Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner

Abstract
:1. Introduction
2. Materials and Methods
2.1. Computer-Assisted Analysis
2.2. Cell Lines, Viruses, and Insect Larvae
2.3. Construction of an ac111-Knockout Virus
2.4. Viral Propagation Analysis
2.5. Real-Time PCR (qPCR) Analysis
2.6. Transmission Electron Microscopy (TEM)
2.7. Bioassays in T. ni and S. exigua Larvae
3. Results
3.1. Sequence Analysis of Ac111 and Its Homologues
3.2. Construction of an ac111-Knockout AcMNPV
3.3. Ac111 is Not Required forBV Production
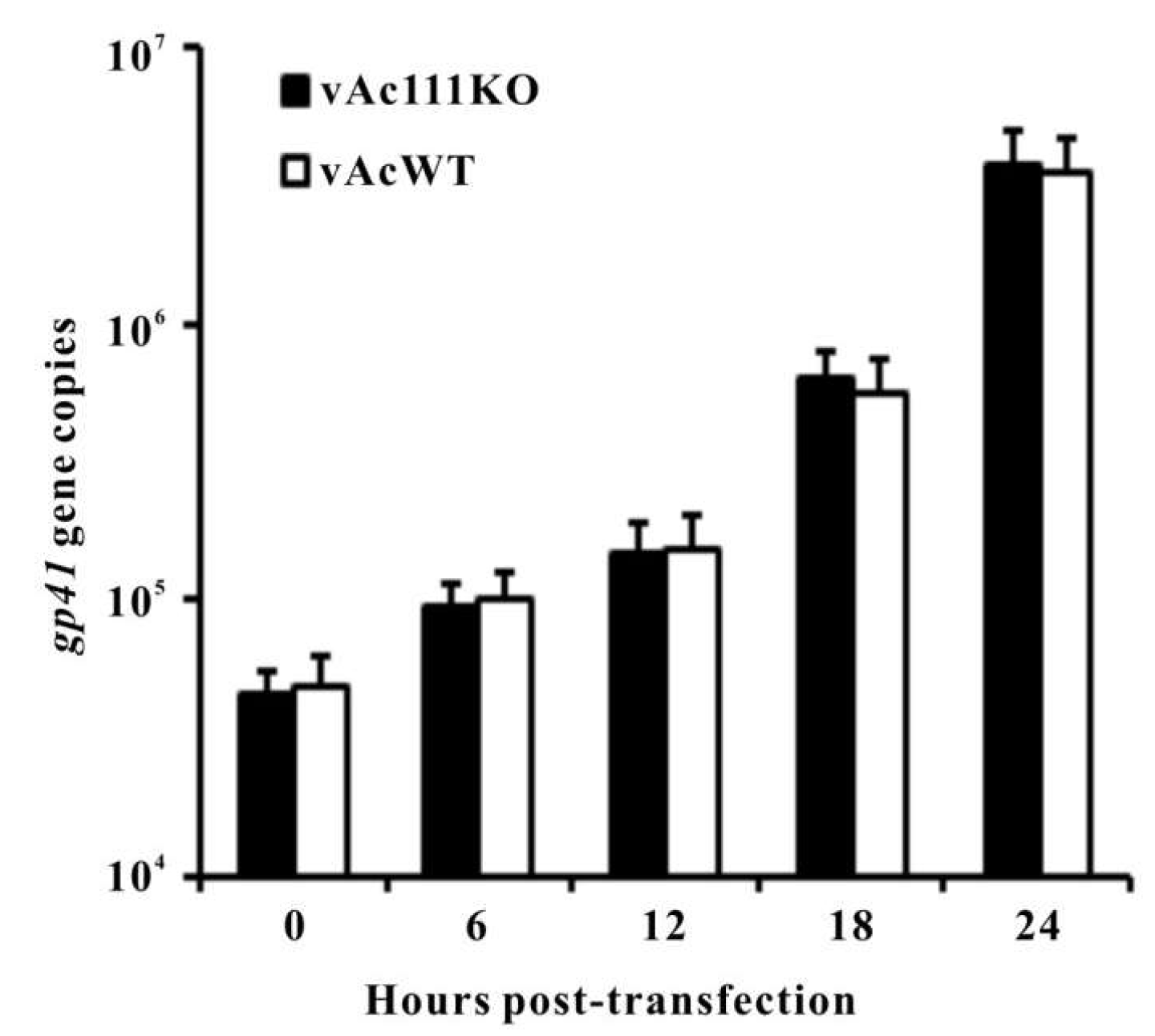
3.4. Ac111 is Not Required for Viral DNA Replication
3.5. Ac111 is Not Essential for Virus Morphogenesis
3.6. Ac111 Plays a Role in Oral Infectivity in a Host-Dependent Manner
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Herniou, E.A.; Olszewski, J.A.; Cory, J.S.; O’Reilly, D.R. The genome sequence and evolution of baculoviruses. Annu. Rev. Entomol. 2003, 48, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Zanotto, A.P.M.; Kessing, B.D.; Maruniak, J.E. Phylogenetic interrelationships among baculoviruses: Evolutionary rates and host associations. J. Invertebr. Pathol. 1993, 62, 147–164. [Google Scholar] [CrossRef]
- Ayres, M.D.; Howard, S.C.; Kuzio, J.; Lopez-Ferber, M.; Possee, R.D. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Virology 1994, 202, 586–605. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, M.J.; Miele, S.A.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The ac53, ac78, ac101, and ac103 genes are newly discovered core genes in the family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; van Oers, M.M.; Donly, B.C.; Erlandson, M.A.; Hegedus, D.D.; Theilmann, D.A. Autographa californica multiple nucleopolyhedrovirus AC83 is a per os infectivity factor (PIF) protein required for occlusion-derived virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV envelopes. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Huang, Z.; Wei, D.; Hu, Z.; Yang, K.; Pang, Y. Identification of Autographa californica nucleopolyhedrovirus ac93 as a core gene and its requirement for intranuclear microvesicle formation and nuclear egress of nucleocapsids. J. Virol. 2011, 85, 11664–11674. [Google Scholar] [CrossRef] [PubMed]
- Braunagel, S.C.; Summers, M.D. Autographa californica nuclear polyhedrosis virus, PDV, and ECV viral envelopes and nucleocapsids: Structural proteins, antigens, lipid and fatty acid profiles. Virology 1994, 202, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.; Arif, B.M. The baculoviruses occlusion-derived virus: Virion structure and function. Adv. Virus Res. 2007, 69, 99–165. [Google Scholar] [PubMed]
- Blissard, G.W.; Wenz, J.R. Baculovirus gp64 envelope glycoprotein is sufficient to mediate pH-dependent membrane fusion. J. Virol. 1992, 66, 6829–6835. [Google Scholar] [PubMed]
- Braunagel, S.C.; Summers, M.D. Molecular biology of the baculovirus occlusion-derived virus envelope. Curr. Drug Targets 2007, 8, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Nie, Y.; Harris, S.; Erlandson, M.A.; Theilmann, D.A. Autographa californica multiple nucleopolyhedrovirus core gene ac96 encodes a per os infectivity factor (pif-4). J. Virol. 2009, 83, 12569–12578. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Nie, Y.; Wang, Q.; Deng, F.; Wang, R.; Wang, H.; Vlak, J.M.; Chen, X.; Hu, Z. Open reading frame 132 of Helicoverpa armigera nucleopolyhedrovirus encodes a functional per os infectivity factor (PIF-2). J. Gen. Virol. 2006, 87, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, P.; Kuzio, J.; Williams, G.V.; Wilson, J.A. Analysis of p74, a PDV envelope protein of Autographa californica nucleopolyhedrovirus required for occlusion body infectivity in vivo. J. Gen. Virol. 1997, 78, 3091–3100. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Sparks, W.O.; Bonning, B.C. Autographa californica multiple nucleopolyhedrovirus ODV-E56 envelope protein is required for oral infectivity and can be substituted functionally by Rachiplusia ou multiple nucleopolyhedrovirus ODV-E56. J. Gen. Virol. 2010, 91, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Jiantao, L.; Zhu, L.; Zhang, S.; Deng, Z.; Huang, Z.; Yuan, M.; Wu, W.; Yang, K. The Autographa californica multiple nucleopolyhedrovirus ac110 gene encodes a new per os infectivity factor. Virus Res. 2016, 221, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Kikhno, I.; Gutierrez, S.; Croizier, L.; Croizier, G.; Ferber, M.L. Characterization of pif, a gene required for the per os infectivity of Spodoptera littoralis nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 3013–3022. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Fang, M.; Erlandson, M.A.; Theilmann, D.A. Analysis of the Autographa californica multiple nucleopolyhedrovirus overlapping gene pair lef3 and ac68 reveals that ac68 is a per os infectivity factor and that lef3 is critical, but not essential, for virus replication. J. Virol. 2012, 86, 3985–3994. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, T.; Washburn, J.O.; Sitapara, R.; Sid, E.; Volkman, L.E. Specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of Heliothis virescens larvae is mediated by products of pif genes ac119 and ac022 but not by ac115. J. Virol. 2005, 79, 15258–15264. [Google Scholar] [CrossRef] [PubMed]
- Pijlman, G.P.; Pruijssers, A.J.; Vlak, J.M. Identification of pif-2, a third conserved baculovirus gene required for per os infection of insects. J. Gen. Virol. 2003, 84, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Sparks, W.O.; Harrison, R.L.; Bonning, B.C. Autographa californica multiple nucleopolyhedrovirus ODV-E56 is a per os infectivity factor, but is not essential for binding and fusion of occlusion-derived virus to the host midgut. Virology 2011, 409, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Chen, L.; Guo, A.; Yu, S.; Yang, R.; Wu, X. The Bombyx mori nucleopolyhedrovirus (BmNPV) ODV-E56 envelope protein is also a per os infectivity factor. Virus Res. 2011, 155, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wang, W.; Wang, Y.; Yuan, M.; Yang, K. The baculovirus core gene ac83 is required for nucleocapsid assembly and per os infectivity of Autographa californica nucleopolyhedrovirus. J. Virol. 2013, 87, 10573–10586. [Google Scholar] [CrossRef] [PubMed]
- Simon, O.; Palma, L.; Williams, T.; Lopez-Ferber, M.; Caballero, P. Analysis of a naturally-occurring deletion mutant of Spodoptera frugiperda multiple nucleopolyhedrovirus reveals sf58 as a new per os infectivity factor of lepidopteran-infecting baculoviruses. J. Invertebr. Pathol. 2012, 109, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Detvisitsakun, C.; Cain, E.L.; Passarelli, A.L. The Autographa californica M nucleopolyhedrovirus fibroblast growth factor accelerates host mortality. Virology 2007, 365, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, R.; Popham, H.J.; Straschil, U.; Goulding, D.; O’Reilly, D.R.; Olszewski, J.A. Characterization of two Autographa californica nucleopolyhedrovirus proteins, Ac145 and Ac150, which affect oral infectivity in a host-dependent manner. J. Virol. 2004, 78, 6439–6448. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Lan, D.; Zhao, S.; Liu, L.; Xue, Y.; Zhang, Y.; Wang, Y.; Chen, X. The Ac124 protein is not essential for the propagation of Autographa californica multiple nucleopolyhedrovirus, but it is a viral pathogenicity factor. Arch. Virol. 2015, 160, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.; Li, Z.; Yuan, M.; Feng, G.; Yu, Q.; Yang, K.; Pang, Y. ac18 is not essential for the propagation of Autographa californica multiple nucleopolyhedrovirus. Virology 2007, 367, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhou, Y.; Lei, C.; Sun, X. Autographa californica multiple nucleopolyhedrovirus orf114 is not essential for virus replication in vitro, but its knockout reduces per os infectivity in vivo. Virus Genes 2012, 45, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Chen, L.; Hu, X.; Yu, S.; Yang, R.; Wu, X. Autographa californica multiple nucleopolyhedrovirus odv-e66 is an essential gene required for oral infectivity. Virus Res. 2011, 158, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Ohkawa, T.; Washburn, J.O.; Volkman, L.E. Effects of Ac150 on virulence and pathogenesis of Autographa californica multiple nucleopolyhedrovirus in noctuid hosts. J. Gen. Virol. 2005, 86, 1619–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.R.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissard, G.W. The transcriptome of the baculovirus Autographa californica multiple nucleopolyhedrovirus in Trichoplusia ni cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef] [PubMed]
- Ono, C.; Kamagata, T.; Taka, H.; Sahara, K.; Asano, S.; Bando, H. Phenotypic grouping of 141 BmNPVs lacking viral gene sequences. Virus Res. 2012, 165, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucl. Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Anderson, J.B.; Derbyshire, M.K.; DeWeese-Scott, C.; Gonzales, N.R.; Gwadz, M.; Hao, L.; He, S.; Hurwitz, D.I.; Jackson, J.D.; et al. CDD: A conserved domain database for interactive domain family analysis. Nucl. Acids Res. 2007, 35, D237–D240. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Sternberg, M.J. Protein structure prediction on the Web: A case study using the Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci.USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.B.; Nicholas, H.B.; Deerfield, D.W. GeneDoc: Analysis and visualization of genetic variation. Embnew News 1997, 4, 14. [Google Scholar]
- Hara, K.; Funakoshi, M.; Tsuda, K.; Kawarabata, T. New Spodoptera exigua cell lines susceptible to Spodoptera exigua nuclear polyhedrosis virus. Vitro Cell. Dev. Biol. Anim. 1993, 29A, 904–907. [Google Scholar] [CrossRef]
- Luckow, V.A.; Lee, S.C.; Barry, G.F.; Olins, P.O. Efficient generation of infectious recombinant baculoviruses by site-specific transposon-mediated insertion of foreign genes into a baculovirus genome propagated in Escherichia coli. J. Virol. 1993, 67, 4566–4579. [Google Scholar] [PubMed]
- Li, G.; Pang, Y.; Chen, Q.; Su, Z.; Wen, X. Studies on the artificial diet for beet armyworm, Spodoptera exigua. Chin. J. Biol. Control 2002, 18, 132–134. [Google Scholar]
- O’Reilly, D.R.; Miller, L.K.; Luckow, V.A. Baculovirus Expression Vectors: A Laboratory Manual; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Yuan, M.; Wu, W.; Liu, C.; Wang, Y.; Hu, Z.; Yang, K.; Pang, Y. A highly conserved baculovirus gene p48 (ac103) is essential for BV production and ODV envelopment. Virology 2008, 379, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lin, T.; Pan, L.; Yu, M.; Li, Z.; Pang, Y.; Yang, K. Autographa californica multiple nucleopolyhedrovirus nucleocapsid assembly is interrupted upon deletion of the 38k gene. J. Virol. 2006, 80, 11475–11485. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Long, Z.; Qiu, J.; Yuan, M.; Li, G.; Yang, K. An ac34 deletion mutant of Autographa californica nucleopolyhedrovirus exhibits delayed late gene expression and a lack of virulence in vivo. J. Virol. 2012, 86, 10432–10443. [Google Scholar] [CrossRef] [PubMed]
- Li, S.N.; Wang, J.Y.; Yuan, M.J.; Yang, K. Disruption of the baculovirus core gene ac78 results in decreased production of multiple nucleocapsid-enveloped occlusion-derived virions and the failure of primary infection in vivo. Virus Res. 2014, 191, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Passarelli, A.L. Autographa californica multiple nucleopolyhedrovirus Ac92 (ORF92, P33) is required for budded virus production and multiply enveloped occlusion-derived virus formation. J. Virol. 2010, 84, 12351–12361. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.; vanBeek, N.; Wood, H. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses innoctuid larvae. J. Invertebr. Pathol. 1986, 48, 187–192. [Google Scholar] [CrossRef]
- Li, S.; Department of Biology, Zhaoqing University, China. Viral growth curveanalyses in Se301 cells and plaque assays. 2018.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Applications | Primer | Sequence |
|---|---|---|
| Amplification of ac111 5′ flanking sequences | ac111PF1 | 5′-GAGCTCGTTTCCATAATAATGAGGATACGCT-3′ |
| ac111PR1 | 5′-GGATCCATCCATAGTGCAGCACAGGCAAT-3′ | |
| Amplification of ac111 3′ flanking sequences | ac111PF2 | 5′-CTGCAGATCGGTGTACGAAGATGTCACATATA-3′ |
| ac111PR2 | 5′-AAGCTTAGATTATTTTAATTTGTGAACTCGTACC-3′ | |
| Amplification of the 5′ end of the ac111 native promoter and ORF | ac111PF3 | 5′-GAGCTCTACAAAACGATGCATTTATAGCGC-3′ |
| Amplification of the 3′ end of ac111 tagged with an HA | ac111PR3 | 5′-GGATCCTTAGGCGTAATCTGGGACGTCGTATGGGTA- TTTATATTTGTTTTCTTTGTTATAACCG-3′ |
| Amplification of the 5′ end of ac111 target deletion region | ac111PF4 | 5′-AATTATTCGGTGCAAAATTTTTACAAC-3′ |
| Amplification of the 3′ end of ac111 target deletion region | ac111PR4 | 5′-TTCTTGATGTTACCATCGTGAAGCGTTG-3′ |
| Amplification of Cm cassette | CmPF | 5′-CATGTCTGGATCCTTCGAATAAATACCTGTGACGG-3′ |
| CmPR | 5′-GGATTCTAAACCAGCAATAGACATAAGCGGC-3’ | |
| Real-time PCR, specific for AcMNPV DNA | gp41-F | 5′-CGTAGTGGTAGTAATCGCCGC-3′ |
| gp41-R | 5′-AGTCGAGTCGCGTCGCTTT-3′ |
| Inoculation Methods and Viruses or Control | Insect Larvae a | Dose | Mortality at 7 Days p.i. (%) | Dose-Mortality Regression | LT50 (hr) (95% Confidence Interval) | ||
|---|---|---|---|---|---|---|---|
| LD50c (95% Confidence Interval) | slope ± SE | Chi2d/dfe | |||||
| Injection | |||||||
| vAcWT | T. ni | 1 TCID50 | 63 | ||||
| vAc111:HA | T. ni | 1 TCID50 | 63 | ||||
| vAc111KO | T. ni | 1 TCID50 | 67 | ||||
| vAcWT | S. exigua | 2 TCID50 | 70 | ||||
| vAc111:HA | S. exigua | 2 TCID50 | 67 | ||||
| vAc111KO | S. exigua | 2 TCID50 | 70 | ||||
| H2O | T. ni S. exigua | 3.3b | |||||
| Per os | |||||||
| vAcWT | T. ni | 54 (34–84) | 1.33 ± 0.22 | 4.80/3 | 85 (81–90) | ||
| vAc111:HA | T. ni | 59 (36–92) | 1.37 ± 0.22 | 4.76/3 | 86 (82–91) | ||
| vAc111KO | T. ni | 300 (187–482) | 1.39 ± 0.28 | 1.82/2 | 106 (101–112) | ||
| vAcWT | S. exigua | 4535 (2823–7283) | 1.13 ± 0.18 | 7.72/3 | 140 (126–155) | ||
| vAc111:HA | S. exigua | 4650 (2989–6877) | 1.42 ± 0.20 | 2.59/3 | 141 (129–154) | ||
| vAc111KO | S. exigua | 5006 (2975–8423) | 1.51 ± 0.21 | 2.21/3 | 148 (133–164) | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Li, L.; Zhao, H.; Liu, W. Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner. Viruses 2018, 10, 527. https://doi.org/10.3390/v10100527
Li S, Li L, Zhao H, Liu W. Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner. Viruses. 2018; 10(10):527. https://doi.org/10.3390/v10100527
Chicago/Turabian StyleLi, Sainan, Lu Li, Haizhou Zhao, and Wenhua Liu. 2018. "Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner" Viruses 10, no. 10: 527. https://doi.org/10.3390/v10100527
APA StyleLi, S., Li, L., Zhao, H., & Liu, W. (2018). Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner. Viruses, 10(10), 527. https://doi.org/10.3390/v10100527