The Abrogation of Phosphorylation Plays a Relevant Role in the CCR5 Signalosome Formation with Natural Antibodies to CCR5

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Reagents
2.3. Study Population
2.4. Ethics Statement
2.5. CCR5 Internalization Assay
2.6. Western Blotting
2.7. Immunoprecipitation
2.8. siRNA Nucleofection
2.9. Statistical Analysis
3. Results
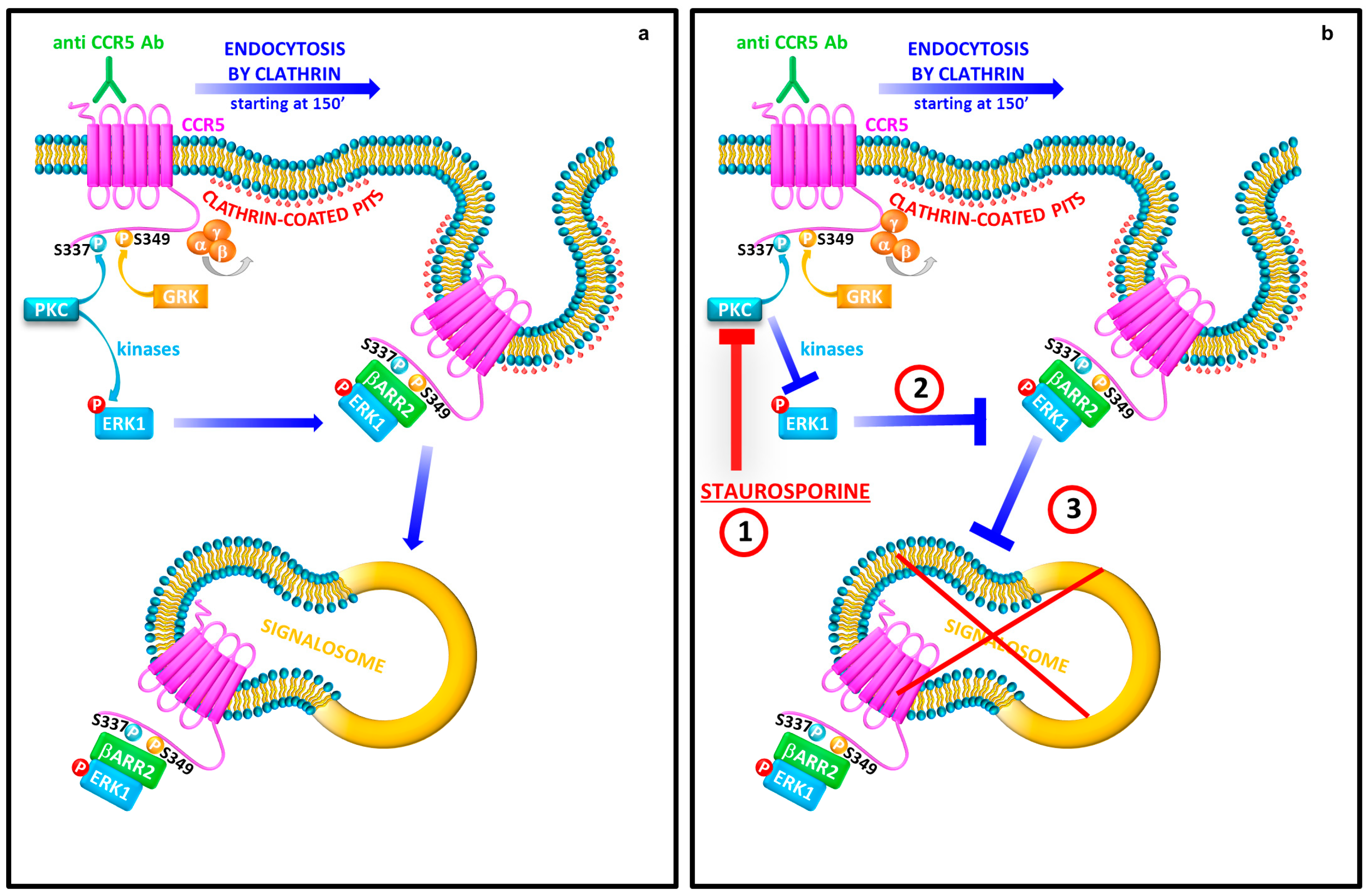
3.1. Staurosporine Treatment Reduces the Cytoplasmic Accumulation of CCR5
3.2. Protein Kinases Activity Plays a Key Role for the CCR5 Signalosome Formation and Stabilization
3.3. Staurosporine Treatment Alters the Regulation of the Core Components of CCR5 Pathway
3.4. β-arrestin2 Is Crucial for the CCR5 Signalosome Formation
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hall, R.A.; Lefkowitz, R.J. Regulation of G protein-coupled receptor signaling by scaffold proteins. Circ. Res. 2002, 91, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J.; Shenoy, S.K. Transduction of receptor signals by β-arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L.M.; Lefkowitz, R.J. The role of β-arrestins in the termination and transduction of G-protein-coupled receptor signals. J. Cell Sci. 2002, 115, 455–465. [Google Scholar] [PubMed]
- Jean-Charles, P.-Y.; Rajiv, V.; Shenoy, S.K. Ubiquitin-Related Roles of β-Arrestins in Endocytic Trafficking and Signal Transduction. J. Cell. Physiol. 2016, 231, 2071–2080. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Lefkowitz, R.J. GRKs and β-arrestins: Roles in receptor silencing, trafficking and signaling. Trends Endocrinol. Metab. 2006, 17, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.K.; Drake, M.T.; Nelson, C.D.; Houtz, D.A.; Xiao, K.; Madabushi, S.; Reiter, E.; Premont, R.T.; Lichtarge, O.; Lefkowitz, R.J. β-arrestin-dependent, G protein-independent ERK1/2 activation by the β2 adrenergic receptor. J. Biol. Chem. 2006, 281, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.; Kelly, E.; Strange, P.G. Pathways for internalization and recycling of the chemokine receptor CCR5. Blood 2002, 99, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Lederman, M.M.; Penn-Nicholson, A.; Cho, M.; Mosier, D. Biology of CCR5 and its role in HIV infection and treatment. JAMA 2006, 296, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Pastori, C.; Weiser, B.; Barassi, C.; Uberti-Foppa, C.; Ghezzi, S.; Longhi, R.; Calori, G.; Burger, H.; Kemal, K.; Poli, G.; et al. Long-lasting CCR5 internalization by antibodies in a subset of long-term nonprogressors: A possible protective effect against disease progression. Blood 2006, 107, 4825–4833. [Google Scholar] [CrossRef] [PubMed]
- Venuti, A.; Pastori, C.; Pennisi, R.; Riva, A.; Sciortino, M.T.; Lopalco, L. Class B β-arrestin2-dependent CCR5 signalosome retention with natural antibodies to CCR5. Sci. Rep. 2016, 6, 39382. [Google Scholar] [CrossRef] [PubMed]
- Venuti, A.; Pastori, C.; Siracusano, G.; Riva, A.; Sciortino, M.T.; Lopalco, L. ERK1-Based Pathway as a New Selective Mechanism To Modulate CCR5 with Natural Antibodies. J. Immunol. 2015, 195, 3045–3057. [Google Scholar] [CrossRef] [PubMed]
- Pollok-Kopp, B.; Schwarze, K.; Baradari, V.K.; Oppermann, M. Analysis of ligand-stimulated CC chemokine receptor 5 (CCR5) phosphorylation in intact cells using phosphosite-specific antibodies. J. Biol. Chem. 2003, 278, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Bomsel, M.; Pastori, C.; Tudor, D.; Alberti, C.; Garcia, S.; Ferrari, D.; Lazzarin, A.; Lopalco, L. Natural mucosal antibodies reactive with first extracellular loop of CCR5 inhibit HIV-1 transport across human epithelial cells. AIDS Lond. Engl. 2007, 21, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, M.T.; Suzuki, M.; Taddeo, B.; Roizman, B. RNAs extracted from herpes simplex virus 1 virions: Apparent selectivity of viral but not cellular RNAs packaged in virions. J. Virol. 2001, 75, 8105–8116. [Google Scholar] [CrossRef] [PubMed]
- Eishingdrelo, H.; Sun, W.; Li, H.; Wang, L.; Eishingdrelo, A.; Dai, S.; McKew, J.C.; Zheng, W. ERK and β-arrestin interaction: A converging point of signaling pathways for multiple types of cell surface receptors. J. Biomol. Screen. 2015, 20, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Signoret, N.; Pelchen-Matthews, A.; Mack, M.; Proudfoot, A.E.; Marsh, M. Endocytosis and recycling of the HIV coreceptor CCR5. J. Cell Biol. 2000, 151, 1281–1294. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, M. Chemokine receptor CCR5: Insights into structure, function, and regulation. Cell. Signal. 2004, 16, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, S.; Rose, J.J.; Lodge, R.; Murphy, P.M.; Foley, J.F. Distinct mechanisms of agonist-induced endocytosis for human chemokine receptors CCR5 and CXCR4. Mol. Biol. Cell 2003, 14, 3305–3324. [Google Scholar] [CrossRef] [PubMed]
- Signoret, N.; Hewlett, L.; Wavre, S.; Pelchen-Matthews, A.; Oppermann, M.; Marsh, M. Agonist-induced endocytosis of CC chemokine receptor 5 is clathrin dependent. Mol. Biol. Cell 2005, 16, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.K. Deubiquitinases and their emerging roles in β-arrestin-mediated signaling. Methods Enzymol. 2014, 535, 351–370. [Google Scholar] [CrossRef] [PubMed]
- DeFea, K.A. β-arrestins as regulators of signal termination and transduction: How do they determine what to scaffold? Cell. Signal. 2011, 23, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; DeFea, K.A. β-Arrestin-kinase scaffolds: Turn them on or turn them off? Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 5, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.K.; Xiao, K.; Lefkowitz, R.J. Emerging paradigms of β-arrestin-dependent seven transmembrane receptor signaling. Trends Biochem. Sci. 2011, 36, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Ahn, S.; Shukla, A.K.; Lefkowitz, R.J. Molecular mechanism of β-arrestin-biased agonism at seven-transmembrane receptors. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Eichel, K.; Jullié, D.; von Zastrow, M. β-Arrestin drives MAP kinase signalling from clathrin-coated structures after GPCR dissociation. Nat. Cell Biol. 2016, 18, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes. Cell 2006, 124, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Aivazian, D. Targeting Rab GTPases to distinct membrane compartments. Nat. Rev. Mol. Cell Biol. 2004, 5, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Chavrier, P.; Parton, R.G.; Hauri, H.P.; Simons, K.; Zerial, M. Localization of low molecular weight GTP binding proteins to exocytic and endocytic compartments. Cell 1990, 62, 317–329. [Google Scholar] [CrossRef]
- Ullrich, O.; Reinsch, S.; Urbé, S.; Zerial, M.; Parton, R.G. Rab11 regulates recycling through the pericentriolar recycling endosome. J. Cell Biol. 1996, 135, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Barbero, P.; Bittova, L.; Pfeffer, S.R. Visualization of Rab9-mediated vesicle transport from endosomes to the trans-Golgi in living cells. J. Cell Biol. 2002, 156, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Lopalco, L.; Barassi, C.; Pastori, C.; Longhi, R.; Burastero, S.E.; Tambussi, G.; Mazzotta, F.; Lazzarin, A.; Clerici, M.; Siccardi, A.G. CCR5-reactive antibodies in seronegative partners of HIV-seropositive individuals down-modulate surface CCR5 in vivo and neutralize the infectivity of R5 strains of HIV-1 in vitro. J. Immunol. 2000, 164, 3426–3433. [Google Scholar] [CrossRef] [PubMed]
- Barassi, C.; Lazzarin, A.; Lopalco, L. CCR5-specific mucosal IgA in saliva and genital fluids of HIV-exposed seronegative subjects. Blood 2004, 104, 2205–2206. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Vanderwinden, J.-M.; Cihak, J.; Wittamer, V.; Le Poul, E.; Issafras, H.; Stangassinger, M.; Vassart, G.; Marullo, S.; Schlndorff, D.; et al. Multiple active states and oligomerization of CCR5 revealed by functional properties of monoclonal antibodies. Mol. Biol. Cell 2002, 13, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Bouhlal, H.; Hocini, H.; Quillent-Grégoire, C.; Donkova, V.; Rose, S.; Amara, A.; Longhi, R.; Haeffner-Cavaillon, N.; Beretta, A.; Kaveri, S.V.; et al. Antibodies to C-C chemokine receptor 5 in normal human IgG block infection of macrophages and lymphocytes with primary R5-tropic strains of HIV-1. J. Immunol. 2001, 166, 7606–7611. [Google Scholar] [CrossRef] [PubMed]
- Ditzel, H.J.; Rosenkilde, M.M.; Garred, P.; Wang, M.; Koefoed, K.; Pedersen, C.; Burton, D.R.; Schwartz, T.W. The CCR5 receptor acts as an alloantigen in CCR5Δ32 homozygous individuals: Identification of chemokineand HIV-1-blocking human antibodies. Proc. Natl. Acad. Sci. USA 1998, 95, 5241–5245. [Google Scholar] [CrossRef] [PubMed]
- Lopalco, L. Natural anti-CCR5 antibodies in HIV-infection and -exposure. J. Transl. Med. 2011, 9 (Suppl. 1), S4. [Google Scholar] [CrossRef] [PubMed]
- Pastori, C.; Diomede, L.; Venuti, A.; Fisher, G.; Jarvik, J.; Bomsel, M.; Sanvito, F.; Lopalco, L. Induction of HIV-blocking anti-CCR5 IgA in Peyers’s patches without histopathological alterations. J. Virol. 2014, 88, 3623–3635. [Google Scholar] [CrossRef] [PubMed]
- Barassi, C.; Soprana, E.; Pastori, C.; Longhi, R.; Buratti, E.; Lillo, F.; Marenzi, C.; Lazzarin, A.; Siccardi, A.G.; Lopalco, L. Induction of murine mucosal CCR5-reactive antibodies as an anti-human immunodeficiency virus strategy. J. Virol. 2005, 79, 6848–6858. [Google Scholar] [CrossRef] [PubMed]
- Bönsch, C.; Munteanu, M.; Rossitto-Borlat, I.; Fürstenberg, A.; Hartley, O. Potent Anti-HIV Chemokine Analogs Direct Post-Endocytic Sorting of CCR5. PLoS ONE 2015, 10, e0125396. [Google Scholar] [CrossRef] [PubMed]
- Ajuebor, M.N.; Carey, J.A.; Swain, M.G. CCR5 in T cell-mediated liver diseases: What’s going on? J. Immunol. 2006, 177, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Velázquez, M.; Xolalpa, W.; Pestell, R.G. The potential to target CCL5/CCR5 in breast cancer. Expert Opin. Ther. Targets 2014, 18, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-J.; Du, Y.; Zhao, X.; Ma, L.-Y.; Cao, G.-W. Inflammation-related factors predicting prognosis of gastric cancer. World J. Gastroenterol. 2014, 20, 4586–4596. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venuti, A.; Pastori, C.; Siracusano, G.; Pennisi, R.; Riva, A.; Tommasino, M.; Sciortino, M.T.; Lopalco, L. The Abrogation of Phosphorylation Plays a Relevant Role in the CCR5 Signalosome Formation with Natural Antibodies to CCR5. Viruses 2018, 10, 9. https://doi.org/10.3390/v10010009
Venuti A, Pastori C, Siracusano G, Pennisi R, Riva A, Tommasino M, Sciortino MT, Lopalco L. The Abrogation of Phosphorylation Plays a Relevant Role in the CCR5 Signalosome Formation with Natural Antibodies to CCR5. Viruses. 2018; 10(1):9. https://doi.org/10.3390/v10010009
Chicago/Turabian StyleVenuti, Assunta, Claudia Pastori, Gabriel Siracusano, Rosamaria Pennisi, Agostino Riva, Massimo Tommasino, Maria Teresa Sciortino, and Lucia Lopalco. 2018. "The Abrogation of Phosphorylation Plays a Relevant Role in the CCR5 Signalosome Formation with Natural Antibodies to CCR5" Viruses 10, no. 1: 9. https://doi.org/10.3390/v10010009