Thiol-Activated Hydrogen Sulfide Donors Antiviral and Anti-Inflammatory Activity in Respiratory Syncytial Virus Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Virus Preparation and Titration
2.4. Determination of Lactate Dehydrogenase Activity
2.5. Quantitative Real-Time PCR
2.6. Western Blotting
2.7. Animal Studies
2.8. Statistical Analysis
3. Results
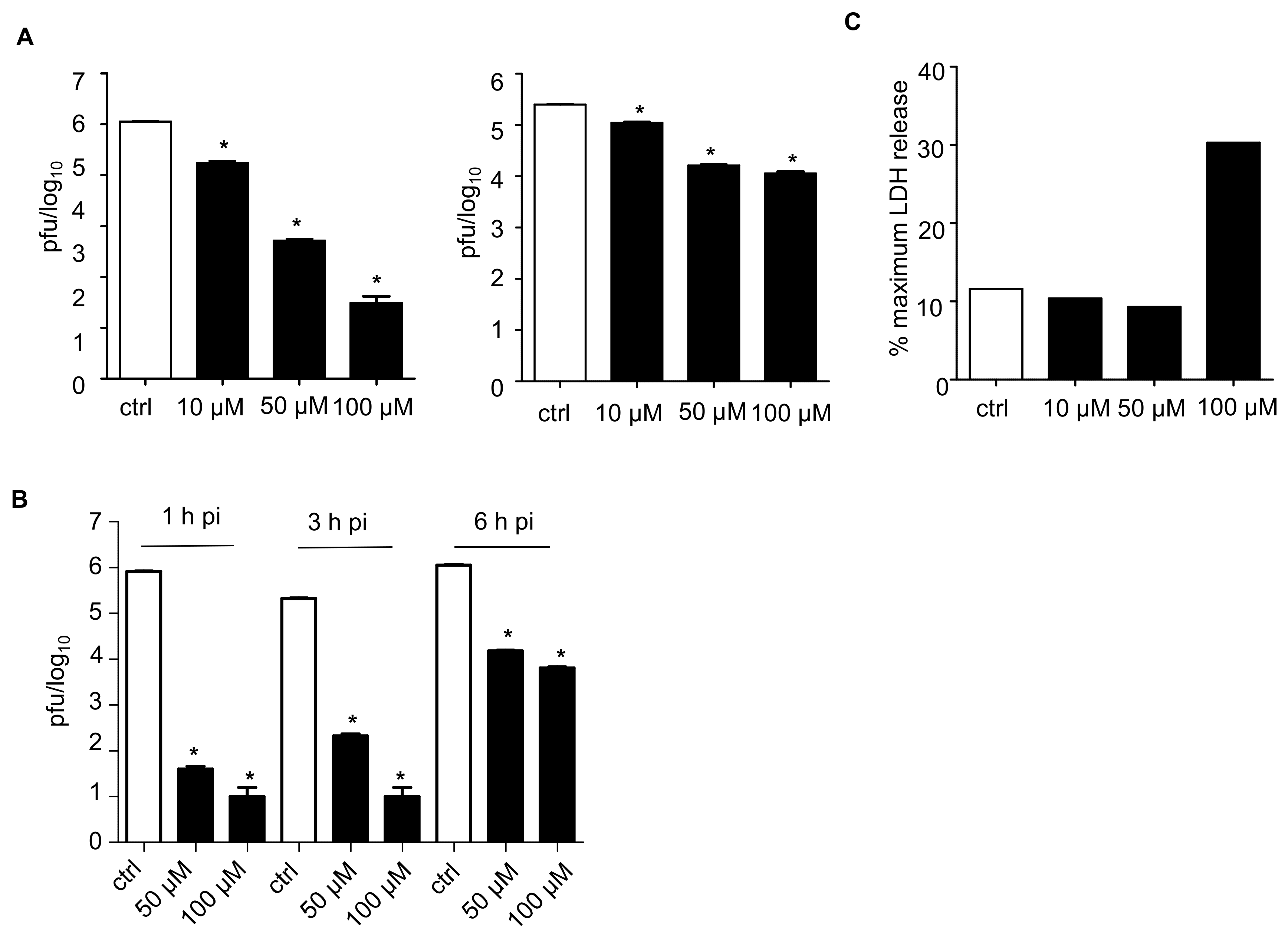
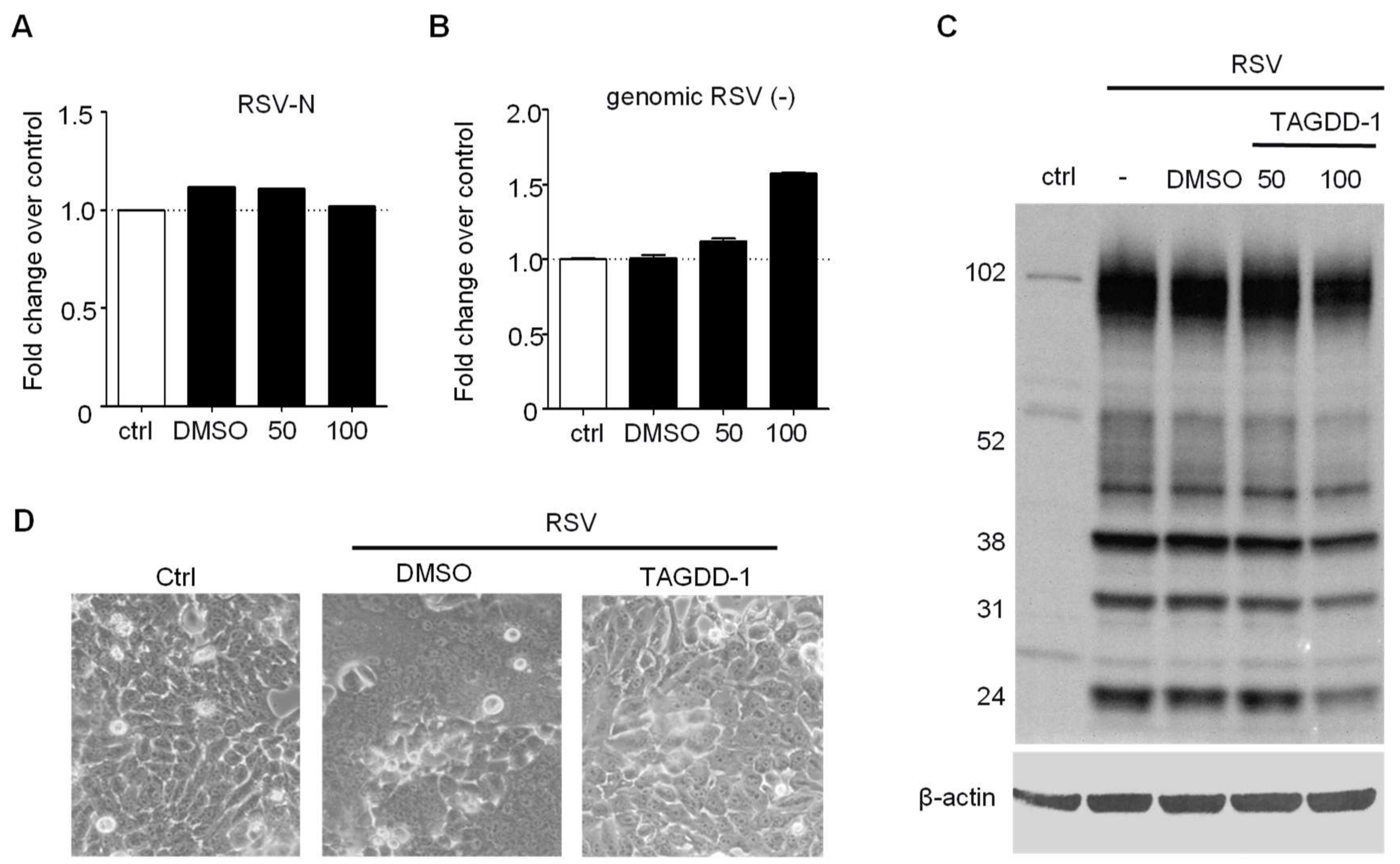
3.1. Effect of TAGDDs on RSV Replication In Vitro
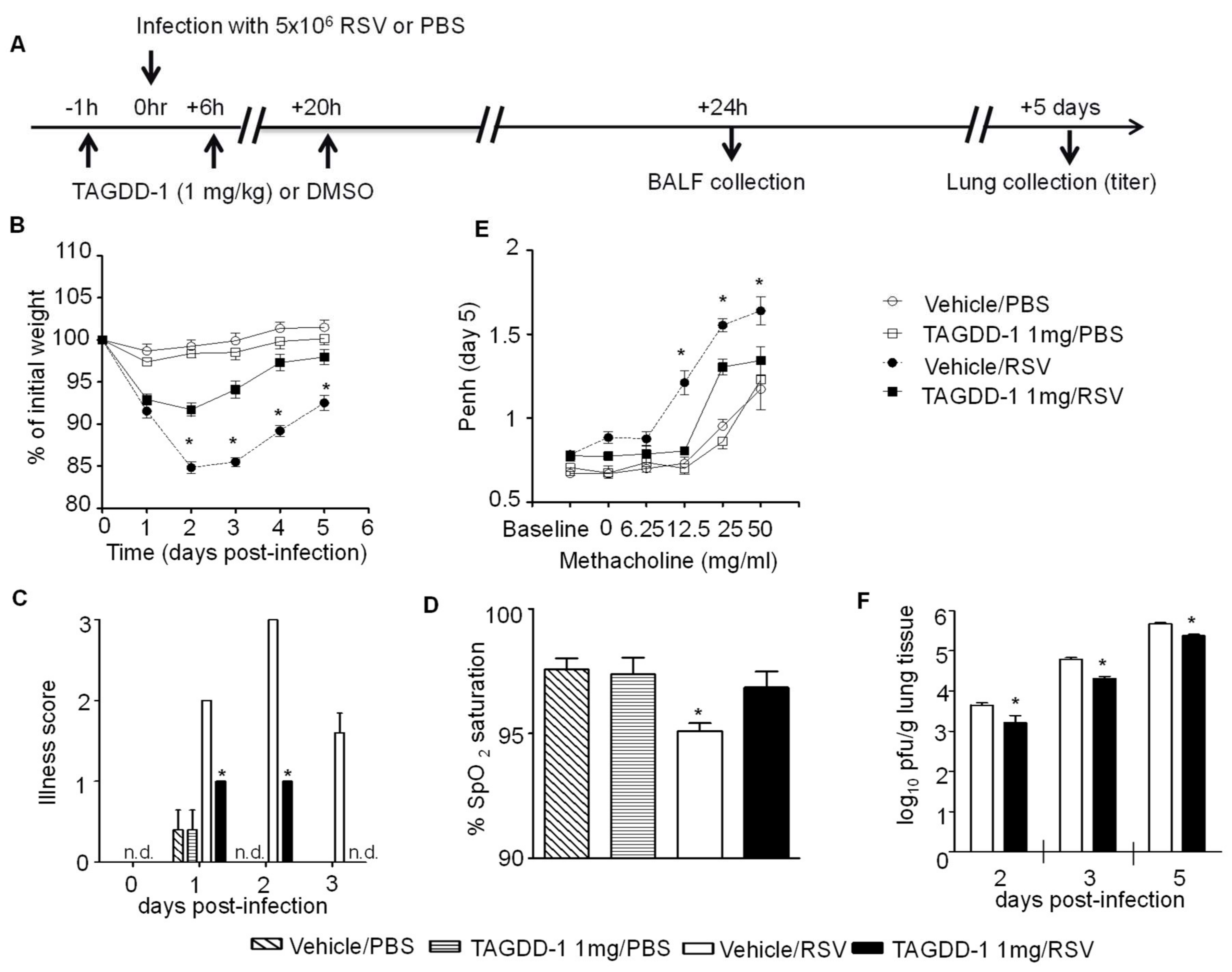
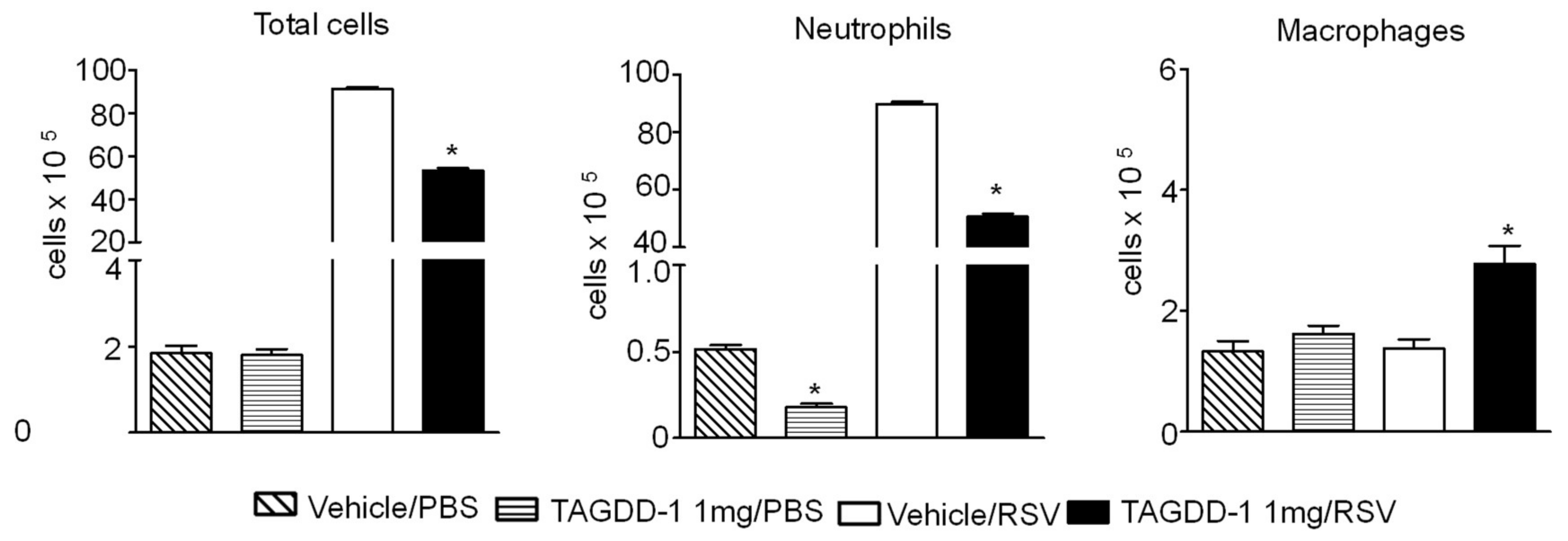
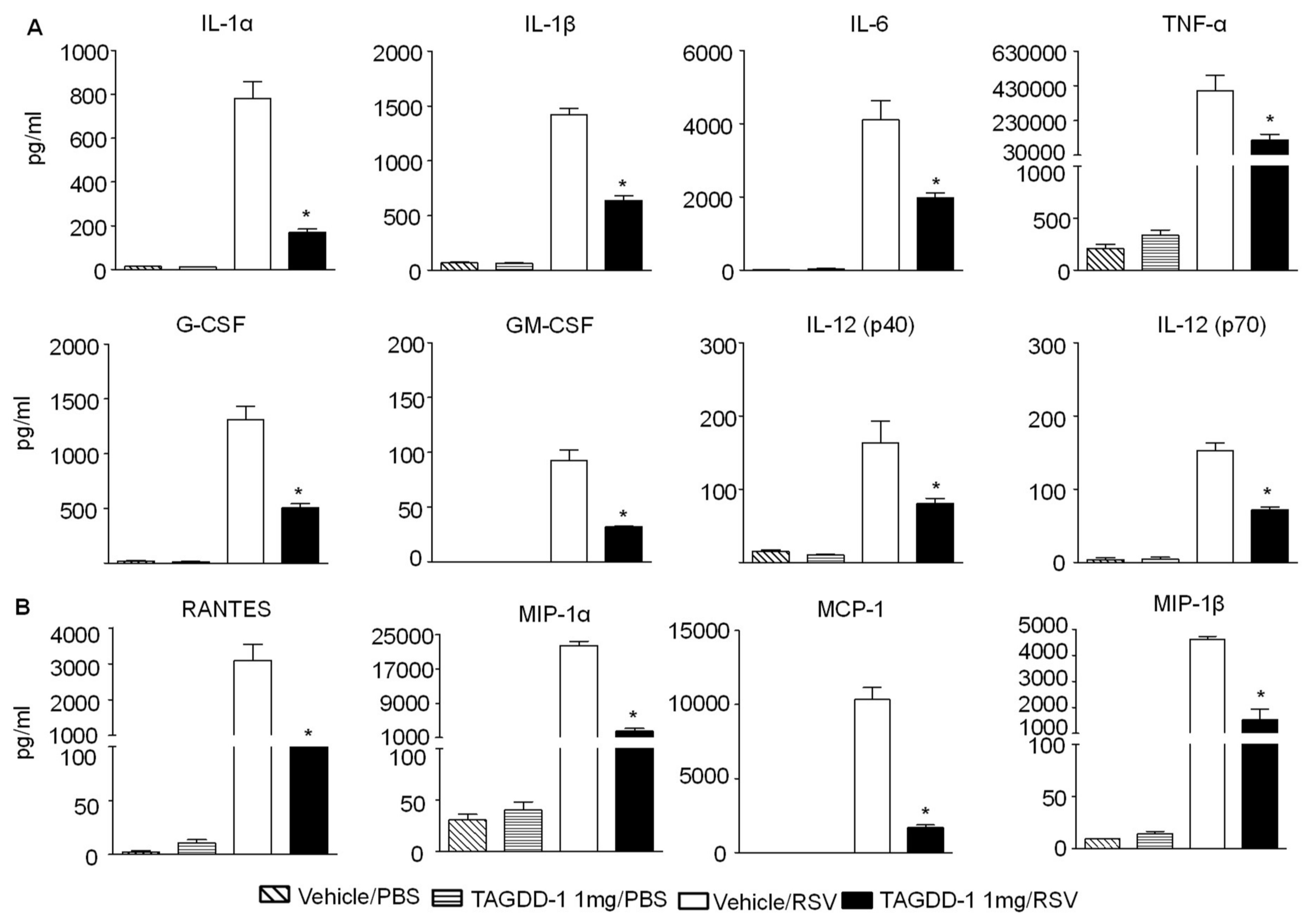
3.2. Effect of TAGDD-1 on RSV Infection In Vivo
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Wang, R. The message in the air: Hydrogen sulfide metabolism in chronic respiratory diseases. Respir. Physiol. Neurobiol. 2012, 184, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, G.; Wondimu, T.; Ross, B.; Wang, R. Hydrogen sulfide and asthma. Exp. Physiol. 2011, 96, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Signaling pathways for the vascular effects of hydrogen sulfide. Curr. Opin. Nephrol. Hypertens. 2011, 20, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 2007, 6, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Kashfi, K.; Olson, K.R. Biology and therapeutic potential of hydrogen sulfide and hydrogen sulfide-releasing chimeras. Biochem. Pharmacol. 2013, 85, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Pacheco, A.; Xian, M. Medicinal chemistry: Insights into the development of novel H2S donors. Handb. Exp. Pharmacol. 2015, 230, 365–388. [Google Scholar] [PubMed]
- Li, H.; Ma, Y.; Escaffre, O.; Ivanciuc, T.; Komaravelli, N.; Kelley, J.P.; Coletta, C.; Szabo, C.; Rockx, B.; Garofalo, R.P.; et al. Role of hydrogen sulfide in paramyxovirus infections. J. Virol. 2015, 89, 5557–5568. [Google Scholar] [CrossRef] [PubMed]
- Ivanciuc, T.; Sbrana, E.; Ansar, M.; Bazhanov, N.; Szabo, C.; Casola, A.; Garofalo, R.P. Hydrogen sulfide is an antiviral and antiinflammatory endogenous gasotransmitter in the airways. Role in respiratory syncytial virus infection. Am. J. Respir. Cell Mol. Biol. 2016, 55, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Weinberg, G.A.; Iwane, M.K.; Blumkin, A.K.; Edwards, K.M.; Staat, M.A.; Auinger, P.; Griffin, M.R.; Poehling, K.A.; Erdman, D.; et al. The burden of respiratory syncytial virus infection in young children. N. Engl. J. Med. 2009, 360, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.H.; Moore, P.K. Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): New insights into the biology of hydrogen sulfide. Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Bazhanov, N.; Escaffre, O.; Freiberg, A.N.; Garofalo, R.P.; Casola, A. Broad-range antiviral activity of hydrogen sulfide against highly pathogenic RNA viruses. Sci. Rep. 2017, 7, 41029. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Kang, J.; Park, C.M.; Bagdon, P.E.; Peng, B.; Xian, M. Thiol-activated gem-dithiols: A new class of controllable hydrogen sulfide donors. Org. Lett. 2014, 16, 4536–4539. [Google Scholar] [CrossRef] [PubMed]
- Ueba, O. Respiratory syncytial virus: I. concentration and purification of the infectious virus. Acta Med. Okayama 1978, 32, 265–272. [Google Scholar] [PubMed]
- Olszewska-Pazdrak, B.; Casola, A.; Saito, T.; Alam, R.; Crowe, S.E.; Mei, F.; Ogra, P.L.; Garofalo, R.P. Cell-specific expression of RANTES, MCP-1, and MIP-1alpha by lower airway epithelial cells and eosinophils infected with respiratory syncytial virus. J. Virol. 1998, 72, 4756–4764. [Google Scholar] [PubMed]
- Kisch, A.L.; Johnson, K.M. A plaque assay for respiratory syncytial virus. Proc. Soc. Exp. Biol. Med. 1963, 112, 583. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Plata, A.; Baron, S.; Poast, J.S.; Adegboyega, P.A.; Casola, A.; Garofalo, R.P. Activity and regulation of alpha interferon in respiratory syncytial virus and human metapneumovirus experimental infections. J. Virol. 2005, 79, 10190–10199. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.M.; Guerrero-Plata, A.; Suarez-Real, G.; Adegboyega, P.A.; Colasurdo, G.N.; Khan, A.M.; Garofalo, R.P.; Casola, A. Antioxidant Treatment Ameliorates Respiratory Syncytial Virus-induced Disease and Lung Inflammation. Am. J. Respir. Crit. Care Med. 2006, 174, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.R.; Eurich, D.T.; Gamble, J.M.; Senthilselvan, A.; Marrie, T.J. Oxygen saturations less than 92% are associated with major adverse events in outpatients with pneumonia: A population-based cohort study. Clin. Infect. Dis. 2011, 52, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, D.; Teijaro, J.R.; Farber, D.L. Pulse-oximetry accurately predicts lung pathology and the immune response during influenza infection. Virology 2009, 390, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Davis, I.C.; Lazarowski, E.R.; Hickman-Davis, J.M.; Fortenberry, J.A.; Chen, F.P.; Zhao, X.; Sorscher, E.; Graves, L.M.; Sullender, W.M.; Matalon, S. Leflunomide prevents alveolar fluid clearance inhibition by respiratory syncytial virus. Am. J. Respir. Crit. Care Med. 2006, 173, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Haeberle, H.A.; Kuziel, W.A.; Dieterich, H.J.; Casola, A.; Gatalica, Z.; Garofalo, R.P. Inducible expression of inflammatory chemokines in respiratory syncytial virus-infected mice: Role of MIP-1alpha in lung pathology. J. Virol. 2001, 75, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Haeberle, H.A.; Casola, A.; Gatalica, Z.; Petronella, S.; Dieterich, H.J.; Ernst, P.B.; Brasier, A.R.; Garofalo, R.P. IkappaB Kinase Is a Critical Regulator of Chemokine Expression and Lung Inflammation in Respiratory Syncytial Virus Infection. J. Virol. 2004, 78, 2232–2241. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.L.; Garofalo, R.P.; Cron, S.G.; Hosakote, Y.M.; Atmar, R.L.; Macias, C.G.; Piedra, P.A. Immunopathogenesis of respiratory syncytial virus bronchiolitis. J. Infect. Dis. 2007, 195, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Jartti, T.; Mansbach, J.M.; Laham, F.R.; Jewell, A.M.; Espinola, J.A.; Piedra, P.A.; Camargo, C.A., Jr. Respiratory syncytial virus genomic load and disease severity among children hospitalized with bronchiolitis: Multicenter cohort studies in the United States and Finland. J. Infect. Dis. 2015, 211, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Devincenzo, J.P.; El Saleeby, C.M.; Bush, A.J. Respiratory syncytial virus load predicts disease severity in previously healthy infants. J. Infect. Dis. 2005, 191, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Production and physiological effects of hydrogen sulfide. Antioxid. Redox Signal. 2014, 20, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Dymock, B.W.; Moore, P.K. GYY4137, a novel water-soluble, H2S-releasing molecule. Methods Enzymol. 2015, 554, 143–167. [Google Scholar] [PubMed]
- Fullmer, J.J.; Khan, A.M.; Elidemir, O.; Chiappetta, C.; Stark, J.M.; Colasurdo, G.N. Role of cysteinyl leukotrienes in airway inflammation and responsiveness following RSV infection in BALB/c mice. Pediatr. Allergy Immunol. 2005, 16, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Hosakote, Y.M.; Liu, T.; Castro, S.M.; Garofalo, R.P.; Casola, A. Respiratory syncytial virus induces oxidative stress by modulating antioxidant enzymes. Am. J. Respir. Cell Mol. Biol. 2009, 41, 348–357. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazhanov, N.; Ivanciuc, T.; Wu, H.; Garofalo, M.; Kang, J.; Xian, M.; Casola, A. Thiol-Activated Hydrogen Sulfide Donors Antiviral and Anti-Inflammatory Activity in Respiratory Syncytial Virus Infection. Viruses 2018, 10, 249. https://doi.org/10.3390/v10050249
Bazhanov N, Ivanciuc T, Wu H, Garofalo M, Kang J, Xian M, Casola A. Thiol-Activated Hydrogen Sulfide Donors Antiviral and Anti-Inflammatory Activity in Respiratory Syncytial Virus Infection. Viruses. 2018; 10(5):249. https://doi.org/10.3390/v10050249
Chicago/Turabian StyleBazhanov, Nikolay, Teodora Ivanciuc, Haotian Wu, Matteo Garofalo, Jianming Kang, Ming Xian, and Antonella Casola. 2018. "Thiol-Activated Hydrogen Sulfide Donors Antiviral and Anti-Inflammatory Activity in Respiratory Syncytial Virus Infection" Viruses 10, no. 5: 249. https://doi.org/10.3390/v10050249