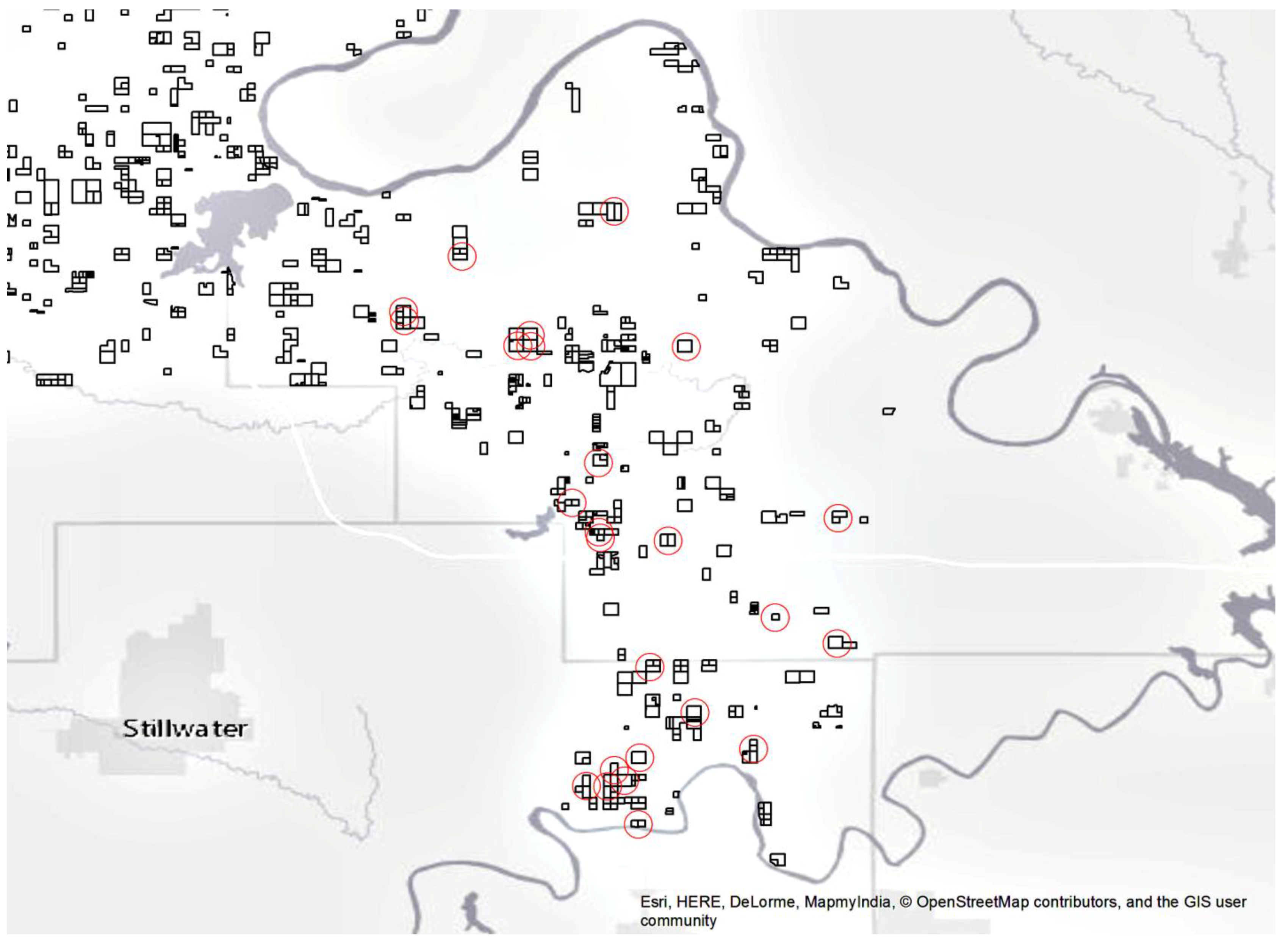
Figure 3.
Tree relative dominance based on basal area (Tree RBA) as well as overstory tree, sapling, seedling, and snag relative density (RD) measured in Cross Timbers forests of north-central Oklahoma. Categories are either individual species or groupings of related species.
Figure 3.
Tree relative dominance based on basal area (Tree RBA) as well as overstory tree, sapling, seedling, and snag relative density (RD) measured in Cross Timbers forests of north-central Oklahoma. Categories are either individual species or groupings of related species.
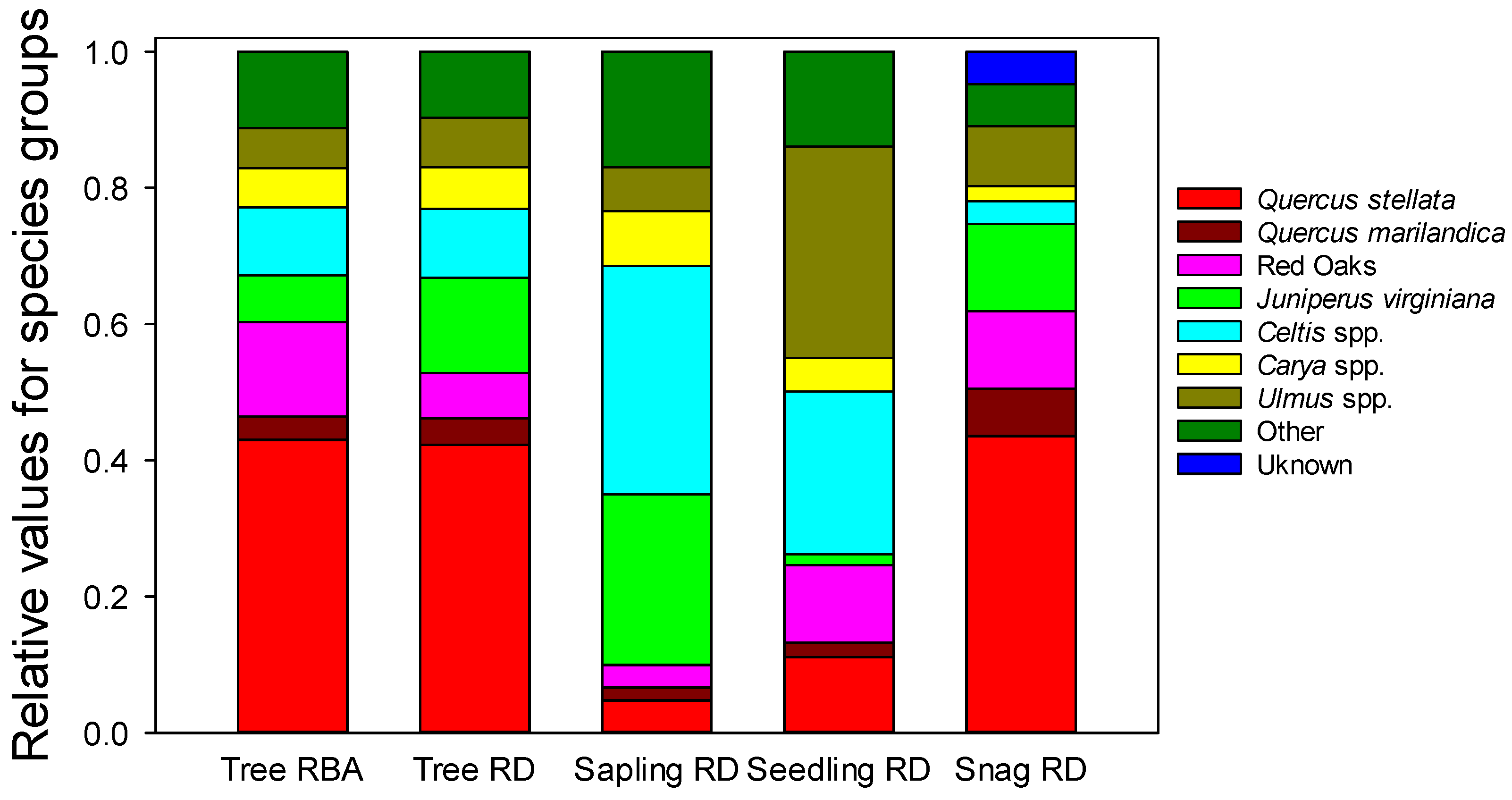
Figure 4.
Major tree species associations measured in Cross Timbers forests of north-central Oklahoma presented in bi-decadal increments. All data were collected using increment cores at breast height such that each data point represents the 5-year period when the stem was recruited into the sapling size class. Juniperus virginiana were sampled separately from the other species and are offset for clarity of display.
Figure 4.
Major tree species associations measured in Cross Timbers forests of north-central Oklahoma presented in bi-decadal increments. All data were collected using increment cores at breast height such that each data point represents the 5-year period when the stem was recruited into the sapling size class. Juniperus virginiana were sampled separately from the other species and are offset for clarity of display.
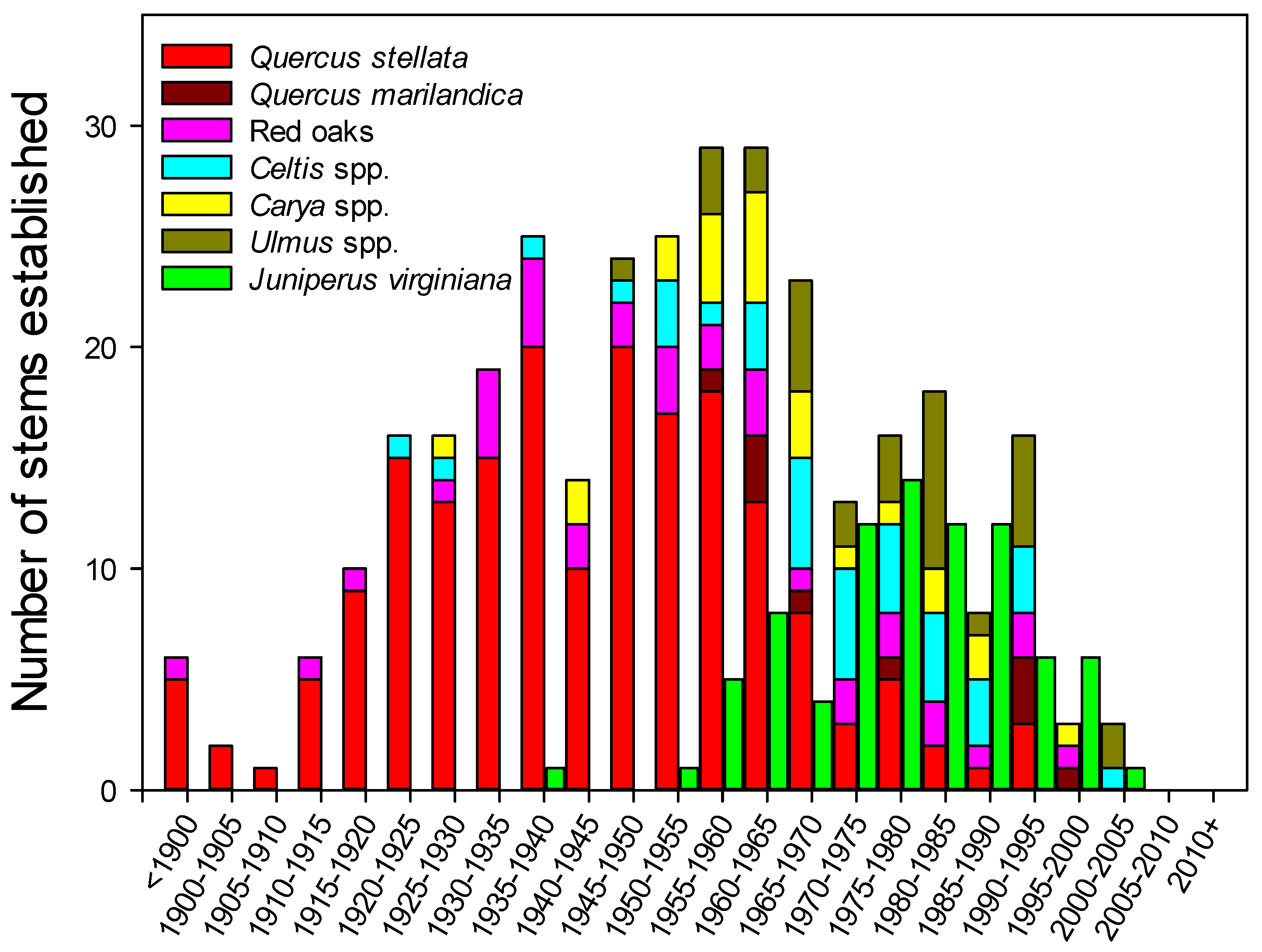
Figure 5.
All Juniperus virginiana stems by bi-decadal age class with 5-year precipitation average. Data were based on stem disks at ground level or from cores at breast height that were corrected to year of establishment. Young J. virginiana with establishment dates after 2007 are not expected to have reached breast height and were not included in the sampling.
Figure 5.
All Juniperus virginiana stems by bi-decadal age class with 5-year precipitation average. Data were based on stem disks at ground level or from cores at breast height that were corrected to year of establishment. Young J. virginiana with establishment dates after 2007 are not expected to have reached breast height and were not included in the sampling.
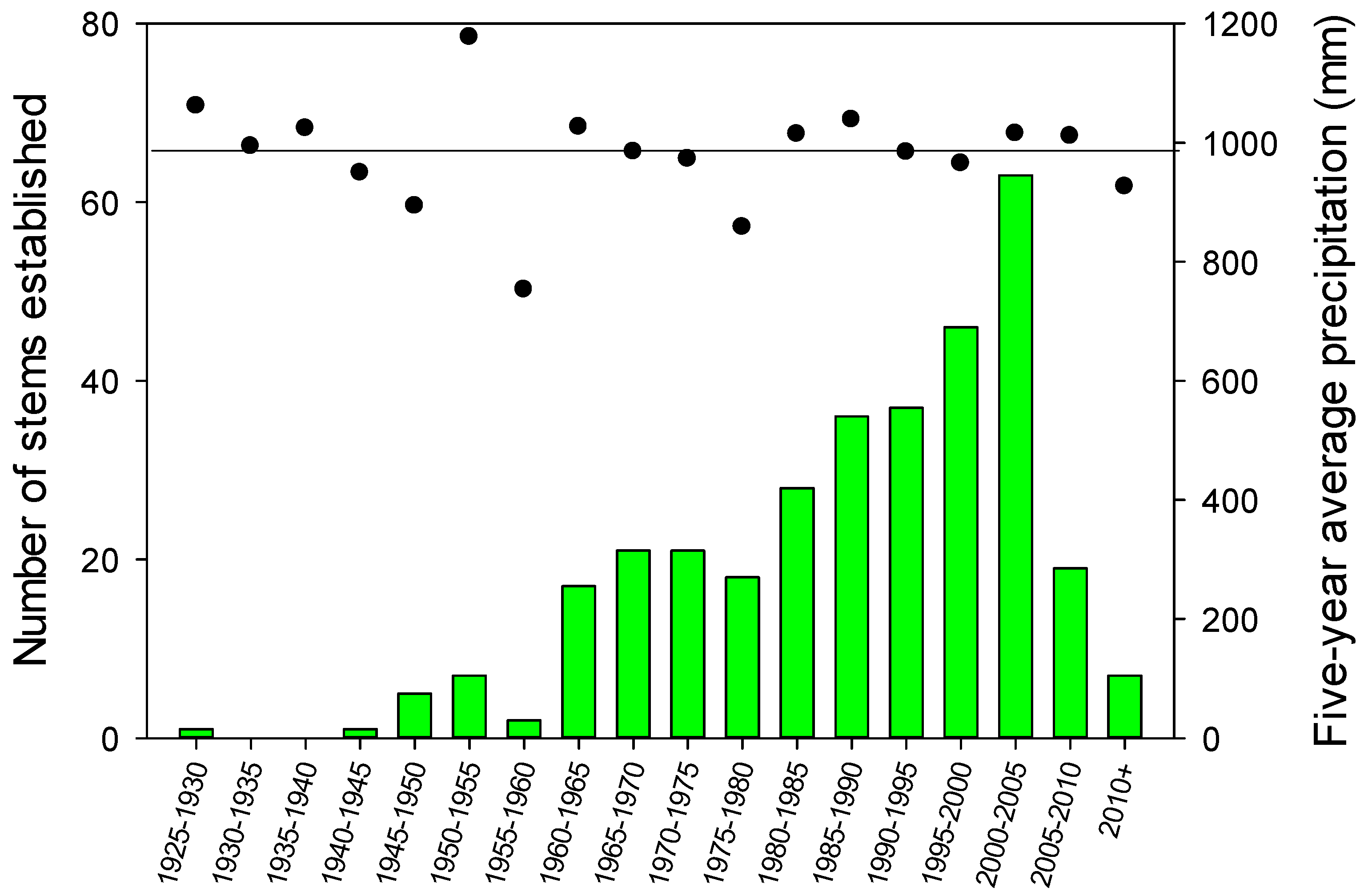
Figure 6.
Relationship between age and dbh and relationship between age and height for Juniperus virginiana stems. Data were based on stem disks at ground level or from cores at breast height that were corrected to year of establishment. Trees shorter than 1.37 m were not sampled for age.
Figure 6.
Relationship between age and dbh and relationship between age and height for Juniperus virginiana stems. Data were based on stem disks at ground level or from cores at breast height that were corrected to year of establishment. Trees shorter than 1.37 m were not sampled for age.
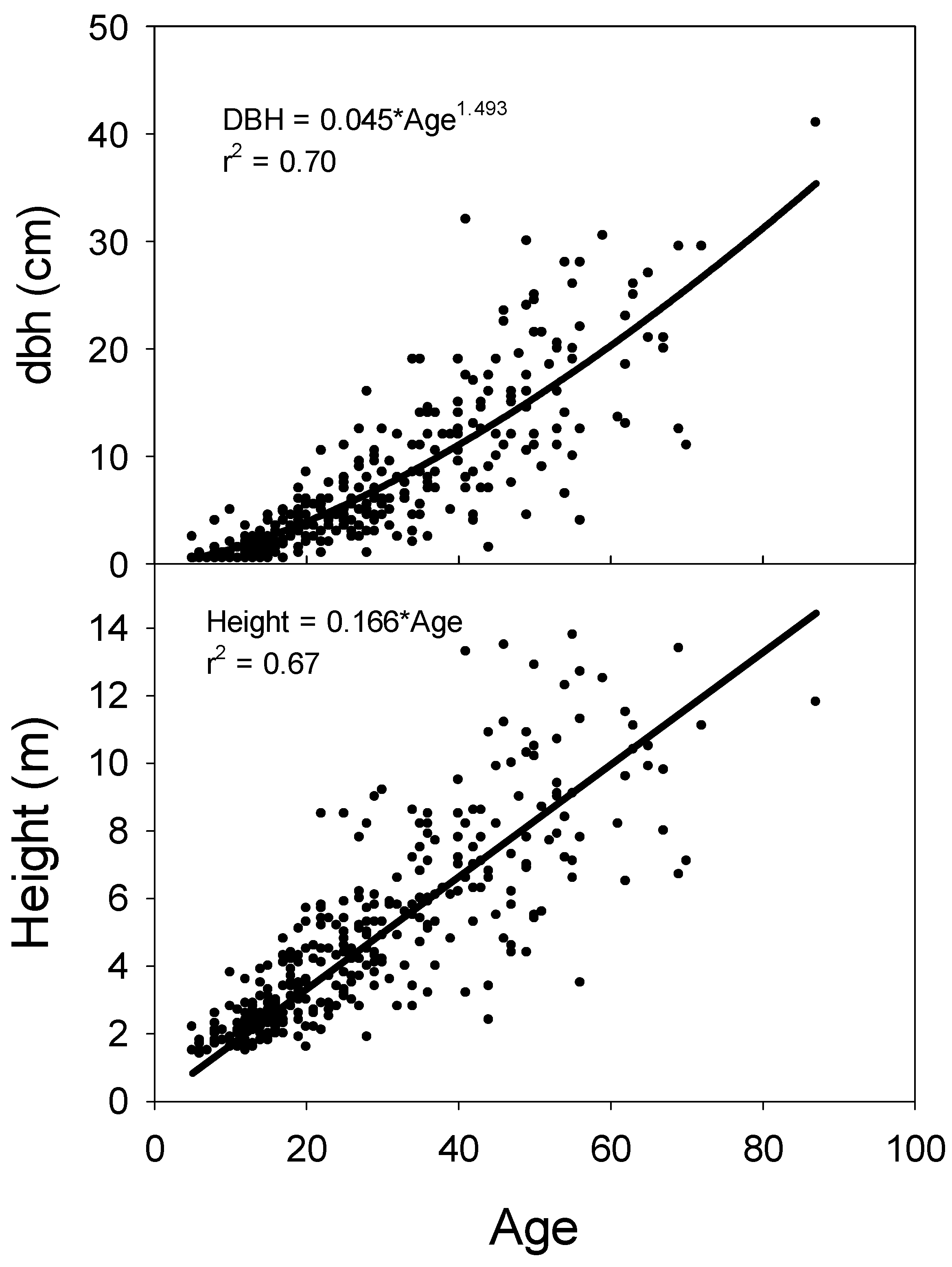
Figure 7.
Relationship between age and dbh for Juniperus virginiana (J.v.), Quercus stellata (Q.s.), and a combined Mesic group of Carya spp., Celtis spp., and Ulmus spp. (CCU). All ages were taken at dbh and not adjusted. The circled outlier for the mesic species grouping was removed for calculation of relationship between age and dbh.
Figure 7.
Relationship between age and dbh for Juniperus virginiana (J.v.), Quercus stellata (Q.s.), and a combined Mesic group of Carya spp., Celtis spp., and Ulmus spp. (CCU). All ages were taken at dbh and not adjusted. The circled outlier for the mesic species grouping was removed for calculation of relationship between age and dbh.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number encountered (N) and average dbh and height of trees (dbh > 10 cm), sapling (dbh 0.1 to 10 cm) and seedlings (shorter than 1.37 m) from 130 plots measured in Cross Timbers forests of north-central Oklahoma. Std. = standard deviation. Trees were measured in plots 0.04 ha in size (5.2 ha total). For hardwoods, saplings were measured in plots 0.004 ha in size (0.52 ha total) and seedlings in plots 0.001 ha in size (0.13 ha total). Juniperus virginiana L. overstory, saplings and seedlings were measured in the 0.04 ha plots (5.2 ha total). For seedlings, all Celtis spp. are lumped as Celtis occidentalis L.
Table 1.
Number encountered (N) and average dbh and height of trees (dbh > 10 cm), sapling (dbh 0.1 to 10 cm) and seedlings (shorter than 1.37 m) from 130 plots measured in Cross Timbers forests of north-central Oklahoma. Std. = standard deviation. Trees were measured in plots 0.04 ha in size (5.2 ha total). For hardwoods, saplings were measured in plots 0.004 ha in size (0.52 ha total) and seedlings in plots 0.001 ha in size (0.13 ha total). Juniperus virginiana L. overstory, saplings and seedlings were measured in the 0.04 ha plots (5.2 ha total). For seedlings, all Celtis spp. are lumped as Celtis occidentalis L.
| Overstory Trees | Saplings | Seedlings | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dbh (cm) | Height (m) | dbh (cm) | Height (m) | Height (m) | |||||||||
| Genus Species | N | Mean | Std. | Mean | Std. | N | Mean | Std. | Mean | Std. | N | Mean | Std. |
| Acer negundo L. | 2 | 21.0 | 14.1 | - | - | 5 | 4.6 | 2.6 | 4.9 | 1.7 | 0 | - | - |
| Carya illinoinensis (Wangenh.) K.Koch | 37 | 27.8 | 14.1 | 13.3 | 5.5 | 4 | 4.5 | 3.0 | 5.2 | 2.8 | 8 | 0.4 | 0.2 |
| Carya texana Buckley | 90 | 18.3 | 8.5 | 11.9 | 4.4 | 36 | 4.3 | 2.8 | 4.2 | 2.3 | 62 | 0.4 | 0.3 |
| Celtis laevigata Willdenow | 50 | 19.1 | 9.7 | 11.1 | 4.5 | 21 | 2.8 | 2.0 | 3.6 | 1.9 | 0 | - | - |
| Celtis occidentalis L. | 164 | 20.8 | 14.1 | 11.3 | 4.7 | 141 | 2.4 | 2.1 | 3.1 | 1.6 | 338 | 0.4 | 0.3 |
| Cercis canadensis L. | 4 | 10.9 | 1.0 | - | - | 107 | 2.4 | 2.1 | 2.9 | 1.2 | 69 | 0.5 | 0.3 |
| Cornus drummondii C.A.Mey. | 0 | - | - | - | - | 149 | 1.1 | 0.7 | 2.0 | 0.9 | 158 | 0.8 | 0.4 |
| Diospyros virginiana L. | 10 | 18.0 | 7.7 | 10.5 | 5.1 | 8 | 4.0 | 2.4 | 4.2 | 1.5 | 8 | 0.6 | 0.3 |
| Fraxinus spp. L. | 36 | 28.6 | 13.8 | 15.7 | 4.6 | 6 | 3.0 | 2.5 | 3.5 | 2.1 | 5 | 0.8 | 0.3 |
| Gleditsia triacanthos L. | 1 | 12.5 | - | - | - | 1 | 3.0 | - | 4.4 | - | 3 | 0.5 | 0.1 |
| Gymnocladus dioicus (L.) K. Koch | 6 | 21.8 | 22.2 | 12.9 | 5.5 | 0 | - | - | - | - | 0 | - | - |
| Juglans nigra L. | 23 | 23.8 | 16.0 | 10.3 | 4.9 | 3 | 6.3 | 3.1 | 4.7 | 1.7 | 0 | - | - |
| Juniperus virginiana L. | 296 | 16.4 | 6.2 | 8.6 | 2.1 | 1210 | 3.0 | 2.4 | 3.2 | 1.5 | 891 | 0.7 | 0.4 |
| Morus rubra L. | 29 | 19.5 | 9.4 | 12.0 | 3.7 | 0 | - | - | - | - | 5 | 0.5 | 0.2 |
| Platanus occidentalis L. | 11 | 36.7 | 13.9 | 18.7 | 0.1 | 0 | - | - | - | - | 0 | - | - |
| Prunus angustifolia Marsh. | 2 | 11.0 | 1.4 | - | - | 0 | - | - | - | - | 0 | - | - |
| Prunus mexicana S.Watson | 8 | 12.9 | 2.4 | 6.4 | 2.5 | 26 | 1.9 | 1.7 | 4.6 | 1.9 | 54 | 0.8 | 0.4 |
| Prunus serotina Ehrh. | 0 | - | - | - | - | 0 | - | - | - | - | 2 | 0.4 | 0.0 |
| Quercus macrocarpa Michx. | 10 | 24.2 | 15.1 | 10.7 | 2.1 | 0 | - | - | - | - | 2 | 0.4 | 0.0 |
| Quercus marilandica Muenchh. | 82 | 21.1 | 9.3 | 10.7 | 2.7 | 9 | 5.2 | 3.1 | 2.4 | 0.9 | 30 | 0.2 | 0.2 |
| Quercus muehlenbergii Engelm. | 18 | 27.0 | 13.8 | 10.3 | 2.8 | 11 | 2.5 | 2.4 | 4.4 | 1.8 | 57 | 0.3 | 0.3 |
| Quercus shumardii Buckland | 2 | 31.0 | 0.7 | 13.1 | - | 0 | - | - | - | - | 0 | - | - |
| Quercus stellata Wangenh. | 888 | 22.9 | 9.3 | 12.0 | 3.2 | 23 | 4.7 | 3.3 | 3.3 | 2.3 | 157 | 0.3 | 0.2 |
| Quercus velutina Lam. | 137 | 31.7 | 16.2 | 16.3 | 4.3 | 16 | 2.9 | 2.6 | 1.8 | 0.5 | 161 | 0.3 | 0.3 |
| Rhus copallina L. | 0 | - | - | - | - | 4 | 1.0 | 0.0 | 2.1 | 0.4 | 18 | 0.4 | 0.3 |
| Robinia pseudoacacia L. | 0 | - | - | - | - | 4 | 1.5 | 1.0 | 3.2 | 1.5 | 2 | 1.0 | 0.2 |
| Salix nigra Marshall | 3 | 37.5 | 5.8 | 12.4 | - | 0 | - | - | - | - | 0 | - | - |
| Sapindus drummondii L. | 14 | 15.9 | 4.1 | 9.3 | 1.9 | 35 | 2.3 | 1.8 | 3.6 | 1.6 | 31 | 0.3 | 0.3 |
| Sideroxylon languinosum Michx. | 41 | 17.8 | 6.0 | 9.5 | 2.5 | 9 | 2.1 | 2.3 | 2.8 | 1.1 | 82 | 0.4 | 0.3 |
| Ulmus americana L. | 150 | 18.8 | 10.7 | 11.0 | 3.5 | 27 | 3.5 | 2.9 | 3.8 | 1.4 | 433 | 0.3 | 0.2 |
| Ulmus rubra Muhl. | 3 | 34.2 | 26.5 | 20.1 | - | 4 | 3.0 | 2.3 | 2.8 | 0.2 | 6 | 0.1 | 0.0 |
| Viburnum rufidulum Raf. | 0 | - | - | - | - | 3 | 1.7 | 1.2 | 1.6 | 0.1 | 3 | 0.3 | 0.1 |
| Zanthoxylum americanum Mill. | 0 | - | - | - | - | 10 | 1.0 | 0.0 | 2.9 | 1.6 | 0 | - | - |
Table 2.
Average basal area (BA) and density for overstory trees (dbh >10 cm) and average density for sapling (dbh 0.1 to 10 cm) and seedlings (shorter than 1.37 m) measured in 130 plots in Cross Timbers forests of north-central Oklahoma. Std. = standard deviation. For seedlings, all Celtis spp. are considered Celtis occidentalis.
Table 2.
Average basal area (BA) and density for overstory trees (dbh >10 cm) and average density for sapling (dbh 0.1 to 10 cm) and seedlings (shorter than 1.37 m) measured in 130 plots in Cross Timbers forests of north-central Oklahoma. Std. = standard deviation. For seedlings, all Celtis spp. are considered Celtis occidentalis.
| Overstory Trees | Saplings | Seedlings | ||||||
|---|---|---|---|---|---|---|---|---|
| BA (m2 ha−1) | Stems (ha−1) | Stems (ha−1) | Stems (ha−1) | |||||
| Genus Species | Mean | Std. | Mean | Std. | Mean | Std. | Mean | Std. |
| Acer negundo | 0.0 | 0.2 | 0.4 | 3.1 | 9.6 | 8.1 | 0.0 | 0.0 |
| Carya illinoinensis | 0.5 | 2.2 | 7.1 | 25.0 | 7.7 | 6.9 | 61.5 | 7.5 |
| Carya texana | 0.6 | 1.9 | 17.3 | 41.6 | 67.3 | 23.4 | 476.9 | 33.9 |
| Celtis laevigata | 0.3 | 1.1 | 9.6 | 27.0 | 40.4 | 18.5 | 0.0 | 0.0 |
| Celtis occidentalis | 1.6 | 4.1 | 31.5 | 68.8 | 271.2 | 69.9 | 2600.0 | 111.6 |
| Cercis canadensis | 0.0 | 0.1 | 0.8 | 5.3 | 205.8 | 109.4 | 530.8 | 40.1 |
| Cornus drummondii | 0.0 | 0.0 | 0.0 | 0.0 | 325.0 | 140.2 | 1215.4 | 164.7 |
| Diospyros virginiana | 0.1 | 0.5 | 1.9 | 14.1 | 15.4 | 13.3 | 61.5 | 7.5 |
| Fraxinus spp. | 0.6 | 2.2 | 6.9 | 25.3 | 11.5 | 6.8 | 38.5 | 6.5 |
| Gleditsia triacanthos | 0.0 | 0.0 | 0.2 | 2.2 | 1.9 | 2.4 | 23.1 | 6.6 |
| Gymnocladus dioicus | 0.1 | 0.8 | 1.2 | 8.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| Juglans nigra | 0.3 | 1.6 | 4.4 | 15.7 | 5.8 | 4.2 | 0.0 | 0.0 |
| Juniperus virginiana | 1.3 | 3.3 | 56.5 | 117.2 | 232.7 | 429.1 | 171.4 | 484.0 |
| Morus rubra | 0.2 | 0.7 | 5.6 | 16.0 | 0.0 | 0.0 | 38.5 | 5.7 |
| Platanus occidentalis | 0.3 | 2.0 | 2.1 | 19.9 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus angustifolia | 0.0 | 0.0 | 0.4 | 4.4 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus mexicana | 0.0 | 0.1 | 1.5 | 6.0 | 50.0 | 19.2 | 415.4 | 39.4 |
| Prunus serotina | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.4 | 3.1 |
| Quercus macrocarpa | 0.1 | 0.7 | 1.9 | 9.7 | 0.0 | 0.0 | 15.4 | 4.4 |
| Quercus marilandica | 0.7 | 2.4 | 15.8 | 48.3 | 17.3 | 11.0 | 230.8 | 17.5 |
| Quercus muehlenbergii | 0.3 | 1.0 | 3.5 | 10.2 | 21.2 | 12.9 | 438.5 | 36.4 |
| Quercus shumardii | 0.0 | 0.2 | 0.4 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| Quercus stellata | 8.2 | 8.1 | 170.8 | 174.4 | 44.2 | 21.4 | 1207.7 | 71.3 |
| Quercus velutina | 2.6 | 5.3 | 26.4 | 45.3 | 30.8 | 11.9 | 1238.5 | 69.2 |
| Rhus copallina | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 9.8 | 138.5 | 17.5 |
| Robinia pseudoacacia | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 7.7 | 15.4 | 3.1 |
| Salix nigra | 0.1 | 0.7 | 0.6 | 6.6 | 0.0 | 0.0 | 0.0 | 0.0 |
| Sapindus drummondii | 0.1 | 0.5 | 2.7 | 21.6 | 67.3 | 45.0 | 238.5 | 41.6 |
| Sideroxylon lanuginosum | 0.2 | 0.7 | 7.9 | 21.8 | 17.3 | 7.0 | 630.8 | 29.1 |
| Ulmus americana | 1.1 | 2.7 | 28.9 | 64.7 | 51.9 | 20.2 | 3330.8 | 198.4 |
| Ulmus rubra | 0.1 | 0.7 | 0.6 | 3.8 | 7.7 | 5.9 | 46.2 | 9.3 |
| Viburnum rufidulum | 0.0 | 0.0 | 0.0 | 0.0 | 5.8 | 4.2 | 23.1 | 3.8 |
| Zanthoxylum americanum | 0.0 | 0.0 | 0.0 | 0.0 | 19.2 | 24.4 | 0.0 | 0.0 |
| Total | 19.0 | - | 407.0 | - | 1542.0 | - | 13,202.0 | - |
Table 3.
Number of times that species were identified on properties (Prop; 25) or plots (130, 5 plots per property with 1 property having 10) for overstory trees (dbh >10 cm), saplings (dbh 0.1 to 10 cm), and seedlings (shorter than 1.37 m), expressed as a percent. Overstory trees were measured in plots 0.04 ha in size. For hardwoods, saplings were measured in plots 0.004 ha in size and seedlings were measured in plots 0.001 ha in size. Juniperus virginiana saplings and seedlings were measured in the 0.04 ha plots. For seedlings, all Celtis spp. are lumped as Celtis occidentalis.
Table 3.
Number of times that species were identified on properties (Prop; 25) or plots (130, 5 plots per property with 1 property having 10) for overstory trees (dbh >10 cm), saplings (dbh 0.1 to 10 cm), and seedlings (shorter than 1.37 m), expressed as a percent. Overstory trees were measured in plots 0.04 ha in size. For hardwoods, saplings were measured in plots 0.004 ha in size and seedlings were measured in plots 0.001 ha in size. Juniperus virginiana saplings and seedlings were measured in the 0.04 ha plots. For seedlings, all Celtis spp. are lumped as Celtis occidentalis.
| Overstory | Sapling | Seedling | ||||
|---|---|---|---|---|---|---|
| Genus Species | Prop | Plot | Prop | Plot | Prop | Plot |
| Acer negundo | 8 | 2 | 8 | 2 | 0 | 0 |
| Carya illinoinensis | 24 | 10 | 8 | 2 | 24 | 5 |
| Carya texana | 52 | 22 | 40 | 16 | 60 | 21 |
| Celtis laevigata | 60 | 22 | 28 | 8 | 0 | 0 |
| Celtis occidentalis | 68 | 35 | 56 | 30 | 92 | 61 |
| Cercis canadensis | 8 | 2 | 56 | 27 | 48 | 17 |
| Cornus drummondii | 0 | 0 | 48 | 18 | 60 | 18 |
| Diospyros virginiana | 12 | 3 | 8 | 2 | 24 | 5 |
| Fraxinus spp. | 40 | 12 | 8 | 4 | 4 | 2 |
| Gleditsia triacanthos | 4 | 1 | 4 | 1 | 4 | 1 |
| Gymnocladus dioicus | 12 | 2 | 0 | 0 | 0 | 0 |
| Juglans nigra | 28 | 9 | 8 | 2 | 0 | 0 |
| Juniperus virginiana | 96 | 53 | 100 | 84 | 92 | 46 |
| Morus rubra | 52 | 14 | 0 | 0 | 16 | 3 |
| Platanus occidentalis | 12 | 2 | 0 | 0 | 0 | 0 |
| Prunus angustifolia | 4 | 1 | 0 | 0 | 0 | 0 |
| Prunus mexicana | 28 | 6 | 24 | 10 | 48 | 16 |
| Prunus serotina | 0 | 0 | 0 | 0 | 4 | 2 |
| Quercus macrocarpa | 16 | 5 | 0 | 0 | 4 | 1 |
| Quercus marilandica | 52 | 21 | 12 | 4 | 44 | 14 |
| Quercus muehlenbergii | 44 | 12 | 12 | 4 | 44 | 17 |
| Quercus shumardii | 8 | 2 | 0 | 0 | 0 | 0 |
| Quercus stellata | 92 | 78 | 20 | 6 | 88 | 34 |
| Quercus velutina | 72 | 35 | 36 | 10 | 68 | 33 |
| Rhus copallina | 0 | 0 | 4 | 1 | 24 | 6 |
| Robinia pseudoacacia | 0 | 0 | 4 | 2 | 8 | 2 |
| Salix nigra | 4 | 1 | 0 | 0 | 0 | 0 |
| Sapindus drummondii | 8 | 2 | 16 | 4 | 16 | 5 |
| Sideroxylon lanuginosum | 60 | 18 | 28 | 7 | 88 | 34 |
| Ulmus americana | 88 | 38 | 28 | 12 | 100 | 61 |
| Ulmus rubra | 8 | 2 | 8 | 2 | 8 | 2 |
| Viburnum rufidulum | 0 | 0 | 8 | 2 | 12 | 2 |
| Zanthoxylum americanum | 0 | 0 | 4 | 1 | 0 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hoff, D.L.; Will, R.E.; Zou, C.B.; Lillie, N.D. Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA. Forests 2018, 9, 75. https://doi.org/10.3390/f9020075
AMA Style
Hoff DL, Will RE, Zou CB, Lillie ND. Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA. Forests. 2018; 9(2):75. https://doi.org/10.3390/f9020075
Chicago/Turabian StyleHoff, Daniel L., Rodney E. Will, Chris B. Zou, and Nathan D. Lillie. 2018. "Encroachment Dynamics of Juniperus virginiana L. and Mesic Hardwood Species into Cross Timbers Forests of North-Central Oklahoma, USA" Forests 9, no. 2: 75. https://doi.org/10.3390/f9020075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.