Hepatitis Delta Virus RNA Replication

Abstract
:1. Introduction
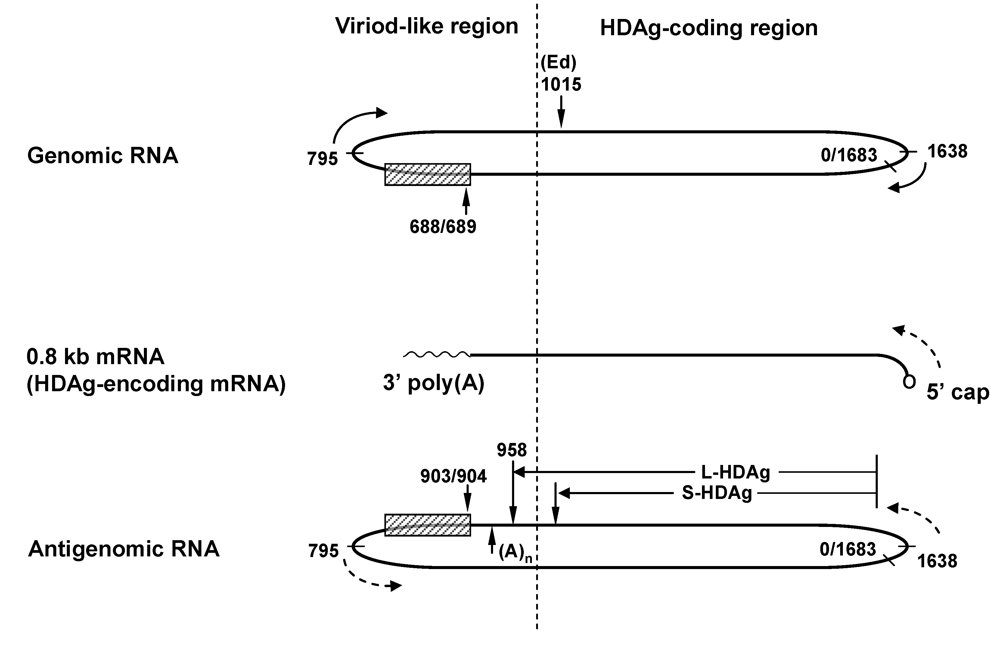
2. Structure of HDV and HDV RNA
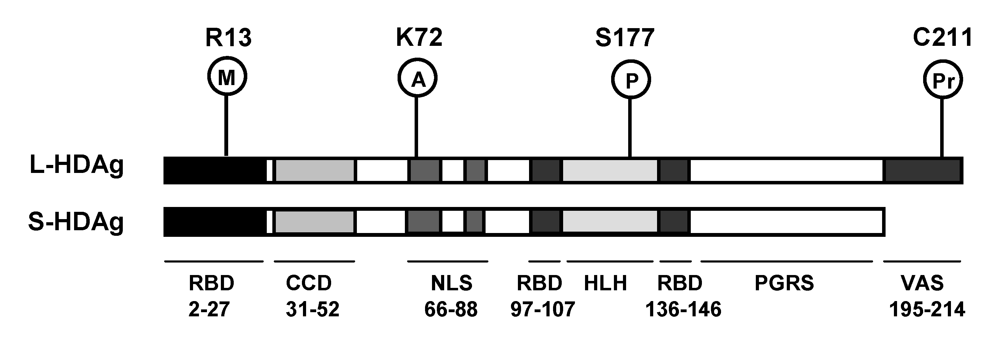
3. HDAg


4. An Overview of the HDV Replication Cycle
5. The Role of HDAg in HDV RNA Replication
6. The Functions of HDAg
6.1. HDAg as a carrier for nuclear import of HDV RNA
6.2. HDAg as a transcription regulator for Pol II
6.3. HDAg as an RNA chaperone
6.4. L-HDAg and RNA replication
7. Enzymology of HDV RNA Replication

{kind=link}
{kind=link}
| Characteristic | mRNA transcription (G to m) | Genomic RNA synthesis (AG to G) | Antigenomic RNA synthesis (G to AG) | References |
|---|---|---|---|---|
| S-HDAg requirement | ||||
| R-13 methylation | Yes | Yes | No | 41,66 |
| K-72 acetylation | Yes | Yes | No | 55,66 |
| S-177 phosphorylation | Yes | Yes | No | 54,66 |
| Assisted by recombinant S-HDAg from E. col | ? | No | Yes | 63 |
| Cytoplasmic transport after synthesis | Yes | Yes | No | 44 |
| Inhibition by L-HDAg | ? | Yes | No | 45 |
| Sensitive to low concentration of α-aminitin (1–5 μg/mL) | Yes | Yes | No | 40,44,47,52 |
| Site of synthesis | nucleoplasm | nucleoplasm | Nucleolus | 24,40 |
| Polymerases | Pol II | Pol II | Pol I (?) | 13,14,19,20,23, 39,40,43,52,53 |
8. Perspectives
References
- Rizzetto, M.; Canese, M.G.; Arico, S.; Crivelli, O.; Trepo, C.; Bonino, F.; Verme, G. Immunofluorescence detection of new antigen-antibody system (delta/anti-delta) associated to hepatitis B virus in liver and in serum of HBsAg carriers. Gut 1977, 18, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Rizzetto, M.; Hoyer, B.; Canese, M.G.; Shih, J.W.; Purcell, R.H.; Gerin, J.L. delta Agent: Association of delta antigen with hepatitis B surface antigen and RNA in serum of delta-infected chimpanzees. Proc. Natl. Acad. Sci. USA 1980, 77, 6124–6128. [Google Scholar] [CrossRef]
- Bonino, F.; Hoyer, B.; Shih, J.W.; Rizzetto, M.; Purcell, R.H.; Gerin, J.L. Delta hepatitis agent: Structural and antigenic properties of the delta-associated particle. Infect. Immun. 1984, 43, 1000–1005. [Google Scholar] [PubMed]
- Sureau, C.; Guerra, B.; Lanford, R.E. Role of the large hepatitis B virus envelope protein in infectivity of the hepatitis delta virion. J. Virol. 1993, 67, 366–372. [Google Scholar] [PubMed]
- Govindarajan, S.; Chin, K.P.; Redeker, A.G.; Peters, R.L. Fulminant B viral hepatitis: Role of delta agent. Gastroenterology 1984, 86, 1417–1420. [Google Scholar] [PubMed]
- Hoofnagle, J.H. Type D (delta) hepatitis. Jama 1989, 261, 1321–1325. [Google Scholar] [PubMed]
- He, L.F.; Ford, E.; Purcell, R.H.; London, W.T.; Phillips, J.; Gerin, J.L. The size of the hepatitis delta agent. J. Med. Virol. 1989, 27, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Ryu, W.S.; Netter, H.J.; Bayer, M.; Taylor, J. Ribonucleoprotein complexes of hepatitis delta virus. J. Virol. 1993, 67, 3281–3287. [Google Scholar] [PubMed]
- Kos, A.; Dijkema, R.; Arnberg, A.C.; van der Meide, P.H.; Schellekens, H. The hepatitis delta (delta) virus possesses a circular RNA. Nature 1986, 323, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.S.; Choo, Q.L.; Weiner, A.J.; Ou, J.H.; Najarian, R.C.; Thayer, R.M.; Mullenbach, G.T.; Denniston, K.J.; Gerin, J.L.; Houghton, M. Structure, sequence and expression of the hepatitis delta (delta) viral genome. Nature 1986, 323, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.Y.; Sharmeen, L.; Dinter-Gottlieb, G.; Taylor, J. Characterization of self-cleaving RNA sequences on the genome and antigenome of human hepatitis delta virus. J. Virol. 1988, 62, 4439–4444. [Google Scholar] [PubMed]
- Macnaughton, T.B.; Wang, Y.J.; Lai, M.M. Replication of hepatitis delta virus RNA: Effect of mutations of the autocatalytic cleavage sites. J. Virol. 1993, 67, 2228–2234. [Google Scholar] [PubMed]
- Jayan, G.C.; Casey, J.L. Inhibition of hepatitis delta virus RNA editing by short inhibitory RNA-mediated knockdown of ADAR1 but not ADAR2 expression. J. Virol. 2002, 76, 12399–12404. [Google Scholar] [CrossRef] [PubMed]
- Jayan, G.C.; Casey, J.L. Increased RNA editing and inhibition of hepatitis delta virus replication by high-level expression of ADAR1 and ADAR2. J. Virol. 2002, 76, 3819–3827. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Lazinski, D.W. Replicating hepatitis delta virus RNA is edited in the nucleus by the small form of ADAR1. Proc. Natl. Acad. Sci. USA 2002, 99, 15118–15123. [Google Scholar] [CrossRef]
- Kuo, M.Y.; Chao, M.; Taylor, J. Initiation of replication of the human hepatitis delta virus genome from cloned DNA: Role of delta antigen. J. Virol. 1989, 63, 1945–1950. [Google Scholar] [PubMed]
- Chang, F.L.; Chen, P.J.; Tu, S.J.; Wang, C.J.; Chen, D.S. The large form of hepatitis delta antigen is crucial for assembly of hepatitis delta virus. Proc. Natl. Acad. Sci. USA 1991, 88, 8490–8494. [Google Scholar] [CrossRef]
- Ryu, W.S.; Bayer, M.; Taylor, J. Assembly of hepatitis delta virus particles. J. Virol. 1992, 66, 2310–2315. [Google Scholar] [PubMed]
- Li, Y.J.; Stallcup, M.R.; Lai, M.M. Hepatitis delta virus antigen is methylated at arginine residues, and methylation regulates subcellular localization and RNA replication. J. Virol. 2004, 78, 13325–13334. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.J.; Chen, D.S.; Chen, P.J. The conserved serine 177 in the delta antigen of hepatitis delta virus is one putative phosphorylation site and is required for efficient viral RNA replication. J. Virol. 2001, 75, 9087–9095. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.J.; Tsay, Y.G.; Juan, L.J.; Fu, T.F.; Huang, W.H.; Chen, D.S.; Chen, P.J. The small delta antigen of hepatitis delta virus is an acetylated protein and acetylation of lysine 72 may influence its cellular localization and viral RNA synthesis. Virology 2004, 319, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Jeng, K.S.; Lai, M.M. Transcription of subgenomic mRNA of hepatitis delta virus requires a modified hepatitis delta antigen that is distinct from antigenomic RNA synthesis. J. Virol. 2008, 82, 9409–9416. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.S.; Watson, J.A.; Havel, C.M.; White, J.M. Identification of a prenylation site in delta virus large antigen. Science 1992, 256, 1331–1333. [Google Scholar] [PubMed]
- Makino, S.; Chang, M.F.; Shieh, C.K.; Kamahora, T.; Vannier, D.M.; Govindarajan, S.; Lai, M.M. Molecular cloning and sequencing of a human hepatitis delta (delta) virus RNA. Nature 1987, 329, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Sheu, G.T.; Lai, M.M. Recombinant hepatitis delta antigen from E. coli promotes hepatitis delta virus RNA replication only from the genomic strand but not the antigenomic strand. Virology 2000, 278, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Tavanez, J.P.; Cunha, C.; Silva, M.C.; David, E.; Monjardino, J.; Carmo-Fonseca, M. Hepatitis delta virus ribonucleoproteins shuttle between the nucleus and the cytoplasm. RNA 2002, 8, 637–646. [Google Scholar] [CrossRef]
- Chou, H.C.; Hsieh, T.Y.; Sheu, G.T.; Lai, M.M. Hepatitis delta antigen mediates the nuclear import of hepatitis delta virus RNA. J. Virol. 1998, 72, 3684–3690. [Google Scholar] [PubMed]
- Chen, P.J.; Kalpana, G.; Goldberg, J.; Mason, W.; Werner, B.; Gerin, J.; Taylor, J. Structure and replication of the genome of the hepatitis delta virus. Proc. Natl. Acad. Sci. USA 1986, 83, 8774–8778. [Google Scholar] [CrossRef]
- Macnaughton, T.B.; Shi, S.T.; Modahl, L.E.; Lai, M.M. Rolling circle replication of hepatitis delta virus RNA is carried out by two different cellular RNA polymerases. J. Virol. 2002, 76, 3920–3927. [Google Scholar] [CrossRef] [PubMed]
- Branch, A.D.; Robertson, H.D. A replication cycle for viroids and other small infectious RNA's. Science 1984, 223, 450–455. [Google Scholar] [PubMed]
- Reid, C.E.; Lazinski, D.W. A host-specific function is required for ligation of a wide variety of ribozyme-processed RNAs. Proc. Natl. Acad. Sci. USA 2000, 97, 424–429. [Google Scholar] [CrossRef]
- Hsieh, S.Y.; Taylor, J. Regulation of polyadenylation of hepatitis delta virus antigenomic RNA. J. Virol. 1991, 65, 6438–6446. [Google Scholar] [PubMed]
- Modahl, L.E.; Lai, M.M. Transcription of hepatitis delta antigen mRNA continues throughout hepatitis delta virus (HDV) replication: A new model of HDV RNA transcription and replication. J. Virol. 1998, 72, 5449–5456. [Google Scholar] [PubMed]
- Casey, J.L.; Gerin, J.L. Hepatitis D virus RNA editing: Specific modification of adenosine in the antigenomic RNA. J. Virol. 1995, 69, 7593–7600. [Google Scholar] [PubMed]
- Luo, G.X.; Chao, M.; Hsieh, S.Y.; Sureau, C.; Nishikura, K.; Taylor, J. A specific base transition occurs on replicating hepatitis delta virus RNA. J. Virol. 1990, 64, 1021–1027. [Google Scholar] [PubMed]
- Lazinski, D.W.; Taylor, J.M. Relating structure to function in the hepatitis delta virus antigen. J. Virol. 1993, 67, 2672–2680. [Google Scholar] [PubMed]
- Lee, C.Z.; Lin, J.H.; Chao, M.; McKnight, K.; Lai, M.M. RNA-binding activity of hepatitis delta antigen involves two arginine-rich motifs and is required for hepatitis delta virus RNA replication. J. Virol. 1993, 67, 2221–2227. [Google Scholar] [PubMed]
- Xia, Y.P.; Yeh, C.T.; Ou, J.H.; Lai, M.M. Characterization of nuclear targeting signal of hepatitis delta antigen: Nuclear transport as a protein complex. J. Virol. 1992, 66, 914–921. [Google Scholar] [PubMed]
- Beard, M.R.; MacNaughton, T.B.; Gowans, E.J. Identification and characterization of a hepatitis delta virus RNA transcriptional promoter. J. Virol. 1996, 70, 4986–4995. [Google Scholar] [PubMed]
- Filipovska, J.; Konarska, M.M. Specific HDV RNA-templated transcription by pol II in vitro. RNA 2000, 6, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Gudima, S.; Wu, S.Y.; Chiang, C.M.; Moraleda, G.; Taylor, J. Origin of hepatitis delta virus mRNA. J. Virol. 2000, 74, 7204–7210. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Filipovska, J.; Yano, K.; Furuya, A.; Inukai, N.; Narita, T.; Wada, T.; Sugimoto, S.; Konarska, M.M.; Handa, H. Stimulation of RNA polymerase II elongation by hepatitis delta antigen. Science 2001, 293, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Mai, R.T.; Lee, Y.H. Transcription factor YY1 and its associated acetyltransferases CBP and p300 interact with hepatitis delta antigens and modulate hepatitis delta virus RNA replication. J. Virol. 2008, 82, 7313–7324. [Google Scholar] [CrossRef] [PubMed]
- Nedialkov, Y.A.; Gong, X.Q.; Hovde, S.L.; Yamaguchi, Y.; Handa, H.; Geiger, J.H.; Yan, H.; Burton, Z.F. NTP-driven translocation by human RNA polymerase II. J. Biol. Chem. 2003, 278, 18303–18312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yan, H.; Burton, Z.F. Combinatorial control of human RNA polymerase II (RNAP II) pausing and transcript cleavage by transcription factor IIF, hepatitis delta antigen, and stimulatory factor II. J. Biol. Chem. 2003, 278, 50101–50111. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Mura, T.; Chanarat, S.; Okamoto, S.; Handa, H. Hepatitis delta antigen binds to the clamp of RNA polymerase II and affects transcriptional fidelity. Genes Cells 2007, 12, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Jeng, K.S.; Su, P.Y.; Lai, M.M. Hepatitis delta antigens enhance the ribozyme activities of hepatitis delta virus RNA in vivo. J. Virol. 1996, 70, 4205–4209. [Google Scholar] [PubMed]
- Huang, Z.S.; Su, W.H.; Wang, J.L.; Wu, H.N. Selective strand annealing and selective strand exchange promoted by the N-terminal domain of hepatitis delta antigen. J. Biol. Chem. 2003, 278, 5685–5693. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.S.; Wu, H.N. Identification and characterization of the RNA chaperone activity of hepatitis delta antigen peptides. J. Biol. Chem. 1998, 273, 26455–26461. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Chang, T.C.; Lin, C.W.; Tsui, H.L.; Chu, P.B.; Chen, B.S.; Huang, Z.S.; Wu, H.N. Nucleic acid binding properties of the nucleic acid chaperone domain of hepatitis delta antigen. Nucleic Acids Res. 2003, 31, 6481–6492. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.; Hsieh, S.Y.; Taylor, J. Role of two forms of hepatitis delta virus antigen: Evidence for a mechanism of self-limiting genome replication. J. Virol. 1990, 64, 5066–5069. [Google Scholar] [PubMed]
- Glenn, J.S.; White, J.M. trans-dominant inhibition of human hepatitis delta virus genome replication. J. Virol. 1991, 65, 2357–2361. [Google Scholar] [PubMed]
- Sato, S.; Cornillez-Ty, C.; Lazinski, D.W. By inhibiting replication, the large hepatitis delta antigen can indirectly regulate amber/W editing and its own expression. J. Virol. 2004, 78, 8120–8134. [Google Scholar] [CrossRef] [PubMed]
- Macnaughton, T.B.; Li, Y.I.; Doughty, A.L.; Lai, M.M. Hepatitis delta virus RNA encoding the large delta antigen cannot sustain replication due to rapid accumulation of mutations associated with RNA editing. J. Virol. 2003, 77, 12048–12056. [Google Scholar] [CrossRef] [PubMed]
- Macnaughton, T.B.; Lai, M.M. Large hepatitis delta antigen is not a suppressor of hepatitis delta virus RNA synthesis once RNA replication is established. J. Virol. 2002, 76, 9910–9919. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Monjardino, J.; Cheng, D.; Krause, S.; Carmo-Fonseca, M. Localization of hepatitis delta virus RNA in the nucleus of human cells. RNA 1998, 4, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Bell, P.; Brazas, R.; Ganem, D.; Maul, G.G. Hepatitis delta virus replication generates complexes of large hepatitis delta antigen and antigenomic RNA that affiliate with and alter nuclear domain 10. J. Virol. 2000, 74, 5329–5336. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Macnaughton, T.; Gao, L.; Lai, M.M. RNA-templated replication of hepatitis delta virus: Genomic and antigenomic RNAs associate with different nuclear bodies. J. Virol. 2006, 80, 6478–6486. [Google Scholar] [CrossRef] [PubMed]
- Dezelee, S.; Sentenac, A.; Fromageot, P. Role of deoxyribonucleic acid-ribonucleic acid hybrids in eukaryotes. Synthetic ribo- and deoxyribopolynucleotides as template for yeast ribonucleic acid polymerase B (or II). J. Biol. Chem. 1974, 249, 5978–5983. [Google Scholar] [PubMed]
- Johnson, T.L.; Chamberlin, M.J. Complexes of yeast RNA polymerase II and RNA are substrates for TFIIS-induced RNA cleavage. Cell 1994, 77, 217–224. [Google Scholar] [CrossRef] [PubMed]
- MacNaughton, T.B.; Gowans, E.J.; McNamara, S.P.; Burrell, C.J. Hepatitis delta antigen is necessary for access of hepatitis delta virus RNA to the cell transcriptional machinery but is not part of the transcriptional complex. Virology 1991, 184, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.B.; Taylor, J. The RNAs of hepatitis delta virus are copied by RNA polymerase II in nuclear homogenates. J. Virol. 1993, 67, 6965–6972. [Google Scholar] [PubMed]
- Modahl, L.E.; Macnaughton, T.B.; Zhu, N.; Johnson, D.L.; Lai, M.M. RNA-Dependent replication and transcription of hepatitis delta virus RNA involve distinct cellular RNA polymerases. Mol.Cell. Biol. 2000, 20, 6030–6039. [Google Scholar] [CrossRef] [PubMed]
- Moraleda, G.; Taylor, J. Host RNA polymerase requirements for transcription of the human hepatitis delta virus genome. J. Virol. 2001, 75, 10161–10169. [Google Scholar] [CrossRef] [PubMed]
- Abrahem, A.; Pelchat, M. Formation of an RNA polymerase II preinitiation complex on an RNA promoter derived from the hepatitis delta virus RNA genome. Nucleic Acids Res. 2008, 36, 5201–5211. [Google Scholar] [CrossRef] [PubMed]
- Greco-Stewart, V.S.; Miron, P.; Abrahem, A.; Pelchat, M. The human RNA polymerase II interacts with the terminal stem-loop regions of the hepatitis delta virus RNA genome. Virology 2007, 357, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, E.; Brueckner, F.; Cramer, P. Molecular basis of RNA-dependent RNA polymerase II activity. Nature 2007, 450, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Macnaughton, T.B.; Lai, M.M. Genomic but not antigenomic hepatitis delta virus RNA is preferentially exported from the nucleus immediately after synthesis and processing. J. Virol. 2002, 76, 3928–3935. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Yung, B.Y.; Syu, W.J.; Lee, Y.H. The nucleolar phosphoprotein B23 interacts with hepatitis delta antigens and modulates the hepatitis delta virus RNA replication. J. Biol. Chem. 2001, 276, 25166–25175. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Chang, S.C.; Chen, C.J.; Chang, M.F. The nucleolin binding activity of hepatitis delta antigen is associated with nucleolus targeting. J. Biol. Chem. 1998, 273, 7650–7656. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Chen, Y.S.; Chen, P.J. Nucleolar targeting of hepatitis delta antigen abolishes its ability to initiate viral antigenomic RNA replication. J. Virol. 2008, 82, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Greco-Stewart, V.S.; Schissel, E.; Pelchat, M. The hepatitis delta virus RNA genome interacts with the human RNA polymerases I and III. Virology 2009, 386, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Maida, Y.; Yasukawa, M.; Furuuchi, M.; Lassmann, T.; Possemato, R.; Okamoto, N.; Kasim, V.; Hayashizaki, Y.; Hahn, W.C.; Masutomi, K. An RNA-dependent RNA polymerase formed by TERT and the RMRP RNA. Nature 2009, 461, 230–235. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Tseng, C.-H.; Lai, M.M.C. Hepatitis Delta Virus RNA Replication. Viruses 2009, 1, 818-831. https://doi.org/10.3390/v1030818
Tseng C-H, Lai MMC. Hepatitis Delta Virus RNA Replication. Viruses. 2009; 1(3):818-831. https://doi.org/10.3390/v1030818
Chicago/Turabian StyleTseng, Chung-Hsin, and Michael M. C. Lai. 2009. "Hepatitis Delta Virus RNA Replication" Viruses 1, no. 3: 818-831. https://doi.org/10.3390/v1030818