Ebola Virus Causes Intestinal Tract Architectural Disruption and Bacterial Invasion in Non-Human Primates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Use and Viral Challenge
2.2. In Situ Hybridization
2.3. Immunofluorescence Assay
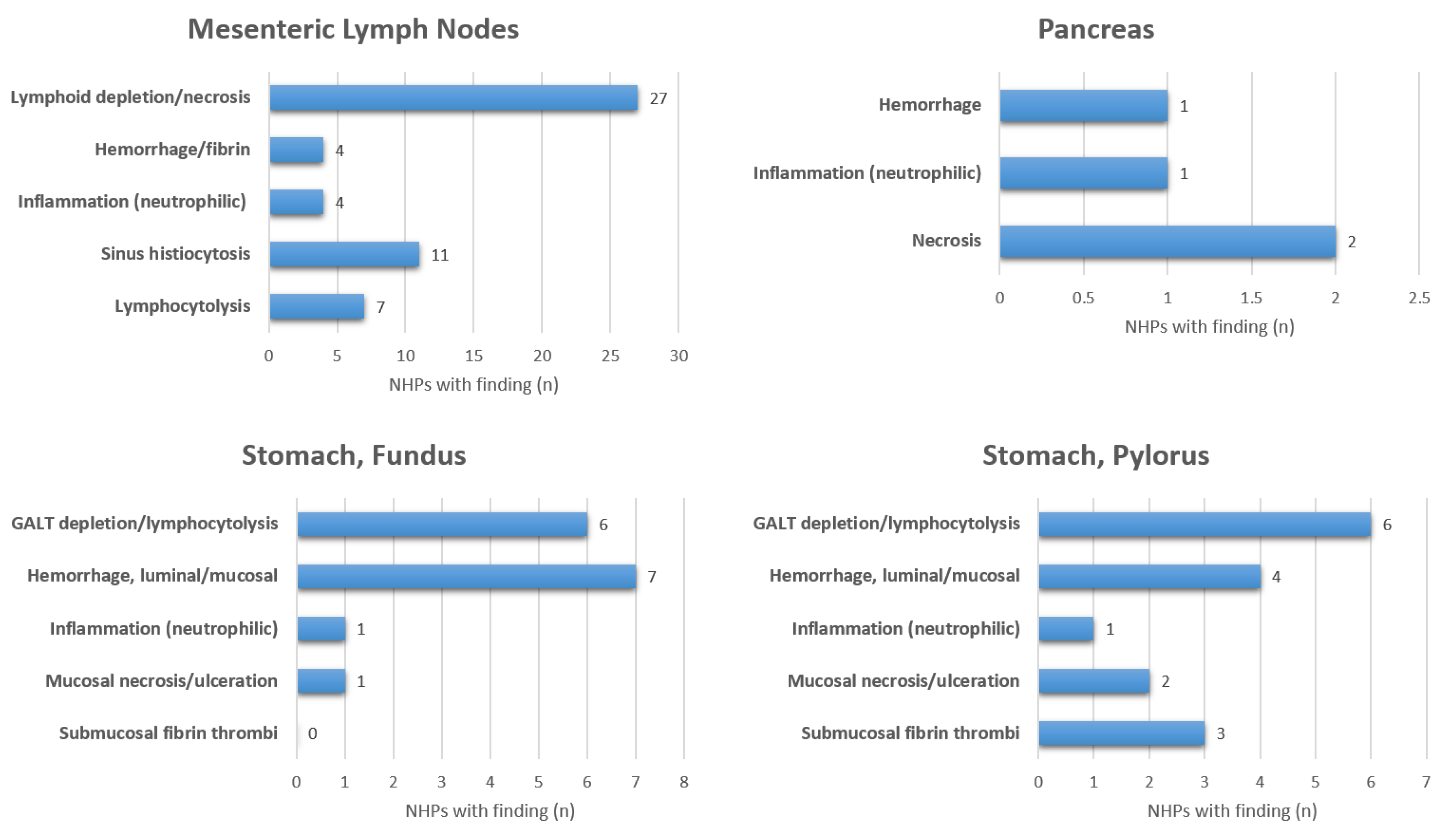
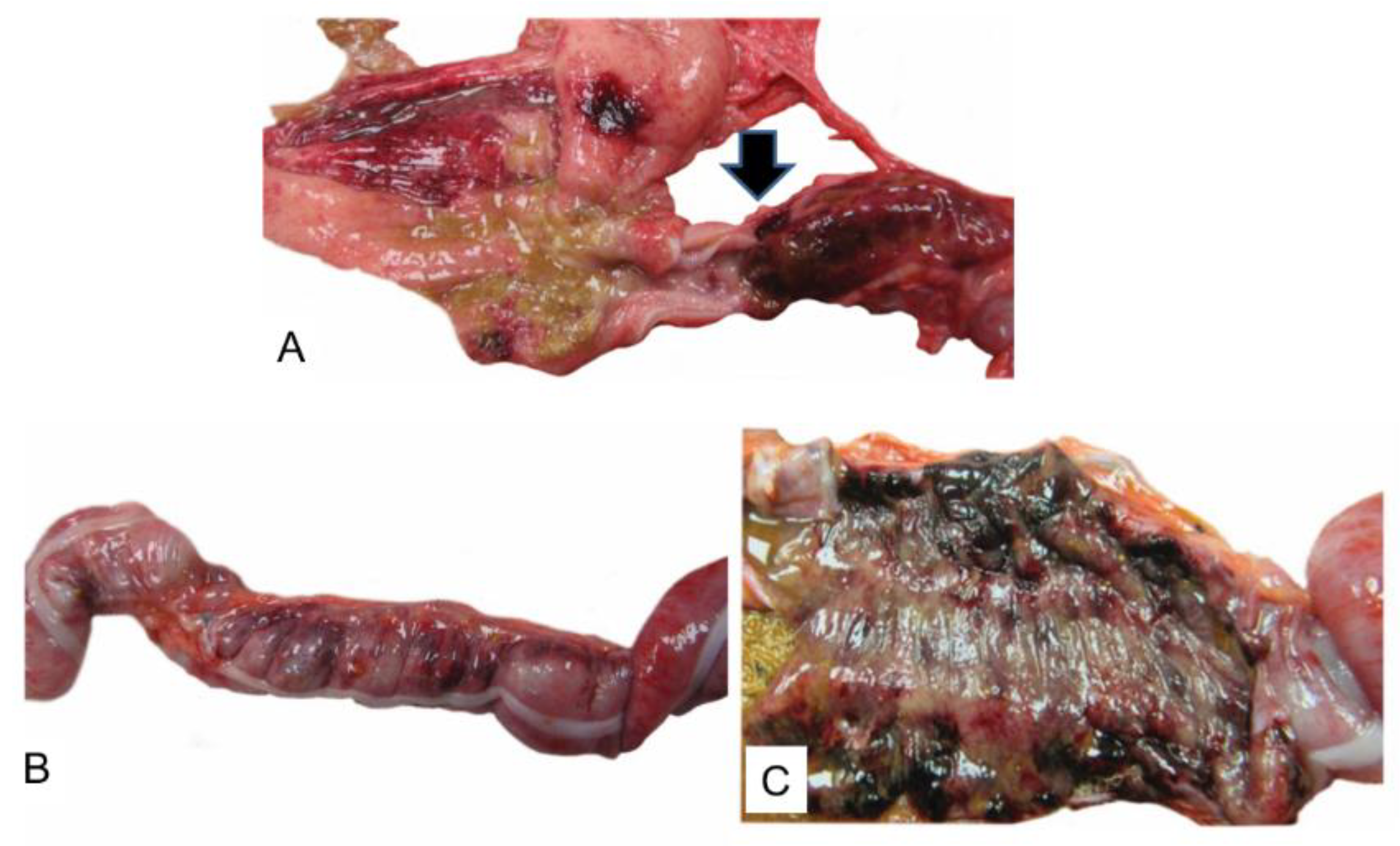
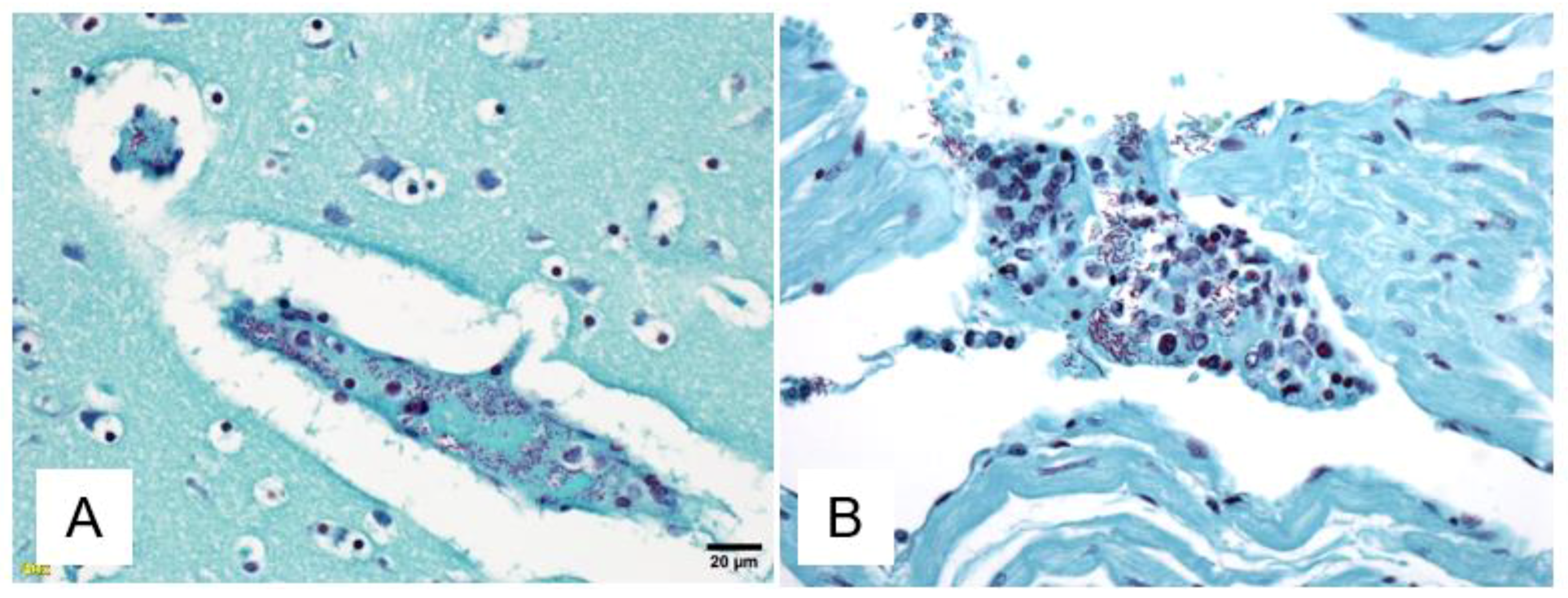
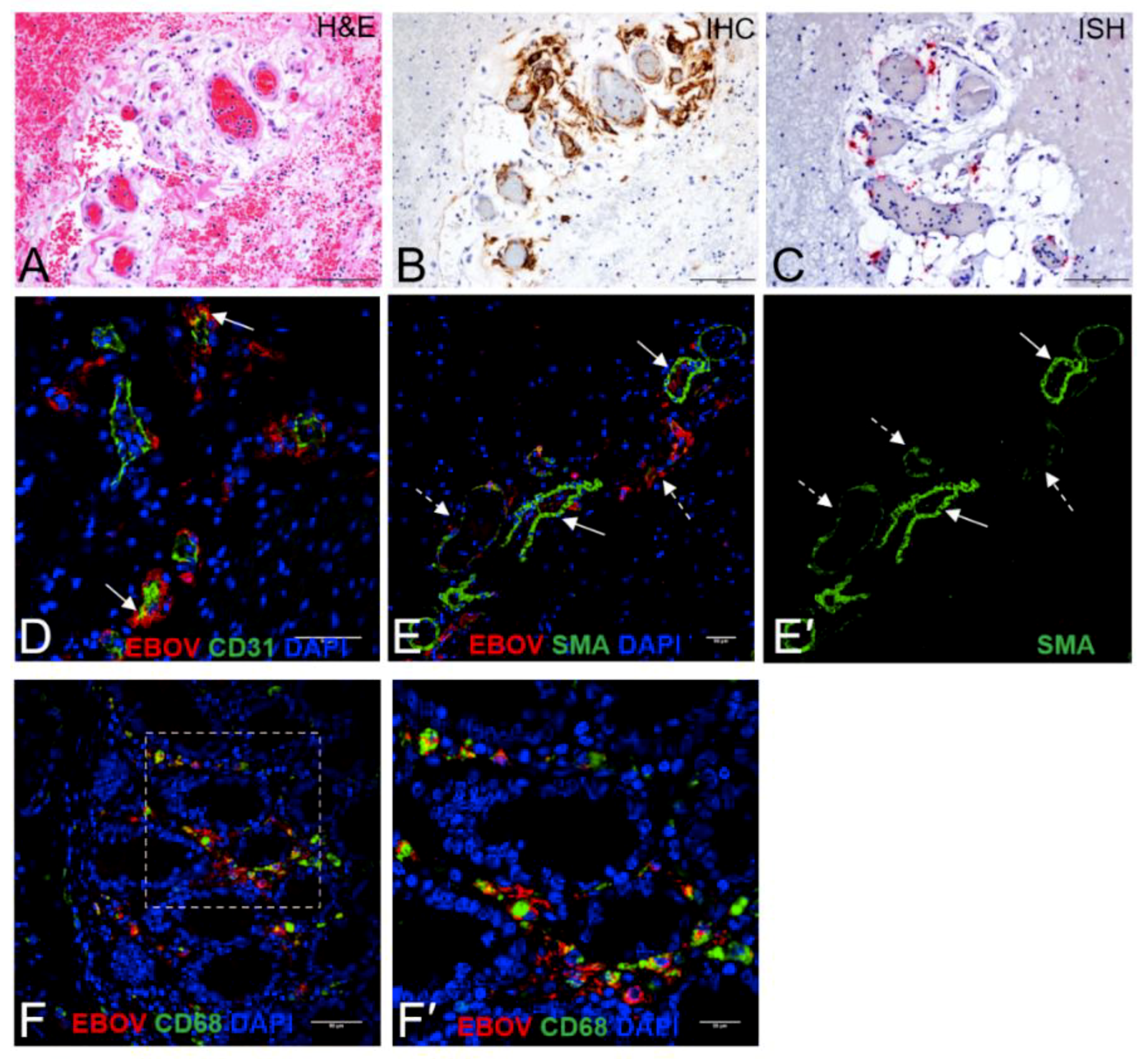
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lado, M.; Walker, N.F.; Baker, P.; Haroon, S.; Brown, C.S.; Youkee, D.; Studd, N.; Kessete, Q.; Maini, R.; Boyles, T.; et al. Clinical features of patients isolated for suspected Ebola virus disease at Connaught Hospital, Freetown, Sierra Leone: A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 1024–1033. [Google Scholar] [CrossRef]
- Chertow, D.S.; Kleine, C.; Edwards, J.K.; Scaini, R.; Giuliani, R.; Sprecher, A. Ebola virus disease in West Africa—Clinical manifestations and management. N. Engl. J. Med. 2014, 371, 2054–2057. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.J.; Duan, X.Z.; Gao, X.D.; Li, L.; Li, C.; Ji, D.; Li, W.G.; Wang, L.F.; Meng, Y.H.; Yang, X.; et al. Clinical presentations and outcomes of patients with Ebola virus disease in Freetown, Sierra Leone. Infect. Dis. Poverty 2016, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Mu, J.; Qin, E.; Wang, Y.; Liu, L.; Wu, D.; Jia, H.; Li, Z.; Guo, T.; Wang, X.; et al. Clinical characteristics of 154 patients suspected of having Ebola virus disease in the Ebola holding center of Jui Government Hospital in Sierra Leone during the 2014 Ebola outbreak. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2089–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuels, B.; Wichmann, D.; Emmerich, P.; Schmidt-Chanasit, J.; de Heer, G.; Kluge, S.; Sow, A.; Renne, T.; Gunther, S.; Lohse, A.W.; et al. A case of severe Ebola virus infection complicated by gram-negative septicemia. N. Engl. J. Med. 2014, 371, 2394–2401. [Google Scholar] [CrossRef] [PubMed]
- Nalin, D.R.; Hirschhorn, N. Ebola and cholera. Am. J. Trop. Med. Hyg. 2015, 92, 1081. [Google Scholar] [CrossRef] [PubMed]
- Dickson, S.J.; Clay, K.A.; Adam, M.; Ardley, C.; Bailey, M.S.; Burns, D.S.; Cox, A.T.; Craig, D.G.; Espina, M.; Ewington, I.; et al. Enhanced case management can be delivered for patients with EVD in Africa: Experience from a UK military Ebola treatment centre in Sierra Leone. J. Infect. 2018, 76, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar-Jahromi, M.; Martins, K.A.O.; Cardile, A.P.; Reisler, R.B.; Christopher, G.W.; Bavari, S. Treatment-focused Ebola trials, supportive care and future of filovirus care. Expert Rev. Anti Infect. Ther. 2018, 16, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne, F.; Fowler, R.A.; Adhikari, N.K.; Murthy, S.; Brett-Major, D.M.; Jacobs, M.; Uyeki, T.M.; Vallenas, C.; Norris, S.L.; Fischer, W.A., 2nd; et al. Evidence-based guidelines for supportive care of patients with Ebola virus disease. Lancet 2018, 391, 700–708. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Mehta, A.K.; Davey, R.T., Jr.; Liddell, A.M.; Wolf, T.; Vetter, P.; Schmiedel, S.; Grunewald, T.; Jacobs, M.; Arribas, J.R.; et al. Clinical Management of Ebola Virus Disease in the United States and Europe. N. Engl. J. Med. 2016, 374, 636–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manrique, M.; Pareja-Tobes, E.; Pareja, E.; Pareja-Tobes, P.; Tobes, R. Bacterial sequences detected in 99 out of 99 serum samples from Ebola patients. bioRxiv 2016. [Google Scholar] [CrossRef]
- Carroll, M.W.; Haldenby, S.; Rickett, N.Y.; Palyi, B.; Garcia-Dorival, I.; Liu, X.; Barker, G.; Bore, J.A.; Koundouno, F.R.; Williamson, E.D.; et al. Deep Sequencing of RNA from Blood and Oral Swab Samples Reveals the Presence of Nucleic Acid from a Number of Pathogens in Patients with Acute Ebola Virus Disease and Is Consistent with Bacterial Translocation across the Gut. mSphere 2017, 2, e00325-17. [Google Scholar] [CrossRef] [PubMed]
- Martines, R.B.; Ng, D.L.; Greer, P.W.; Rollin, P.E.; Zaki, S.R. Tissue and cellular tropism, pathology and pathogenesis of Ebola and Marburg viruses. J. Pathol. 2015, 235, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Hensley, L.E.; Larsen, T.; Young, H.A.; Reed, D.S.; Geisbert, J.B.; Scott, D.P.; Kagan, E.; Jahrling, P.B.; Davis, K.J. Pathogenesis of Ebola hemorrhagic fever in cynomolgus macaques: Evidence that dendritic cells are early and sustained targets of infection. Am. J. Pathol. 2003, 163, 2347–2370. [Google Scholar] [CrossRef]
- Veazey, R.S.; Rosenzweig, M.; Shvetz, D.E.; Pauley, D.R.; DeMaria, M.; Chalifoux, L.V.; Johnson, R.P.; Lackner, A.A. Characterization of gut-associated lymphoid tissue (GALT) of normal rhesus macaques. Clin. Immunol. Immunopathol. 1997, 82, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Martins, K.; Cooper, C.; Warren, T.; Wells, J.; Bell, T.; Raymond, J.; Stuthman, K.; Benko, J.; Garza, N.; van Tongeren, S.; et al. Characterization of clinical and immunological parameters during Ebola virus infection of rhesus macaques. Viral Immunol. 2015, 28, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Kugelman, J.R.; Rossi, C.A.; Wiley, M.R.; Ladner, J.T.; Nagle, E.R.; Pfeffer, B.P.; Garcia, K.; Prieto, K.; Wada, J.; Kuhn, J.H.; et al. Informing the Historical Record ofExperimental Nonhuman Primate Infections with Ebola Virus: Genomic Characterization of USAMRIID Ebola Virus/H.sapiens-tc/COD/1995/Kikwit-9510621 Challenge Stock “R4368” and Its Replacement “R4415”. PLoS ONE 2016, 22, e0150919. [Google Scholar]
- Zeng, X.; Blancett, C.D.; Koistinen, K.A.; Schellhase, C.W.; Bearss, J.J.; Radoshitzky, S.R.; Honnold, S.P.; Chance, T.B.; Warren, T.K.; Froude, J.W.; et al. Identification and pathological characterization of persistent asymptomatic Ebola virus infection in rhesus monkeys. Nat. Microbiol. 2017, 2, 17113. [Google Scholar] [CrossRef] [PubMed]
- Aminde, L.N.; Bonko, N.M.; Takah, N.F.; Awungafac, G.; Teno, D. Intussusception as a Cause of Bowel Obstruction in Adults from a Resource Limited Area, Cameroon. Ethiop. J. Health Sci. 2015, 25, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Mahanty, S.; Bray, M. Pathogenesis of filoviral haemorrhagic fevers. Lancet Infect. Dis. 2004, 4, 487–498. [Google Scholar] [CrossRef]
- Reisler, R.B.; Yu, C.; Donofrio, M.J.; Warren, T.K.; Wells, J.B.; Stuthman, K.S.; Garza, N.L.; Vantongeren, S.A.; Donnelly, G.C.; Kane, C.D.; et al. Clinical Laboratory Values as Early Indicators of Ebola Virus Infection in Nonhuman Primates. Emerg. Infect. Dis. 2017, 23, 1316–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.Y.; Duckers, H.J.; Sullivan, N.J.; Sanchez, A.; Nabel, E.G.; Nabel, G.J. Identification of the Ebola virus glycoprotein as the main viral determinant of vascular cell cytotoxicity and injury. Nat. Med. 2000, 6, 886–889. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.; Geisbert, T.W. Ebola virus: The role of macrophages and dendritic cells in the pathogenesis of Ebola hemorrhagic fever. Int. J. Biochem. Cell Biol. 2005, 37, 1560–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vighi, G.; Marcucci, F.; Sensi, L.; Di Cara, G.; Frati, F. Allergy and the gastrointestinal system. Clin. Exp. Immunol. 2008, 153 (Suppl. 1), 3–6. [Google Scholar] [CrossRef] [Green Version]
- Aftab, A. Ansari, Clinical features and pathobiology of Ebolavirus infection. J. Autoimmun. 2014, 55, 1–9. [Google Scholar] [CrossRef]
- Jaax, N.K.; Davis, K.J.; Geisbert, T.J.; Vogel, P.; Jaax, G.P.; Topper, M.; Jahrling, P.B. Lethal experimental infection of rhesus monkeys with Ebola-Zaire (Mayinga) virus by the oral and conjunctival route of exposure. Arch. Pathol. Lab. Med. 1996, 120, 140–155. [Google Scholar] [PubMed]
- Johnson, E.; Jaax, N.; White, J.; Jahrling, P. Lethal experimental infections of rhesus monkeys by aerosolized Ebola virus. Int. J. Exp. Pathol. 1995, 76, 227–236. [Google Scholar] [PubMed]
- Twenhafel, N.A.; Mattix, M.E.; Johnson, J.C.; Robinson, C.G.; Pratt, W.D.; Cashman, K.A.; Wahl-Jensen, V.; Terry, C.; Olinger, G.G.; Hensley, L.E.; et al. Pathology of experimental aerosol Zaire ebolavirus infection in rhesus macaques. Vet. Pathol. 2013, 50, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Ansumana, R.; Jacobsen, K.H.; Sahr, F.; Idris, M.; Bangura, H.; Boie-Jalloh, M.; Lamin, J.M.; Sesay, S. Ebola in Freetown area, Sierra Leone—A case study of 581 patients. N. Engl. J. Med. 2015, 372, 587–588. [Google Scholar] [CrossRef] [PubMed]
- Damkjaer, M.; Rudolf, F.; Mishra, S.; Young, A.; Storgaard, M. Clinical Features and Outcome of Ebola Virus Disease in Pediatric Patients: A Retrospective Case Series. J. Pediatr. 2017, 182, 378–381 e1. [Google Scholar] [CrossRef] [PubMed]
- Slack, S.; Abbey, I.; Smith, D. Abdominal pain and hyperamylasaemia—Not always pancreatitis. BMJ Case Rep. 2010. [Google Scholar] [CrossRef] [PubMed]
- Van der Worp, H.B.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can Animal Models of Disease Reliably Inform Human Studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [PubMed]
- Webb, D.R. Animal models of human disease: Inflammation. Biochem. Pharmacol. 2014, 87, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Bixler, S.L.; Bocan, T.M.; Wells, J.; Wetzel, K.S.; Van Tongeren, S.A.; Dong, L.; Garza, N.L.; Donnelly, G.; Cazares, L.H.; Nuss, J.; et al. Efficacy of favipiravir (T-705) in nonhuman primates infected with Ebola virus or Marburg virus. Antiviral Res. 2018, 151, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Lee, Y.J.; Kim, J.S.; Chung, J.Y.; Chang, S.; Lee, K.; Choe, B.H.; Hong, S.J.; Song, J.S.; Park, K.Y. Epidemiological Correlation between Fecal Adenovirus Subgroups and Pediatric Intussusception in Korea. J. Korean Med. Sci. 2017, 32, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Yang, S.I.; Kim, J.M.; Kim, J.Y. Clinical features and role of viral isolates from stool samples of intussuception in children. Pediatr. Gastroenterol. Hepatol. Nutr. 2013, 16, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Park, S.; Hong, Y.J.; Lee, S.W.; Cho, M.S. Cytomegalovirus-associated intussusception with florid vascular proliferation in an infant. J. Pathol. Transl. Med. 2015, 49, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Dallatomasina, S.; Crestani, R.; Sylvester Squire, J.; Declerk, H.; Caleo, G.M.; Wolz, A.; Stinson, K.; Patten, G.; Brechard, R.; Gbabai, O.B.; et al. Ebola outbreak in rural West Africa: Epidemiology, clinical features and outcomes. Trop. Med. Int. Health 2015, 20, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Champaloux, S.W.; Keita, S.; Martel, L.; Bilivogui, P.; Knust, B.; McCollum, A.M. Sensitivity and Specificity of Suspected Case Definition Used during West Africa Ebola Epidemic. Emerg. Infect. Dis. 2018, 24, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | (N = 31) |
|---|---|
| Mean weight, kg [range] | 4.9 [3.8–7.3] |
| Mean age, months [range] | 48.3 [35–79] |
| Male:Female ratio | 15:16 |
| Day 5 Post-infection (log 10 copies/mL) | 8.68 |
| Survival time (days) [range] | 8.0 [5–15] |
| Found Dead | 6% |
| Mean Time from Death to Necropsy (minutes) [range] | 409.8 [25–2278] |
| NHPs showing evidence of bacterial invasion of intestinal tract tissue (gram-negative rods) | 29% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reisler, R.B.; Zeng, X.; Schellhase, C.W.; Bearss, J.J.; Warren, T.K.; Trefry, J.C.; Christopher, G.W.; Kortepeter, M.G.; Bavari, S.; Cardile, A.P. Ebola Virus Causes Intestinal Tract Architectural Disruption and Bacterial Invasion in Non-Human Primates. Viruses 2018, 10, 513. https://doi.org/10.3390/v10100513
Reisler RB, Zeng X, Schellhase CW, Bearss JJ, Warren TK, Trefry JC, Christopher GW, Kortepeter MG, Bavari S, Cardile AP. Ebola Virus Causes Intestinal Tract Architectural Disruption and Bacterial Invasion in Non-Human Primates. Viruses. 2018; 10(10):513. https://doi.org/10.3390/v10100513
Chicago/Turabian StyleReisler, Ronald B., Xiankun Zeng, Christopher W. Schellhase, Jeremy J. Bearss, Travis K. Warren, John C. Trefry, George W. Christopher, Mark G. Kortepeter, Sina Bavari, and Anthony P. Cardile. 2018. "Ebola Virus Causes Intestinal Tract Architectural Disruption and Bacterial Invasion in Non-Human Primates" Viruses 10, no. 10: 513. https://doi.org/10.3390/v10100513