Progress and Challenges of Protecting North American Ash Trees from the Emerald Ash Borer Using Biological Control

1
United States Department of Agriculture, Agricultural Research Service, Beneficial Insects Introduction Research Unit, Newark, DE 19713, USA
2
United States Department of Agriculture, Forest Service, Northern Research Station, Lansing, MI 48910, USA
3
Department of Environment and Conservation, University of Massachusetts, Amherst, MA 01003, USA
4
United States of Department of Agriculture, Animal and Plant Health Inspection Service, Plant Protection and Quarantine, Science and Technology, Buzzards Bay, MA 02542, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(3), 142; https://doi.org/10.3390/f9030142
Submission received: 11 February 2018
/
Revised: 9 March 2018
/
Accepted: 11 March 2018
/
Published: 15 March 2018
(This article belongs to the Special Issue Understanding and Managing Emerald Ash Borer Impacts on Ash Forests)
Abstract
:After emerald ash borer (EAB), Agrilus planipennis Fairmaire, was discovered in the United States, a classical biological control program was initiated against this destructive pest of ash trees (Fraxinus spp.). This biocontrol program began in 2007 after federal regulatory agencies and the state of Michigan approved release of three EAB parasitoid species from China: Tetrastichus planipennisi Yang (Eulophidae), Spathius agrili Yang (Braconidae), and Oobius agrili Zhang and Huang (Encyrtidae). A fourth EAB parasitoid, Spathius galinae Belokobylskij (Braconidae) from Russia, was approved for release in 2015. We review the rationale and ecological premises of the EAB biocontrol program, and then report on progress in North American ash recovery in southern Michigan, where the parasitoids were first released. We also identify challenges to conserving native Fraxinus using biocontrol in the aftermath of the EAB invasion, and provide suggestions for program improvements as EAB spreads throughout North America. We conclude that more work is needed to: (1) evaluate the establishment and impact of biocontrol agents in different climate zones; (2) determine the combined effect of EAB biocontrol and host plant resistance or tolerance on the regeneration of North American ash species; and (3) expand foreign exploration for EAB natural enemies throughout Asia.
1. Introduction
The movement of forest insects and plant pathogens, caused by the rapidly expanding global economy, poses one of the greatest threats to the ecological sustainability of forested ecosystems throughout the world [1,2,3]. Despite efforts to combat this problem through improved regulatory controls in international trade, the accidental introduction of non-native forest pests in wood packaging materials such as pallets and dunnage, as well as commodities such as nursery stock, lumber, and manufactured goods, continues [4,5,6,7]. Although a relatively small proportion of these introduced species become serious invasive pests in their invaded regions, increasing numbers of forest insects and diseases are devastating natural and urban forests worldwide [8,9].
The most recent and notable example of a destructive invasive insect damaging forests in North America is the emerald ash borer (EAB), Agrilus planipennis Fairmaire (Coleoptera: Buprestidae), introduced from Asia during the 1990s [10,11,12,13,14]. This phloem-feeding beetle attacks ash trees (Fraxinus; Oleaceae) and was discovered as the cause of ash tree mortality in southeast Michigan, USA and nearby Ontario, Canada in 2002. Over the next several years, EAB was discovered throughout the region and well beyond, being spread primarily by human-mediated transport of infested ash materials such as firewood, nursery stock, and lumber [15,16,17]. Consequently, early attempts to eradicate EAB in North America were abandoned, and research, development, and implementation of EAB-management strategies were expanded.
Biological control is now the primary management tool developed to suppress EAB densities in forested ecosystems, thereby conserving or protecting the surviving and regenerating ash trees [18,19,20], whereas systemic insecticides are available to protect high-value ash, mainly landscape trees in urban forests [21,22]. Long-term sustainability of native ash species may also require the development of EAB-resistant or tolerant ash genotypes [23,24]. An earlier review article described progress in developing EAB biocontrol program in the U.S such as foreign exploration for natural enemies in EAB’s native range Asia, host specificity testing and risk assessment for the introduced biocontrol agents, the basic biology of both introduced biocontrol agents and native North American natural enemies, and an overview of EAB biocontrol releases and research through 2014 [20]. The focus of the present paper is an overview of progress and challenges in developing, implementing, and evaluating efforts to manage EAB in forested areas using biological control. Specifically, this paper first discusses the rationale for selecting EAB as a target and the ecological premises for biocontrol, and then highlights not only recent progress made in EAB biocontrol, but also the challenges in implementing biocontrol as an EAB management tool for the conservation of North American ash species. Finally, it proposes potential solutions to overcome these challenges, including the need for expanded long-term research on EAB biocontrol as this pest continues spreading throughout North America.
2. Rationale for Selection of Emerald Ash Borer as Target for Biological Control
Emerald ash borer is a specialist herbivore attacking primarily species of Fraxinus in Asia including China, the Korean Peninsula, and the Russian Far East [25]. Asian ash species are more resistant to EAB than are North America species [24]. The high densities of EAB feeding in the phloem of North American ash cause tree mortality within five to seven years of EAB’s invasion of new locations [26,27,28,29,30,31,32]. As EAB spreads further south in the United States, it was also found attacking another native tree species in the family Oleaceae, the white fringetree (Chionanthus virginicus L.), which is commonly planted as an ornamental in eastern states [33].
Ash trees are widely distributed and highly valued in the deciduous forests of North America [34]; however, the arrival of EAB from Asia has greatly reduced the abundance of many species of ash trees in the invaded regions in the U.S. [35]. There are 16 species of Fraxinus native to North America, each species adapted to different ecological habitats across a range of climates zones, soil types, and moisture gradients, with many species in western states having limited geographical distributions [36,37,38]. Ash trees serve as food, cover, nesting sites, and habitat for mammals, birds, arthropods, and other organisms [39,40]. The earliest infestation of EAB in southeast Michigan resulted in mortality of 99% of healthy overstory ash trees in some infested forests, demonstrating the potential of EAB to functionally extirpate ash trees from the continent [29]. Emerald ash borer has since spread to 32 states and three Canadian provinces and killed hundreds of millions of ash trees in both urban and forested areas [41,42]. As a consequence, the six species of Fraxinus endemic to eastern North America are listed as critically endangered by the International Union for Conservation of Nature: white ash F. americana L., Carolina ash F. caroliniana Mill., black ash F. nigra Marshall, green ash F. pennsylvanica Marshall, pumpkin ash F. profunda (Bush) Bush, and blue ash F. quadrangulata Michx [43]. The loss of ash diversity and abundance in natural forests in the earliest-invaded regions (e.g., Midwestern and Mid-Atlantic States, USA) has already harmed native plants and ash-dependent invertebrates, and altered nutrient cycling and other ecological processes [40,44,45,46,47,48].
Although the environmental and ecological impacts of EAB on the diverse forested ecosystems of North America are not fully understood, several estimates of its economic impacts have been made. In natural forests and timberlands of the United States, more than 7.55 billion timber-sized ash trees were valued at more than $282 billion [35]. Moreover, ash trees were widely planted as landscape trees in urban forests, and an estimate for the undiscounted value of these trees in the United States ranged from $20–60 billion soon after EAB’s discovery [11]. A cost projection of EAB in just 25 northeastern communities of the United States for only one decade (2009–2019) to treat, remove, and replace landscape ash was $25 billion [49], making EAB the most destructive and costly wood-boring insect to invade the United States [50].
All evidence associated with the invasion of the United States and Canada by EAB demonstrates that this invasive insect is driving ecological degradation in the forests of North America, and taking no action against EAB is not a sensible or responsible option. Initial efforts by regulatory agencies focused on the eradication of incipient EAB populations by the creation of an ash-free zones in and around newly detected infestations [51], while imposing quarantine regulations to restrict the movement of firewood of all hardwood species and materials of the genus Fraxinus [52,53]. Although EAB- and ash-quarantine regulations remain in place, efforts to eradicate EAB were abandoned as regulatory agencies in the United States and Canada determined that eradication of EAB was not possible [13,54]. Subsequently, efforts have shifted to developing biological control-based pest management tools and strategies to slow the spread and reduce densities of EAB using conventional and biological controls, and to develop varieties of Fraxinus tolerant or resistant to EAB [24,55,56,57].
3. The Role of Natural Enemies in Suppressing EAB in Its Native Range
Little was known about the biology and natural enemy complexes of EAB in Asia before the foreign exploration work for development of a classical biological control program against EAB in the United States [18,19,20]. In 2003, researchers began foreign exploration for EAB natural enemies in northeastern China, resulting in the discovery of four hymenopteran parasitoid species: (1) Sclerodermus pupariae Yang and Yao (Bethylidae), an ectoparasitoid of larvae, prepupae, and pupae [58,59]; (2) Spathius agrili Yang (Braconidae), an ectoparasitoid of late-instar larvae [26,60]; (3) Tetrastichus planipennisi Yang (Eulophidae), an endoparasitoid of late-instar larvae [26,61]; and (4) Oobius agrili Zhang and Huang (Encyrtidae), an egg parasitoid [62]. Subsequent EAB natural enemy surveys in the Russian Far East from 2008 to 2012 led to the discovery of three additional species of hymenopteran parasitoid: (5) Spathius galinae Belokobylskij and Strazanac (Braconidae), an ectoparasitoid of late-instar larvae [63,64]; (6) Atanycolus nigriventris Vojnovskaja-Krieger (Braconidae), an ectoparasitoid of late-instar larvae [63,64]; and (7) Oobius primorskyensis Yao and Duan (Encyrtidae), an egg parasitoid [65]. In a more recent EAB natural enemy survey in northeastern China, two species of predacious Coleoptera were found attacking late-instar larvae and pupae of EAB: (8) Tenerus sp. (Cleridae); and (9) Xenoglena quadrisignata Mannerheim (Trogossitidae) [66].
Ecological studies at field sites in northeast China and the Russian Far East revealed these insect natural enemies cause high mortality of EAB eggs and larvae and play a critical role in suppressing EAB densities in forested areas of Asia [26,64,66,67]. The abundance and contribution of individual species to EAB control varied by geographic region (Table 1). For example, S. galinae is the dominant EAB larval parasitoid in the Russian Far East, causing up to 63% larval parasitism in some stands, but it has not been observed in China [64]. In contrast, T. planipennisi, is the dominant EAB larval parasitoid in northeast China, causing an average of 40% larval parasitism, but it is observed less frequently in the Russian Far East and further south in Beijing, but never in our most southern survey site in Tianjin, China where S. agrili is more abundant [26,64,66,67]. To date, O. agrili is the only EAB egg parasitoid collected consistently in China where it was found parasitizing 12–62% of EAB eggs [67]. Oobius primorskyensis, the only parasitoid found attacking EAB eggs in the Russian Far East, caused about 23–44% egg parasitism (JJD, unpublished data). Currently, studies are lacking on the ecological factors that determine the structure of EAB parasitoid assemblages and dominance of different species in different regions of the beetles’ native range. We suspect that climatic factors such as temperature and photoperiod, as well as synchronization of EAB and parasitoid phenology in different geographic regions, may have led to differences in the parasitoid assemblages and dominance. Nevertheless, our current knowledge of EAB parasitoid complexes in Asia strongly indicates that the success of EAB biological control in North America will require the introduction and successful establishment of a variety of species and genotypes of EAB parasitoids collected from different climatic regions of Asia.
4. Development of an EAB Biological Control Program in North America
To facilitate implementation of environmentally sound biological control programs, the North American Plant Protection Organization (NAPPO), with members from Canada, Mexico, and the United States, provides guidelines to petition for the release of non-indigenous entomophagous biocontrol agents in member countries [68]. The NAPPO regional standards, developed to analyze the risks and benefits of implementing a biocontrol program, are based on those developed by the International Plant Protection Convention of the Food and Agriculture Organization of the United Nations [69]. Petitions for the release of biocontrol agents are currently reviewed by Agriculture & Agri-Food Canada’s (AAFC) Biological Control Review Committee (BCRC). With experts from each member country, BCRC evaluates biocontrol petitions and makes release recommendations to United States Department of Agriculture, Animal and Plant Health Inspection Service (USDA APHIS) in the United States and to Canadian Food Inspection Agency (CFIA) in Canada. Authority to release biocontrol agents in the United States may be granted by USDA APHIS after posting on the federal register, consideration of public comments, a risk-benefit analysis, and state concurrence; the release of entomophagous biocontrol agents in Canada is coordinated by AAFC and CFIA [68,69,70].
Of the Asiatic natural enemies discovered during foreign exploration for EAB natural enemies, three EAB parasitoid species from China (T. planipennisi, S. agrili and O. agrili) were proposed for introduction, and after extensive host range testing and safety evaluation, a petition for their release was approved in the United States in 2007 [71]. In Canada, releases of T. planipennisi and S. agrili were approved by CFIA in 2013 and of O. agrili in 2015. However, S. agrili has not been released in Canada because its sustained establishment is not confirmed in the United States. Two additional species of insect natural enemy collected from EAB in the Russian Far East were proposed for release in the United States in 2014: the larval ectoparasitoid S. galinae [72] and the egg parasitoid O. primorskyensis (JJD, unpublished data). While the petition for release of S. galinae as an EAB biocontrol agent was approved by USDA APHIS in the United States in 2015 and by CFIA in Canada in 2017 [53], BCRC recommended O. primorskyensis not be approved for release in the United States until reconsideration of the petition after additional research, risk-benefit analysis, and resubmission. The introduction of A. nigriventris from the Russian Far East was also considered, but difficulties in maintaining a viable colony of this species under quarantine laboratory conditions prevented researchers from conducting host range studies for further evaluation (JJD, unpublished data). The host ranges of the hymenopteran parasitoid S. pupariae and the two species of predacious beetle (Tenerus sp. and X. quadrisignata), were deemed by researchers as too broad, and they were not considered further as potential biocontrol agents.
The safety of Asiatic parasitoids petitioned for environmental releases in North America was assessed with data collected from both field surveys of other wood-boring insects in the parasitoids’ native ranges (China and Russian Far East) and laboratory testing with Asian and North American wood-boring and other insects. Data from these studies show that host specificity of the released Asiatic parasitoids is highly constrained by the close phylogenetic proximity of potential nontarget hosts to EAB [19,71,72,73,74]. Field data from China and the Russian Far East show that these parasitoids do not attack other wood-boring insects in ash, such as bark beetles (Scolytidae) or longhorned beetles (Cerambycidae) [73,74]. However, host specificity studies in the laboratory further show that three of these introduced parasitoids—O. agrili, S. agrili, and S. galinae—do attack some Asian and North American species of Agrilus (Table 2). In contrast to the attack on EAB, however, their attack rate is lower on these potentially susceptible non-target Agrilus spp., even under laboratory conditions, which promote maximum parasitism [19,64,71,72,73,74]. Based on both laboratory and field host- range studies, the predicted non-target impact from introduction of these Asiatic parasitoids for EAB biocontrol, if any, would be a low level of parasitism of some non-target Agrilus species in North America.
There are approximately 800 Agrilus species in North America, with about 175 species in the United States [75,76]. Based on the results of these host-range studies, it is possible that the introduced EAB parasitoids may occasionally attack some of the non-target Agrilus species in North America; however, recent host-finding studies show that some EAB parasitoids are attracted to volatiles from ash trees [74,77], indicating a strong affinity to Fraxinus, their host’s food plants. Thus, it can be reasonably predicted that the level of attack on non-ash feeding Agrilus species, if any, would be limited. Field surveys of non-target insects associated with ash trees following field releases of introduced EAB parasitoids in Michigan and Maryland found no evidence of non-target attack from these introduced parasitoids [45]. In contrast, arthropod diversity associated with ash trees is significantly reduced because of the EAB invasion in Maryland [78]. The parasitoids introduced from Asia were selected for high EAB-host specificity, and in the aftermath of the EAB invasion of North America, the resulting conservation of Fraxinus and recovery of forests will produce many desirable ecological benefits [79,80].
5. Introduction and Establishment of EAB Biocontrol Agents
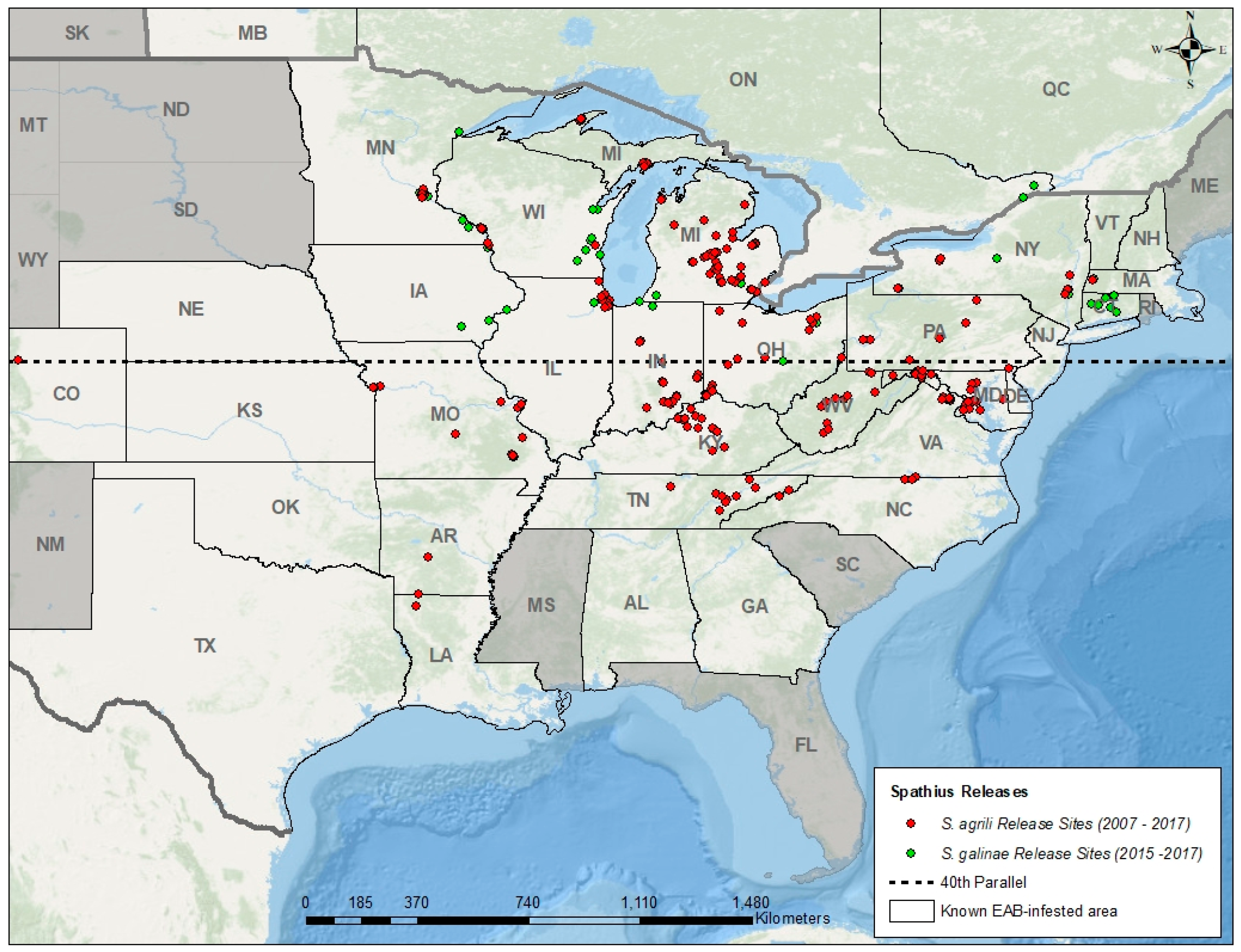
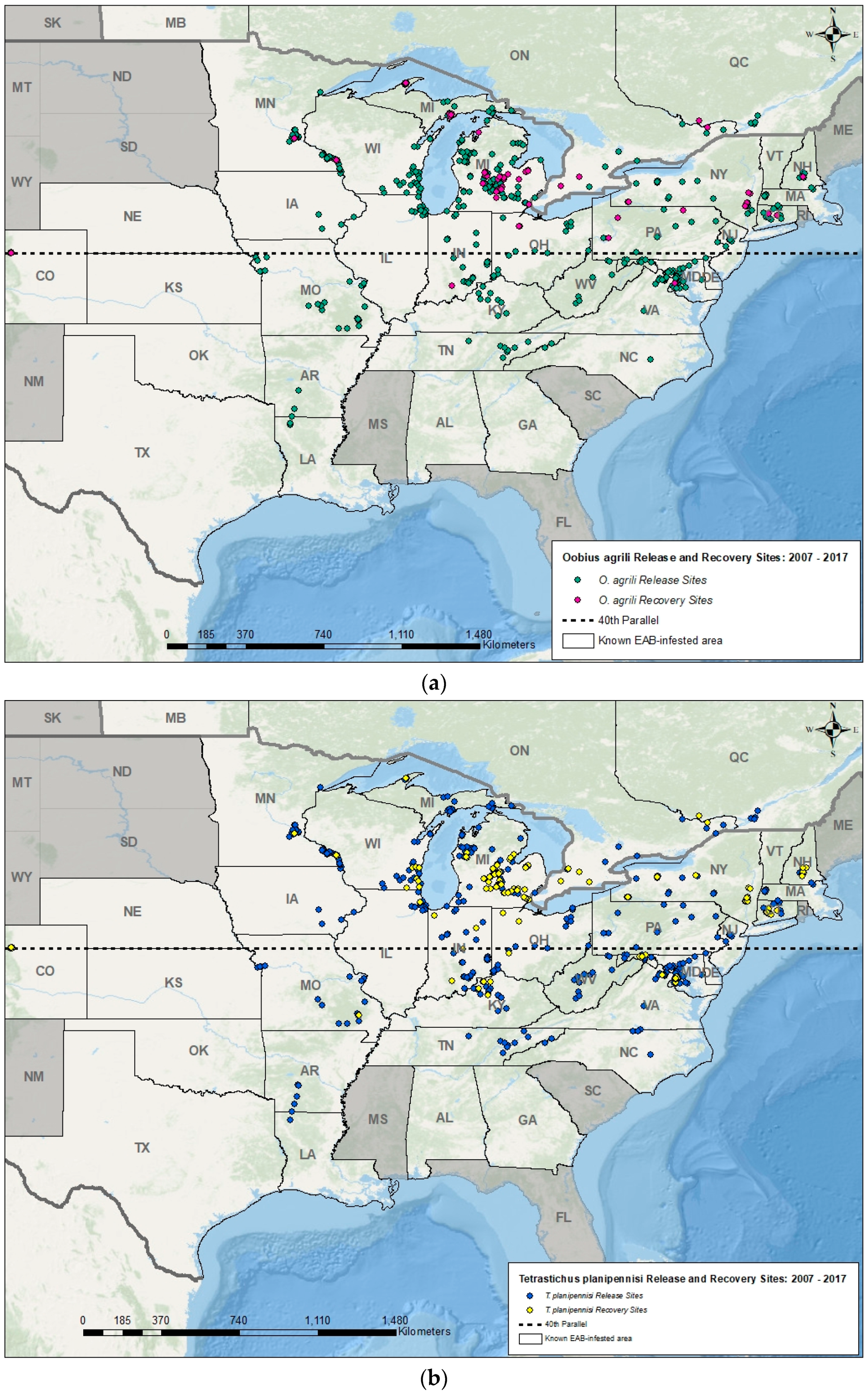
In 2007, after USDA APHIS issued permits for the environmental release of O. agrili, S. agrili, and T. planipennisi in Michigan, small numbers (a few hundred per species) were laboratory-reared and released at a few sites [18,81,82]. In subsequent years, larger numbers of parasitoids were released (tens of thousands) in additional states after USDA APHIS’ EAB biocontrol mass-rearing facility in Brighton, Michigan became operational in 2010 [19,20,83]. To date, more parasitoid releases and data on recovery is ongoing in regions with more ash trees, a longer history of EAB, and where researchers, regulatory agencies, or land managers are actively involved in research or management of EAB using biocontrol. By the end of the 2017 field season, parasitoids had been released in 27 of 32 United States and two of three Canadian provinces invaded by EAB (Figure 1 and Figure 2).
In many regions of the United States, the three Chinese biocontrol agents have been recovered from EAB larvae and eggs one year after their release, indicating successful reproduction and overwintering in the target host. However, only O. agrili (Figure 2a) and T. planipennisi (Figure 2b) are consistently recovered two or more years after their last release, and these two species are now considered established and spreading naturally beyond their initial release sites [84]. By 2013, however, researchers found that S. agrili was not establishing in northern regions of the United States, and release of this species was subsequently restricted to EAB infestations south of the 40th parallel [83] (Figure 1). To date, the establishment of S. agrili has been confirmed at only one site in Maryland (~38th parallel), 7 years after release and at a low rate of parasitism (JJD, unpublished data). Releases of S. galinae began in 2015 north of the 40th parallel, and although it is too soon to confirm establishment, the results of ongoing research on EAB natural enemies in Michigan, Connecticut, Massachusetts, and New York suggest this parasitoid is establishing and spreading [85].
6. Impact of EAB Biocontrol Agents on Target Pest Populations
The question then arises whether populations of the established parasitoids can effectively reduce the invasive EAB populations to a sufficiently low level to allow for ash regeneration and recovery in the aftermath forests. The answer to this question requires long-term research and also depends on impacts of other biotic and abiotic mortality factors of EAB in the targeted forest ecosystem. For example, a population dynamics model parameterized with observed larval and egg parasitism rates (~60%) in Asia, showed that natural enemies in Asia can quickly reduce EAB populations (i.e., with a net population growth rate < 1) when accompanied by moderate to high levels of host plant resistance with no predation from avian predators [86]. When accompanied by heavy woodpecker predation (~60%) in North America (e.g., [87,88,89]), an addition of ~35% of larval parasitism rate is sufficient to reduce the EAB population growth rate to <1, even with limited levels of host tree resistance or tolerance [86,90]. Moreover, other factors are periodically important, such as mortality caused by fungal entomopathogens [91,92] or cold winter temperatures [93].
In the same line of analysis, key abiotic factors such as temperature can also affect the efficacy of EAB biocontrol. For example, there are regional differences in the EAB life cycle because of variation of the heat accumulation in different geographic regions. In warmer climates of the southern United States, EAB eggs and larvae develop faster, thereby reducing exposure times to egg and larval parasitoids and causing EAB population growth rates to increase. In addition, this shortened EAB life cycle may also result in asynchrony of EAB egg and larval stages with adult parasitoid phenology, causing failure of establishment, or reduced impacts of natural enemies on EAB population densities. Consequently, the population-level impact of the introduced EAB biocontrol agents, in the southern United States, may be reduced by the climatic condition that favor a shortened (one year) EAB life cycle. Thus, foreign exploration for EAB natural enemies is needed in southern Asia for biocontrol of EAB in southern regions of the United States.
Data collected recently from a long-term study conducted in Michigan may provide us with some insights into the population-level impact of these introduced biocontrol agents. The long-term study consisted of six forested sites in southern Michigan, each comprised of a release and non-release control plot, which were established between 2007 and 2010. At each release plot, small numbers of adult O. agrili, S. agrili, and T. planipennisi were released, and in subsequent years, infested ash trees are being sampled to estimate EAB egg and larval parasitism, and other causes of larval mortality [81,82].
During the first five years after release of the EAB parasitoids at these study sites, EAB egg parasitism by O. agrili averaged ~1 to 4% from 2008–2011 and then increased to ~28% by 2014 in release plots. The natural spread of O. agrili from the release plots to the control plots was slow and somewhat variable between sites [82]. Overall, the impact of O. agrili in suppressing EAB population growth, as well as the natural spread rate of this biocontrol agent, has yet to be determined, because sampling EAB eggs (1 mm in diameter and cryptically colored) from ash bark layers and crevices is labor intensive and difficult to standardize [82,94]. Moreover, parasitism of EAB eggs by O. agrili is patchy, thus more intensive sampling is needed to find this tiny parasitoid and quantify its impact on EAB population dynamics. Despite these challenges, researchers are confirming the establishment and relatively slow spread of O. agrili in this and other regions of North America (Figure 2a).
In contrast to the sampling of EAB eggs, sampling EAB for larval parasitism is done by debarking live, infested ash trees, a relatively simple and reproducible method. Using this approach, average larval parasitism by T. planipennisi was ~1 to 6% from 2008–2011 and increased to ~30% by 2014 in both the release and control plots [81,95,96]. As more recovery work is done in this and other regions, researchers are finding a rapid spread of T. planipennisi across EAB-infested sites (Figure 2b). More recent life table analyses after seven years of data collection from these six study sites revealed that T. planipennisi contributed significantly to the reduction of net EAB population growth rates approximately four years after its initial release [96]. Moreover, with additional larval mortality from local natural enemies of wood boring insects, such as woodpeckers and native parasitoids (primarily braconids in the genus Atanycolus) [97], the resource-adjusted EAB larval density (per m2 of live phloem tissues) declined ~90% in infested ash trees at both the release and control plots between 2009 and 2014 [96,98]. The decline in the resource-adjusted EAB density may also be attributed in part to the general collapse of EAB populations following widespread mortality of the overstory ash trees. Depletion of host tree resources in a local area would cause EAB adults at some point to disperse in search of more abundant hosts [99,100]. However, many small ash trees and saplings, ranging in size from 2.5- to 15-cm Diameter at Breast Height (DBH)) are still abundant and susceptible to EAB infestation in the study sites [79,86]. For the surviving ash trees, the pest pressure they now experience is reduced, increasing prospects for their survival and reproduction [79,86].
The results from EAB field studies in the United States and Asia reveal that larval parasitism rates by T. planipennisi are inversely correlated ash tree diameter with 95% of larval parasitism in ash trees < 16-cm DBH [26,45,67,101]. This can be attributed to the relatively short ovipositor of T. planipennisi (average 2- to 2.5-mm long), limiting its ability to reach EAB larvae under the thick bark on lower boles of large-diameter ash trees [101]. In the same study, a larger parasitoid species (Atanycolus) with a longer ovipositor (average 4- to 6-mm long), parasitized EAB larvae in ash trees up to 57.4-cm DBH. It has been shown that T. planipennisi is important in protecting ash saplings and basal sprouts (2- to 6-cm DBH) from EAB in post-invasion recovering forests [79,80]. However, the protection of ash trees as they mature, will require establishment of the larger EAB biocontrol agent, S. galinae, which has a longer ovipositor (average 4- to 6-mm long) and capable of parasitizing EAB larvae in large-diameter ash trees [64,102]. Since releases of S. galinae began in several northern states in 2015, it appears to be establishing and spreading, and researchers will continue monitoring its impacts on EAB population dynamics and the health of large-diameter ash trees at study sites.
In theory, highly effective egg parasitoids from EAB’s native range may protect all size-class ash trees against EAB, as they can kill the pest before its larvae bore into the ash phloem to feed. However, the current level of egg parasitism by O. agrili (<29%) by itself is not sufficient to protect ash trees. Introduction of a second species of EAB egg parasitoid, O. primorskyensis, may enhance egg parasitism in some regions of North America and improve ash tree survival [103].
7. Ash Recovery and Regeneration after EAB invasion with Biological Control
Evidence gathered in the native range of EAB has shown that EAB outbreaks in northeastern Asia are rare events in natural forests, and outbreaks occur primarily in isolated plantations and urban plantings of mostly North American ash species (F. pennsylvanica, F. americana, F. velutina) [26,64,86,104]. Even if EAB can occasionally cause significant ash mortality in urban plantings or plantations of North American ash in Asia, no widespread outbreaks comparable to those observed in North American forests have been recorded in forested regions of Asia [26,64]. In addition, large, relatively healthy North American ash trees, mainly F. pennyslvanica and F. americana, have been observed in forested parks in China and urban areas in the Russian Far East [26,64,67]. It is plausible that EAB parasitoids in these regions may be protecting the more susceptible North American ash species. This protection may occur at two different phases. First, saplings or trees of susceptible ash species planted in Asia maybe colonized at low levels of EAB because there are fewer EAB founders coming from the resistant native ash species in Asia, thereby delaying EAB population increase in these susceptible ash trees or saplings. Second, the abundance and diversity of EAB parasitoids in their native range may facilitate a more rapid numerical response to incipient infestations of EAB in the susceptible ash saplings or trees, resulting in direct protection at relatively low EAB densities. In post-EAB invaded forests of North America, ash trees are much scarcer than in forests prior to invasion, and established populations of the introduced EAB parasitoids may conserve surviving native ash by moderating the frequency and amplitude of future EAB outbreaks.
The EAB invasion of forests in southeast Michigan, during the 1990s, resulted in nearly 100% mortality of overstory ash trees by 2010 [29,105]. The potential for recovery of the ash canopy was assessed in this region from 2007 to 2009, and abundant regeneration of smaller height class ash trees, mainly F. americana and F. pennsylvanica, were found. However, young 1–2 year ash seedlings were much less common, and the lack of new seedlings was traced to a depleted seed bank, as few or no nearby mature ash trees existed to provide seed [29,105]. However, the results of a more recent study of regenerating F. pennsylvanica in this region reported abundant seed production on surviving mature ash trees during mast years, as well as on sexually mature small ash trees and basal sprouts regenerating from top-killed trees, suggesting a significant, though greatly reduced, pool of ash trees in this region [106]. In a separate study in southeast Michigan where the establishment and spread of T. planipennisi is now confirmed, densities of ash and other native saplings were higher and densities of weedy species lower in closer proximity to study sites where more parasitoids were released [80]. These results suggest that protection of ash saplings by T. planipennisi favors the recruitment of native woody species over weedy species in gaps as these forests recover from loss of the overstory ash canopy in the aftermath of the EAB invasion.
Researchers also estimated the abundance and condition of ash saplings and trees at the six long-term EAB biocontrol study sites in southern Michigan, where the sustained establishment T. planipennisi and O. agrili have been documented for nearly a decade [79]. Results of this study showed that healthy ash saplings (400–1600 per hectare) and young trees (200–900 per hectare) remained in these study sites, despite formerly high EAB densities that resulted in loss of most overstory ash trees by 2010. In addition, life table analysis of EAB population dynamics at these sites indicates that the net population growth rate of EAB is near or below replacement levels, and that the introduced biocontrol agent T. planipennisi reduced the pest’s net population growth rates at these sites by over 50%. These findings strongly indicate that the introduced EAB parasitoids can provide significant biocontrol services, enhancing ash survival, and promoting forest recovery in North America [79,80,85].
8. Conclusions
Following its accidental introduction into the United States in the 1990s, EAB continues to spread and degrade ash communities and forested ecosystems in North America. The EAB Biocontrol Program, which started over a decade ago via the introduction and establishment of co-evolved natural enemies from the pest’s native range, appears to hold promise for forests of northern regions of North America. This program has documented establishment of the egg parasitoid O. agrili and the larval parasitoid T. planipennisi, both introduced from China, in EAB populations at most release sites in northern United States and southern Canada, where surveys to document parasitoid establishment are ongoing. While the role of O. agrili in reducing EAB population growth requires continued evaluation, the larval parasitoid T. planipennisi has been shown to play a significant role in protecting ash saplings and smaller trees (DBH < 12 cm) in aftermath forests in Michigan [79,80,96]. The suppression of EAB densities is likely to spread geographically as populations of O. agrili and T. planipennisi increase and spread to new areas, protecting the regenerating ash saplings and young trees. To protect growing and surviving ash trees, however, more widespread releases and successful establishment of S. galinae, the largest of the EAB biocontrol agents, are needed. As EAB continues spreading through the southern and western United States, we recommend expanding EAB biocontrol research to: (1) quantify the impacts of EAB biocontrol on ash and other native tree species as forests recover in the aftermath of EAB in northern regions; (2) develop parasitoid release methods for more widespread, remote, or larger ash stands; (3) expand research on synergistic effects of EAB biocontrol and ash resistance or tolerance to EAB in native North American ash species; (4) determine parasitoid establishment in EAB populations in warmer climates; and (5) explore different regions of Asia for EAB natural enemies adapted to climate zones similar to those in the southern and western United States where EAB is invading. Over many decades, it is reasonable to assume that a diverse complex of mortality factors and lower ash density will reduce both the frequency and intensity of EAB outbreaks, permitting the growth, survival, reproduction, and conservation of Fraxinus species.
Acknowledgments
The authors thank Deborah Miller (USDA FS, Lansing, MI), Richard Reardon (USDA FS, Morgantown, WV, USA), Jonathan Lelito and Benjamin Slager (USDA APHIS, Brighton, MI, USA), David Jennings and Paula Shrewsbury (University of Maryland), Houping Liu (Pennsylvania Department of Conservation and Natural Resources), Deborah McCullough and Amos Ziegler (Michigan State University), Scott Salom (Virginia Tech University), Clifford Sadof (Purdue University), Lynne Rieske-Kinney (University of Kentucky), Melissa Fierke (State University of New York, College of Environmental Science and Forestry), Daniel Kashian (Wayne State University), Inés Ibáñez (University of Michigan), and Joseph Elkinton and Kristopher Abell (University of Massachusetts) for their collaborative work on EAB, which helped to facilitate and inspire the write-up of this manuscript. The authors also thank Travis Perkins (Michigan State University) for preparing the maps, the many entomologists, technicians, and student employees from USDA Agricultural Research Service (ARS), Forest Service (FS), and, Michigan State University, University of Maryland, University of Massachusetts, and the many state and provincial regulators and land managers carrying out EAB biocontrol research and implementing EAB biocontrol programs. The authors also greatly appreciate early reviews of this manuscript by Kim Hoelmer (USDA ARS, Newark, DE, USA) and Noah Koller (USDA ARS, E. Lansing, MI, USA) and funding provided by USDA FS, APHIS, and ARS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pimentel, D. Biological invasions of plants and animals in agriculture and forestry. In Ecology of Biological Invasions of North America and Hawaii; Mooney, H.A., Drake, J.A., Eds.; Ecological Studies 58; Springer: New York, NY, USA, 1986; pp. 149–162. [Google Scholar]
- Aukema, J.E.; McCullough, D.G.; Von Holle, B.; Liebhold, A.M.; Britton, K.; Frankel, S.J. Historical accumulation of non-indigenous forest pests in the continental United States. BioScience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C.J. The consequence of tree pests and diseases for ecosystem services. Science 2013, 342, 1235773. [Google Scholar] [CrossRef] [PubMed]
- Liebhold, A.M.; MacDonald, W.L.; Bergdahl, D.; Mastro, V.C. Invasion by exotic forest pests: A threat to forest ecosystems. For. Sci. Monogr. 1995, 41, 1–49. [Google Scholar]
- Haack, R.A. Exotic bark and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can. J. For. Res. 2006, 36, 269–288. [Google Scholar] [CrossRef]
- Haack, R.A.; Britton, K.O.; Brockerhoff, E.G.; Cavey, J.F.; Garrett, L.J.; Kimberley, M.; Lowenstein, F.; Nuding, A.; Olson, L.J.; Turner, J.; et al. Effectiveness of the international phytosanitary standard ISPM15 on reducing wood borer infestation rates in wood packaging material entering the US. PLoS ONE 2014, 9, e96611. [Google Scholar] [CrossRef] [PubMed]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Soc. Am. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.; Fitter, A. The varying success of invaders. Ecology 1996, 77, 1661–1666. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Liebhold, A.M. Ecology of forest insect invasions. Biol. Invasions 2017, 19, 3141–3159. [Google Scholar] [CrossRef]
- Haack, R.A.; Jendek, E.; Liu, H.; Marchant, K.R.; Petrice, T.R.; Poland, T.M.; Ye, H. The emerald ash borer: A new exotic pest in North America. Newsl. Mich. Entomol. Soc. 2002, 47, 1–5. [Google Scholar]
- Federal Register. Emerald Ash Borer; Quarantine and Regulations. 2003; 7 CFR Part 301 [Docket Number 02-125-1]. Available online: https://www.federalregister.gov/documents/2003/10/14/03-25881/emerald-ash-borer-quarantine-and-regulations (accessed on 18 December 2017).
- Cappaert, D.L.; McCullough, D.G.; Poland, T.M.; Siegert, N.W. Emerald ash borer in North America: A research and regulatory challenge. Am. Entomol. 2005, 51, 152–165. [Google Scholar] [CrossRef]
- GAO. Invasive Forest Pests: Lessons Learned from Three Recent Infestations May Aid in Managing Future Efforts. Report of the United States Government Accounting Office. 2006; GAO-06-353. Available online: https://www.gao.gov/assets/250/249776.pdf (accessed on 5 January 2018).
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.W. Dendrochronological reconstruction of the epicentre and early spread of emerald ash borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]
- Poland, T.M.; McCullough, D.G. Emerald ash borer: Invasion of the urban forest and the threat to North America’s ash resource. J. For. 2006, 104, 118–124. [Google Scholar]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impact and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.S.; Liebhold, A.M.; Pugh, S.A.; Crocker, S.J. Regional assessment of emerald ash borer, Agrilus planipennis, impacts in forests of the Eastern United States. Biol. Invasions 2017, 19, 703–711. [Google Scholar] [CrossRef]
- Bauer, L.S.; Liu, H.P.; Miller, D.L.; Gould, J. Developing a classical biological control program for Agrilus planipennis (Coleoptera: Buprestidae), an invasive ash pest in North America. Newsl. Mich. Entomol. Soc. 2008, 53, 38–39. [Google Scholar]
- Bauer, L.S.; Duan, J.J.; Gould, J.R. Emerald ash borer Agrilus planipennis Fairmaire Coleoptera: Buprestidae. In The Use of Classical Biological Control to Preserve Forests in North America; FHTET-2013-2; Van Driesche, R., Reardon, R., Eds.; United States Department of Agriculture, Forest Service, Forest Health and Technology Enterprise Team: Morgantown, WV, USA, 2014; pp. 189–209. Available online: https://www.nrs.fs.fed.us/pubs/48051 (accessed on 30 January 2018).
- Bauer, L.S.; Duan, J.J.; Gould, J.R.; Van Driesche, R.G. Progress in the classical biological control of Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) in North America. Can. Entomol. 2015, 147, 300–317. [Google Scholar] [CrossRef]
- Sadof, C.S.; Hughes, G.P.; Witte, A.R.; Peterson, D.J.; Ginzel, M.D. Tools for staging and managing emerald ash borer in the urban forest. Arboric. Urban For. 2017, 43, 15–26. [Google Scholar]
- Mercader, R.J.; McCullough, D.G.; Storer, A.J.; Bedford, J.; Poland, T.M.; Katovich, S. Evaluation of the potential use of a systemic insecticide and girdled trees in area wide management of the emerald ash borer. For. Ecol. Manag. 2015, 350, 70–80. [Google Scholar] [CrossRef]
- Rigsby, C.M.; Showalter, D.N.; Herms, D.A.; Koch, J.L.; Bonello, P.; Cipollini, D. Physiological responses of emerald ash borer larvae to feeding on different ash species reveal putative resistance mechanisms and insect counter-adaptations. J. Insect Physiol. 2015, 78, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Villari, C.; Herms, D.A.; Whitehill, J.G.A.; Cipollini, D.; Bonello, P. Progress and gaps in understanding mechanisms of ash tree resistance to emerald ash borer; a model for wood-boring insects that kill angiosperms. New Phytol. 2016, 209, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.; Bauer, L.S.; Poland, T.M.; Haack, R.A.; Cognato, A.I.; Smith, J.J. Genetic analysis of emerald ash borer (Agrilus planipennis Fairmaire) populations in Asia and North America. Biol. Invasions 2011, 13, 2869–2887. [Google Scholar] [CrossRef]
- Liu, H.; Bauer, L.S.; Gao, R.; Zhao, T.; Petrice, T.R.; Haack, R.A. Exploratory survey for the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae), and its natural enemies in China. Great Lakes Entomol. 2003, 36, 191–204. [Google Scholar]
- Eyles, A.; Jones, W.; Riedl, K.; Cipollini, D.; Schwartz, S.; Chan, K.; Herms, D.A.; Bonello, P. Comparative phloem chemistry of Manchurian Fraxinus mandshurica and two North American ash species Fraxinus americana and Fraxinus pennsylvanica. J. Chem. Ecol. 2007, 33, 1430–1448. [Google Scholar] [CrossRef] [PubMed]
- Rebek, E.J.; Herms, D.A.; Smitley, D.R. Interspecific variation in resistance to emerald ash borer (Coleoptera: Buprestidae) among North American and Asian ash (Fraxinus spp.). Environ. Entomol. 2008, 37, 242–246. [Google Scholar] [CrossRef]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.S.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by emerald ash borer (Agrilus planipennis). Biol. Invasions 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Tanis, S.R.; McCullough, D.G. Differential persistence of blue ash (Fraxinus quadrangulata) and white ash (Fraxinus americana) following emerald ash borer (Agrilus planipennis) invasion. Can. J. For. Res. 2012, 42, 1542–1550. [Google Scholar] [CrossRef]
- Tanis, S.R.; McCullough, D.G. Host resistance of five Fraxinus species to Agrilus planipennis (Coleoptera: Buprestidae) and effects of paclobutrazol and fertilization. Environ. Entomol. 2015, 41, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.S.; Brown, J.P.; Long, R.P. Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol. Invasions 2013, 15, 371–383. [Google Scholar] [CrossRef]
- Cipollini, D. White fringetree as a novel larval host for emerald ash borer. J. Econ. Entomol. 2015, 108, 370–375. [Google Scholar] [CrossRef] [PubMed]
- USDA–FS (United States Department of Agriculture, Forest Service). Silvics of Forest Trees of the United States; Compiler; Agriculture Handbook 271; Folwells, H.A., Ed.; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1965; pp. 181–196. Available online: https://catalog.hathitrust.org/Record/001507718 (accessed on 20 December 2017).
- Nowak, D.; Crane, D.; Stevens, J.; Walton, J. Potential Damage from Emerald Ash Borer; United States Department of Agriculture, Forest Service, Northern Research Station: Syracuse, NY, USA, 2003; pp. 1–5. Available online: https://www.nrs.fs.fed.us/disturbance/invasive_species/eab/local-resources/downloads/EAB_potential.pdf (accessed on 18 December 2017).
- Eyre, F.H. (Ed.) Forest Cover Types of the United States and Canada; Society of American Foresters: Washington, DC, USA, 1980; 148p, ISBN 13: 978-0686306979. [Google Scholar]
- Harlow, W.M.; Harrar, E.S.; Hardin, J.W.; White, F.M. Textbook of Dendrology, 8th ed.; McGraw Hill Book Company: New York, NY, USA, 1996; ISBN 13: 978-0070265721. [Google Scholar]
- USDA–NRCS. Plants Profile for Fraxinus in North America. 2017. Available online: https://plants.usda.gov/core/profile?symbol=fraxi (accessed on 18 December 2017).
- Koenig, W.D.; Liebhold, A.M.; Bonter, D.N.; Hachachka, W.M.; Dicknson, J.L. Effects of the emerald ash borer on four species of birds. Biol. Invasions 2013, 15, 2095–2103. [Google Scholar] [CrossRef]
- Wagner, D.L.; Todd, K.J. New ecological assessment for the emerald ash borer: A cautionary tale about unvetted host-plant literature. Am. Entomol. 2016, 62, 26–35. [Google Scholar] [CrossRef]
- USDA–APHIS. Initial County EAB Detection Map. 2018. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/emerald_ash_b/downloads/MultiState.pdf (accessed on 7 February 2018).
- Emerald Ash Borer Information. Emerald Ash Borer Information Network. 2016. Available online: http://www.emeraldashborer.info/ (accessed on 18 December 2017).
- IUCN (International Union for Conservation of Nature). IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/ (accessed on 18 December 2017).
- Flower, C.E.; Knight, K.S.; Gonzalez-Meler, M.A. Impacts of the emerald ash borer (Agrilus planipennis) induced ash (Fraxinus spp.) mortality on forest carbon cycling and successional dynamics in the eastern United States. Biol. Invasions 2013, 15, 931–944. [Google Scholar] [CrossRef]
- Jennings, D.E.; Duan, J.J.; Bean, D.; Gould, J.R.; Kimberly, A.R.; Shrewsbury, P.M. Monitoring the establishment and abundance of introduced parasitoids of emerald ash borer larvae in Maryland, U.S.A. Biol. Control 2016, 101, 138–144. [Google Scholar] [CrossRef]
- Stephens, J.P.; Berven, K.A.; Tiegs, S.D. Anthropogenic changes to leaf litter input affect the fitness of a larval amphibian. Freshw. Biol. 2013, 58, 1631–1646. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Klooster, W.S.; Barrington, W.T.; Herms, D.A. Impacts of emerald ash borer-induced tree mortality on leaf litter arthropods and exotic earthworms. Pedobiologia 2011, 54, 261–265. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Barrington, W.T.; Hoebeke, E.R.; Herms, D.A. Vertically stratified ash-limb beetle fauna in northern Ohio. Psyche 2012, 2012, 215891. [Google Scholar] [CrossRef]
- Kovacs, F.K.; Haight, R.G.; McCullough, D.G.; Mercader, R.J.; Siegert, N.W.; Leibhold, A.M. Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecol. Econ. 2010, 69, 569–578. [Google Scholar] [CrossRef]
- Aukema, J.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.J.; Bauer, L.S.; Poland, T.M.; Windell, K. Flight performance of Agrilus planipennis (Coleoptera: Buprestidae) on a flight mill and in free flight. J. Insect Behav. 2010, 23, 128–148. [Google Scholar] [CrossRef]
- USDA–APHIS. Emerald Ash Borer Federal Regulations and Quarantine Notices. 2017. Available online: https://www.aphis.usda.gov/aphis/ourfocus/planthealth/plant-pest-and-disease-programs/pests-and-diseases/emerald-ash-borer/ct_quarantine (accessed on 30 January 2018).
- CFIA. Canadian Food Inspection Agency. Emerald Ash Borer. 2018. Available online: http://www.inspection.gc.ca/plants/plant-pests-invasive-species/insects/emerald-ash-borer/eng/1337273882117/1337273975030 (accessed on 29 January 2018).
- USDA–APHIS. Emerald Ash Borer Program Manual, Agrilus planipennis (Fairmaire), ver. 1.6. 2015. Available online: https://www.aphis.usda.gov/import_export/plants/manuals/domestic/downloads/emerald_ash_borer_manual.pdf (accessed on 7 February 2018).
- Flower, C.E.; Dalton, J.E.; Knight, K.S.; Brikha, M.; Gonzalez-Meler, M.A. To treat or not to treat: Diminishing effectiveness of emamectin benzoate tree injections in ash trees heavily infested by emerald ash borer. Urban For. Urban Green. 2015, 14, 790–795. [Google Scholar] [CrossRef]
- O’Brien, E.M. Conserving Ash (Fraxinus) Populations and Genetic Variation in Forests Invaded by Emerald Ash Borer Using Large-Scale Insecticide Applications. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2017. [Google Scholar]
- Davidson, W.; Rieske, L.K. Establishment of classical biological control targeting emerald ash borer is facilitated by use of insecticides, with little effect on native arthropod communities. Biol. Control 2016, 101, 78–86. [Google Scholar] [CrossRef]
- Wu, H.; Wang, X.Y.; Li, M.L.; Yang, Z.Q.; Zeng, F.Z.; Wang, H.Y.; Bai, L.; Liu, S.J.; Sun, J. Biology and mass rearing of Sclerodermus pupariae Yang et Yao (Hymenoptera: Bethylidae), an important ectoparasitoid of the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae) in China. Acta Entomol. Sin. 2008, 51, 46–54. [Google Scholar]
- Yang, Z.Q.; Wang, X.Y.; Yao, Y.X.; Gould, J.R.; Cao, L.M. A new species of Sclerodermus (Hymenoptera: Bethylidae) parasitizing Agrilus planipennis (Coleoptera: Buprestidae) from China, with a key to Chinese species in the genus. Ann. Entomol. Soc. Am. 2012, 105, 619–627. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Achterberg, C.V.; Choi, W.Y.; Marsh, P.M. First recorded parasitoid from China of Agrilus planipennis: A new species of Spathius (Hymenoptera: Braconidae: Doryctinae). Ann. Entomol. Soc. Am. 2005, 98, 636–642. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Yao, Y.X.; Wang, X.Y. A new species of emerald ash borer parasitoid from China belonging to the genus Tetrastichus (Hymneoptera: Eulophidae). Proc. Entomol. Soc. Wash. 2006, 108, 550–558. [Google Scholar]
- Zhang, Y.Z.; Huang, D.W.; Zhao, T.H.; Liu, H.P.; Bauer, L.S. Two new species of egg parasitoids (Hymenoptera: Encyrtidae) of wood-boring beetle pests from China. Phytoparasitica 2005, 53, 253–260. [Google Scholar] [CrossRef]
- Belokobylskij, S.A.; Yurchenko, G.I.; Strazanac, J.S.; Zaldi’var-Riveron, A.L.; Mastro, V. A new emerald ash borer (Coleoptera: Buprestidae) parasitoid species of Spathius Nees (Hymenoptera: Braconidae: Doryctinae) from the Russian Far East and South Korea. Ann. Entomol. Soc. Am. 2012, 105, 165–178. [Google Scholar] [CrossRef]
- Duan, J.J.; Yurchenko, G.; Fuester, R.W. Occurrence of emerald ash borer (Coleoptera: Buprestidae) and biotic factors affecting its immature stages in the Russian Far East. Environ. Entomol. 2012, 41, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.X.; Duan, J.J.; Hopper, K.R.; Mottern, J.L.; Gates, M.W. A new species of Oobius Trjapitzin (Hymenoptera: Encyrtidae) from the Russian Far East that parasitizes eggs of emerald ash borer (Coleoptera: Buprestidae). Ann. Entomol. Soc. Am. 2016, 106, 629–638. [Google Scholar] [CrossRef]
- Wang, X.Y.; Cao, L.M.; Yang, Z.Q.; Duan, J.J.; Gould, J.R.; Bauer, L.S. Natural enemies of emerald ash borer (Coleoptera: Buprestidae) in northeast China, with notes on two species of parasitic Coleoptera. Can. Entomol. 2016, 148, 329–342. [Google Scholar] [CrossRef]
- Liu, H.; Bauer, L.S.; Miller, D.L.; Zhao, T.; Gao, R.; Song, L.; Luan, Q.; Jin, R.; Gao, C. Seasonal abundance of Agrilus planipennis (Coleoptera: Buprestidae) and its natural enemies Oobius agrili (Hymenoptera: Encyrtidae) and Tetrastichus planipennisi (Hymenoptera: Eulophidae) in China. Biol. Control 2007, 42, 61–71. [Google Scholar] [CrossRef]
- NAPPO. (North American Plant Protection Organization) NAPPO Regional Standards for Phytosanitary Measures (RSPM). RSPM 12: Guidelines for Petition for First Release of Non-Indigenous Entomophagous Biological Control Agents. 2015. Available online: https://www.nappo.org/files/1814/4065/2949/RSPM12_30-07-2015-e.pdf (accessed on 19 January 2018).
- Mason, P.G.; Kabaluk, J.T.; Spence, B.; Gillespie, D.R. Regulation of Biological Control in Canada. In Biological Control Programmes in Canada 2001–2012; Mason, P.G., Gillespie, D.R., Eds.; CABI: Wallingford, UK, 2013; pp. 1–5. [Google Scholar]
- Montgomery, M. Understanding federal regulations as guidelines for classical biological control programs. In Implementation and Status of Biological Control of the Hemlock Woolly Adelgid; Onken, B., Reardon, R., Eds.; FHTET-2011-04; United States Department of Agriculture, Forest Service: Morgantown, WV, USA, 2011; pp. 25–40. Available online: http://www.fs.fed.us/nrs/pubs/jrnl/2011/nrs_2011_montgomery_001.pdf (accessed on 19 January 2018).
- Federal Register. Availability of an environmental assessment for the proposed release of three parasitoids for the biological control of the emerald ash borer Agrilus planipennis in the Continental United States. Fed. Regist. 2007, 72, 28947–28948. Available online: http://www.regulations.gov/#!documentDetail;D=APHIS-2007-0060-0043 (accessed on 18 December 2017).
- Federal Register. Availability of an environmental assessment for field release of the parasitoid Spathius galinae for the biological control of the emerald ash borer (Agrilus planipennis) in the contiguous United States. Fed. Regist. 2015, 80, 7827–7828. Available online: https://www.regulations.gov/docket?D=APHIS-2014-0094 (accessed on 18 December 2017).
- Duan, J.J.; Gould, J.R.; Fuester, R.W. Evaluation of the host specificity of Spathius galinae (Hymenoptera: Braconidae), a larval parasitoid of the emerald ash borer (Coleoptera: Buprestidae) in Northeast Asia. Biol. Control 2015, 89, 91–97. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Wang, X.Y.; Gould, J.R.; Wu, H. Host specificity of Spathius agrili Yang (Hymenoptera: Braconidae), an important parasitoid of the emerald ash borer. Biol. Control 2008, 47, 216–221. [Google Scholar] [CrossRef]
- Bellamy, C.L. World catalogue and bibliography of the jewel beetles (Coleoptera: Buprestidae). In Agrilinae: Agrilina through Trachyini; Pensoft Series Faunistica #79; Pensoft Publishers: Sofia, Bulgaria; Moscow, Russia, 2008; Volume 4, pp. 1932–2684. ISBN 9789546423214. [Google Scholar]
- Nelson, G.H.; Walters, G.C., Jr.; Haines, R.D.; Bellamy, C.L. A Catalog and Bibliography of the Buprestoidea of America North of Mexico; The Coleopterists Society, Special Publication: North Potomac, MD, USA, 2008; pp. 1–274. [Google Scholar]
- Johnson, T.D.; Lelito, J.P.; Raffa, K.F. Responses of two parasitoids, the exotic Spathius agrili Yang and the native Spathius floridanus Ashmead, to volatile cues associated with the emerald ash borer, Agrilus planipennis Fairmaire. Biol. Control 2014, 79, 110–117. [Google Scholar] [CrossRef]
- Jennings, D.E.; Duan, J.J.; Bean, D.; Kimberly, A.R.; Williams, G.L.; Bells, S.K.; Shurtleff, A.S.; Shrewsbury, P.M. Effects of the emerald ash borer invasion on the community composition of arthropods associated with ash tree boles in Maryland, U.S.A. Agric. For. Entomol. 2017, 19, 122–129. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.G. Emerald ash borer biocontrol in ash saplings: The potential for early stage recovery of North American ash trees. For. Ecol. Manag. 2017, 394, 64–72. [Google Scholar] [CrossRef]
- Margulies, E.; Bauer, L.; Ibanez, I. Buying time: Preliminary assessment of biocontrol in the recovery of native forest vegetation in the aftermath of the invasive emerald ash borer. Forests 2017, 8, 369. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Lelito, J.P.; Van Driesche, R.G. Establishment and abundance of Tetrastichus planipennisi (Hymenoptera: Eulophidae) in Michigan: Potential for success in classical biocontrol of the invasive emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2013, 106, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Abell, K.J.; Bauer, L.S.; Duan, J.J.; Van Driesche, R.G. Long-term monitoring of the introduced emerald ash borer (Coleoptera: Buprestidae) egg parasitoid, Oobius agrili (Hymenoptera: Encyrtidae), in Michigan, USA and evaluation of a newly developed monitoring technique. Biol. Control 2014, 79, 36–42. [Google Scholar] [CrossRef]
- USDA–APHIS/ARS/FS. USDA Animal Plant Health Inspection Service/Agricultural Research Service/Forest Service. Emerald Ash Borer Biological Control Release and Recovery Guidelines. 2016. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/emerald_ash_b/downloads/EAB-FieldRelease-Guidelines.pdf (accessed on 18 January 2018).
- MapBioControl.org. Agent Release Tracking and Data Management for Federal, State, and Researchers Releasing Biocontrol Agents for Management of the Emerald Ash Borer. 2018. Available online: http://www.mapbiocontrol.org/ (accessed on 16 January 2018).
- Duan, J.J.; Van Driesche, R.G.; Bauer, L.S.; Reardon, R.; Gould, J.; Elkinton, J.S. The Role of Biocontrol of Emerald Ash Borer in Protecting Ash Regeneration after Invasion; FHAAST-2017-02; United States Department of Agriculture, Forest Service, Forest Health Assessment and Applied Sciences Team: Morgantown, WV, USA, 2017; pp. 1–10. [Google Scholar]
- Duan, J.J.; Van Driesche, R.G.; Bauer, L.S.; Kashian, D.M.; Herms, D.A. Risk to ash from emerald ash borer: Can biological control prevent the loss of ash stands? In Biology and Control of Emerald Ash Borer; van Driesche, R.G., Reardon, R.C., Eds.; FHTET-2014-9; United States Department of Agriculture Forest Service, Forest Health Technology Enterprise Team: Morganton, WV, USA, 2015; pp. 153–163. Available online: https://www.nrs.fs.fed.us/pubs/49310 (accessed on 30 January 2018).
- Lindell, C.A.; McCullough, D.G.; Cappaert, D.; Apostolou, N.M.; Roth, M.B. Factors influencing woodpecker predation on emerald ash borer. Am. Midl. Nat. 2008, 159, 434–444. [Google Scholar] [CrossRef]
- Flower, C.E.; Long, L.C.; Knight, K.S.; Rebbeck, J.; Brown, J.S.; Gonzalez-Meler, M.A.; Whelan, C.J. Native bark-foraging birds preferentially forage in infected ash (Fraxinus spp.) and prove effective predators of the invasive emerald ash borer (Agrilus planipennis Fairmaire). For. Ecol. Manag. 2014, 313, 300–306. [Google Scholar] [CrossRef]
- Jennings, D.E.; Gould, J.R.; Vandenberg, J.D.; Duan, J.J.; Shrewsbury, P.M. Quantifying the impact of woodpecker predation on population dynamics of the emerald ash borer (Agrilus planipennis). PLoS ONE 2013, 8, e83491. [Google Scholar] [CrossRef] [PubMed]
- Jennings, D.E.; Duan, J.J.; Abell, K.J.; Bauer, L.S. Life table evaluation of change in emerald ash borer populations due to biological control. In Biology and Control of Emerald Ash Borer; Van Driesche, R.G., Reardon, R.C., Eds.; FHTET-2014-9; United States Department of Agriculture Forest Service, Forest Health Technology Enterprise Team: Morganton, WV, USA, 2015; pp. 139–151. Available online: https://www.nrs.fs.fed.us/pubs/49312 (accessed on 6 March 2018).
- Liu, H.P.; Bauer, L.S. Susceptibility of Agrilus planipennis (Coleoptera: Buprestidae) to Beauveria bassiana and Metarhizium anisopliae. J. Econ. Entomol. 2006, 99, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, L.A.; Bauer, L.S.; Houping, L.P.; Griggs, M.H.; Vandenberg, J.D. Characterization of Beauveria bassiana (Ascomycota: Hypocreales) isolates associated with Agrilus planipennis (Coleoptera: Buprestidae) populations in Michigan. Biol. Control 2010, 54, 135–140. [Google Scholar] [CrossRef]
- Wu, H.; Li, M.L.; Yang, Z.Q.; Wang, X.Y. Research on cold hardiness of emerald ash borer and its two parasitoids, Spathius agrili Yang (Hym., Braconidae) and Tetrastichus planipennisi Yang (Hym., Eulophidae). Chin. J. Biol. Control 2007, 23, 119–122. (In Chinese) [Google Scholar]
- Abell, K.J.; Bauer, L.S.; Miller, D.L.; Duan, J.J.; Van Driesche, R.G. Monitoring the establishment and flight phenology of parasitoids of emerald ash borer (Coleoptera: Buprestidae) in Michigan by using sentinel eggs and larvae. Fla. Entomol. 2016, 99, 667–672. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Van Driesche, R.G. Population responses of hymenopteran parasitoids to the emerald ash borer (Coleoptera: Buprestidae) in recently invaded areas in north central United States. BioControl 2012, 57, 199–209. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Ulyshen, M.D.; Van Driesche, R.G. Population dynamics of an invasive forest insect and associated natural enemies in the aftermath of invasion: Implications for biological control. J. Appl. Ecol. 2015, 52, 1246–1254. [Google Scholar] [CrossRef]
- Bauer, L.S.; Duan, J.J.; Lelito, J.P.; Liu, H.P.; Gould, J.R. Biology of emerald ash borer parasitoids. In Biology and Control of Emerald Ash Borer; van Driesche, R.G., Reardon, R.C., Eds.; FHTET-2014-9; United States Department of Agriculture Forest Service, Forest Health Technology Enterprise Team: Morganton, WV, USA, 2015; Chapter 6; pp. 97–112. Available online: https://www.nrs.fs.fed.us/pubs/49294 (accessed on 30 January 2018).
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Van Driesche, R.G. Natural enemies implicated in the regulations of an invasive pest: A life table analysis of the population dynamics of the invasive emerald ash borer. Agric. For. Entomol. 2014, 16, 406–416. [Google Scholar] [CrossRef]
- Mercader, R.; Siegert, N.W.; Liebhold, A.M.; McCullough, D.G. Dispersal of the emerald ash borer, Agrilus planipennis, in newly colonized sites. Agric. For. Entomol. 2009, 11, 421–424. [Google Scholar] [CrossRef]
- Siegert, N.W.; McCullough, D.G.; Williams, D.W.; Fraser, I.; Poland, T.M. Dispersal of Agrilus planipennis (Coleoptera: Buprestidae) from discrete epicenters in two outlier sites. Environ. Entomol. 2010, 39, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Abell, K.J.; Duan, J.J.; Bauer, L.S.; Lelito, J.P.; Van Driesche, R.G. The effect of bark thickness on the effectiveness of Tetrastichus planipennisi (Hymen: Eulophidae) and Atanycolus spp. (Hymen: Braconidae) two parasitoids of emerald ash borer (Coleop: Buprestidae). Biol. Control 2012, 63, 320–325. [Google Scholar] [CrossRef]
- Murphy, T.C.; Van Dreische, R.G.; Gould, J.R.; Elkinton, J. Can Spathius galinae attack emerald ash borer larvae feeding in large ash trees? Biol. Control 2017, 114, 8–14. [Google Scholar] [CrossRef]
- Larson, K.M.; Duan, J.J. Differences in the reproductive biology and diapause of two congeneric species of egg parasitoids (Hymenoptera: Encyrtidae) from northeast Asia: Implications for biological control of the invasive emerald ash borer (Coleoptera: Buprestidae). Biol. Control 2016, 101, 39–45. [Google Scholar] [CrossRef]
- Wei, X.; Reardon, D.; Wu, Y.; Sun, J.H. Emerald ash borer, Agrilus planipennis, in China: A review and distribution survey. Acta Entomol. Sin. 2004, 47, 679–685. [Google Scholar]
- Kashian, D.M.; Witter, J.A. Assessing the potential for ash canopy tree replacement via current regeneration following emerald ash borer-caused mortality on southeastern Michigan landscapes. For. Ecol. Manag. 2011, 261, 480–488. [Google Scholar] [CrossRef]
- Kashian, D.M. Sprouting and seed production may promote persistence of green ash in the presence of the emerald ash borer. Ecosphere 2016, 7, e01332. [Google Scholar] [CrossRef]
Figure 1.
Map showing known regions of North America invaded by EAB, and locations where Spathius agrili and S. galinae were released by fall 2017 [84]. In the United States, releases of S. agrili and S. galinae began in 2007 and 2015, respectively. In Canada, release of S. galinae began in 2017. In 2013, release of S. agrili was limited to EAB infestations south of the 40th parallel due to lack of establishment further north. No recovery sites for Spathius are shown because establishment of S. agrili was found at only one release site, and it is too soon to confirm establishment of S. galinae.
Figure 1.
Map showing known regions of North America invaded by EAB, and locations where Spathius agrili and S. galinae were released by fall 2017 [84]. In the United States, releases of S. agrili and S. galinae began in 2007 and 2015, respectively. In Canada, release of S. galinae began in 2017. In 2013, release of S. agrili was limited to EAB infestations south of the 40th parallel due to lack of establishment further north. No recovery sites for Spathius are shown because establishment of S. agrili was found at only one release site, and it is too soon to confirm establishment of S. galinae.
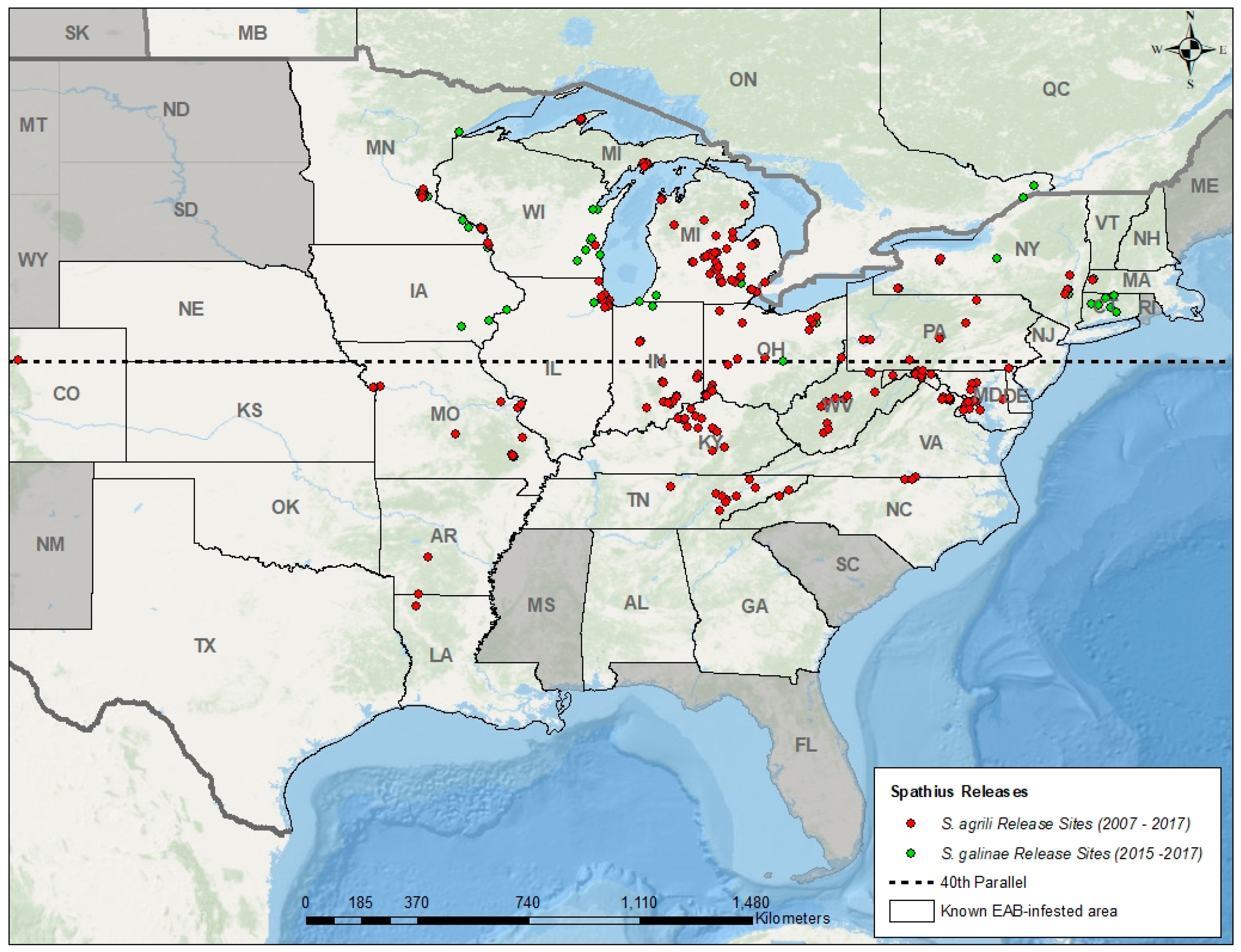
Figure 2.
Maps showing known regions of North America invaded by EAB, and the release and recovery sites for (a) O. agrili and (b) T. planipennisi, EAB biocontrol agents introduced from China in North America from 2007 to 2017, using a variety of methods [83] and documented on the EAB biocontrol geospatial database [84]. In the United States, releases of T. planipennisi and O. agrili began in 2007, and in Canada, releases of these species began in 2013 and 2015, respectively. Establishment of these two biocontrol agents in EAB populations are confirmed at many early release sites.
Figure 2.
Maps showing known regions of North America invaded by EAB, and the release and recovery sites for (a) O. agrili and (b) T. planipennisi, EAB biocontrol agents introduced from China in North America from 2007 to 2017, using a variety of methods [83] and documented on the EAB biocontrol geospatial database [84]. In the United States, releases of T. planipennisi and O. agrili began in 2007, and in Canada, releases of these species began in 2013 and 2015, respectively. Establishment of these two biocontrol agents in EAB populations are confirmed at many early release sites.


{kind=link}
{kind=link}
Table 1.
Natural enemy complexes and their observed attack rates on the emerald ash borer (EAB) in different regions of its native range in northeastern Asia.
Table 1.
Natural enemy complexes and their observed attack rates on the emerald ash borer (EAB) in different regions of its native range in northeastern Asia.
| Geographic Region | Natural Enemies | EAB Stage(s) Attacked | Rate of Attack (Parasitism or Predation) | References |
|---|---|---|---|---|
| Northeast China: Heilongjiang, Jilin, and Liaoning provinces | Oobius agrili | eggs | 12–62% | [26,62,66] |
| Oencyrtus sp. | eggs | 1–2% | [19] | |
| Tetrastichus planipennisi | 3rd and 4th instars | 3–44% | [26,61,66,67] | |
| Spathius agrili | 3rd and 4th instars | 0–13% | [26,60,66] | |
| Atanycolus spp. Foerster (Hymenoptera: Braconidae) | 3rd and 4th instars | 0–23% | [66] | |
| Xorides sp. Latreille (Hymenoptera: Ichneumonidae) | 3rd and 4th instars | 0–11% | [66] | |
| Tenerus sp. Laporte (Coleoptera: Cleridae) | JLand pupae | 0–21% | [66] | |
| Xenoglena quadrisignata Mannerheim (Coleoptera: Trogossitidae) | JL and pupae | 0–1.2% | [66] | |
| Northcentral China: Beijing and Tianjin cities | Oobius agrili | eggs | 0–4.0% | [62,66] |
| Tetrastichus planipennisi | 3rd and 4th instars | 0–7% | [26,61,66,67] | |
| Spathius agrili | 3rd and 4th instars | 44–67% | [61,66] | |
| Atanycolus sp. | 3rd and 4th instars | 0–5% | [66] | |
| Metapelma sp. Westwood (Hymenoptera: Eupelmidae) | 3rd and 4th instars | 0–4% | [66] | |
| Sclerodermus pupariae Yang and Yao (Hymenoptera: Bethylidae) | 3rd and pupae | 1–1.3% | [66] | |
| Russia: Primorsky Kray | Oobius primorskyensis | egg | 23–44% | JJD (unpublished data) |
| Tetrastichus planipennisi | 3rd and 4th instars | 0–7% | [64] | |
| Spathius galinae | 3rd and 4th instars | 0–78% | [64] | |
| Atanycolus nigriventris Vojnovskaja-Krieger (Braconidae: Braconinae) | 3rd and 4th instars | 0–55% | [64] | |
| Atanycolus sp. | 3rd and 4th instars | 0–1% | [64] |
Table 2.
Non-target insect taxa tested with the Asiatic parasitoids petitioned for environmental release in North America as EAB biocontrol agents.
Table 2.
Non-target insect taxa tested with the Asiatic parasitoids petitioned for environmental release in North America as EAB biocontrol agents.
| EAB Parasitoids from Asia | Insect Orders Tested | Insect Families Tested | Insect Species Tested | Agrilus Species Tested | The Only Non-Targets Attacked Were Agrilus Species |
|---|---|---|---|---|---|
| Oobius agrili 1 | 2 | 6 | 18 | 6 | 3 |
| Tetrastichus planipennisi 1 | 3 | 6 | 14 | 5 | 0 |
| Spathius agrili 1 | 2 | 6 | 18 | 9 | 5 |
| Spathius galinae 2 | 3 | 6 | 15 | 6 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Duan, J.J.; Bauer, L.S.; Van Driesche, R.G.; Gould, J.R. Progress and Challenges of Protecting North American Ash Trees from the Emerald Ash Borer Using Biological Control. Forests 2018, 9, 142. https://doi.org/10.3390/f9030142
AMA Style
Duan JJ, Bauer LS, Van Driesche RG, Gould JR. Progress and Challenges of Protecting North American Ash Trees from the Emerald Ash Borer Using Biological Control. Forests. 2018; 9(3):142. https://doi.org/10.3390/f9030142
Chicago/Turabian StyleDuan, Jian J., Leah S. Bauer, Roy G. Van Driesche, and Juli R. Gould. 2018. "Progress and Challenges of Protecting North American Ash Trees from the Emerald Ash Borer Using Biological Control" Forests 9, no. 3: 142. https://doi.org/10.3390/f9030142
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.