Susceptibility of Exopalaemon carinicauda to the Infection with Shrimp Hemocyte Iridescent Virus (SHIV 20141215), a Strain of Decapod Iridescent Virus 1 (DIV1)


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Viral Inoculum
2.3. Challenge Tests
2.4. Detection of DIV1 Using TaqMan-Probe-Based Real-Time Quantitative PCR (TaqMan qPCR)
2.5. Histopathological Examination
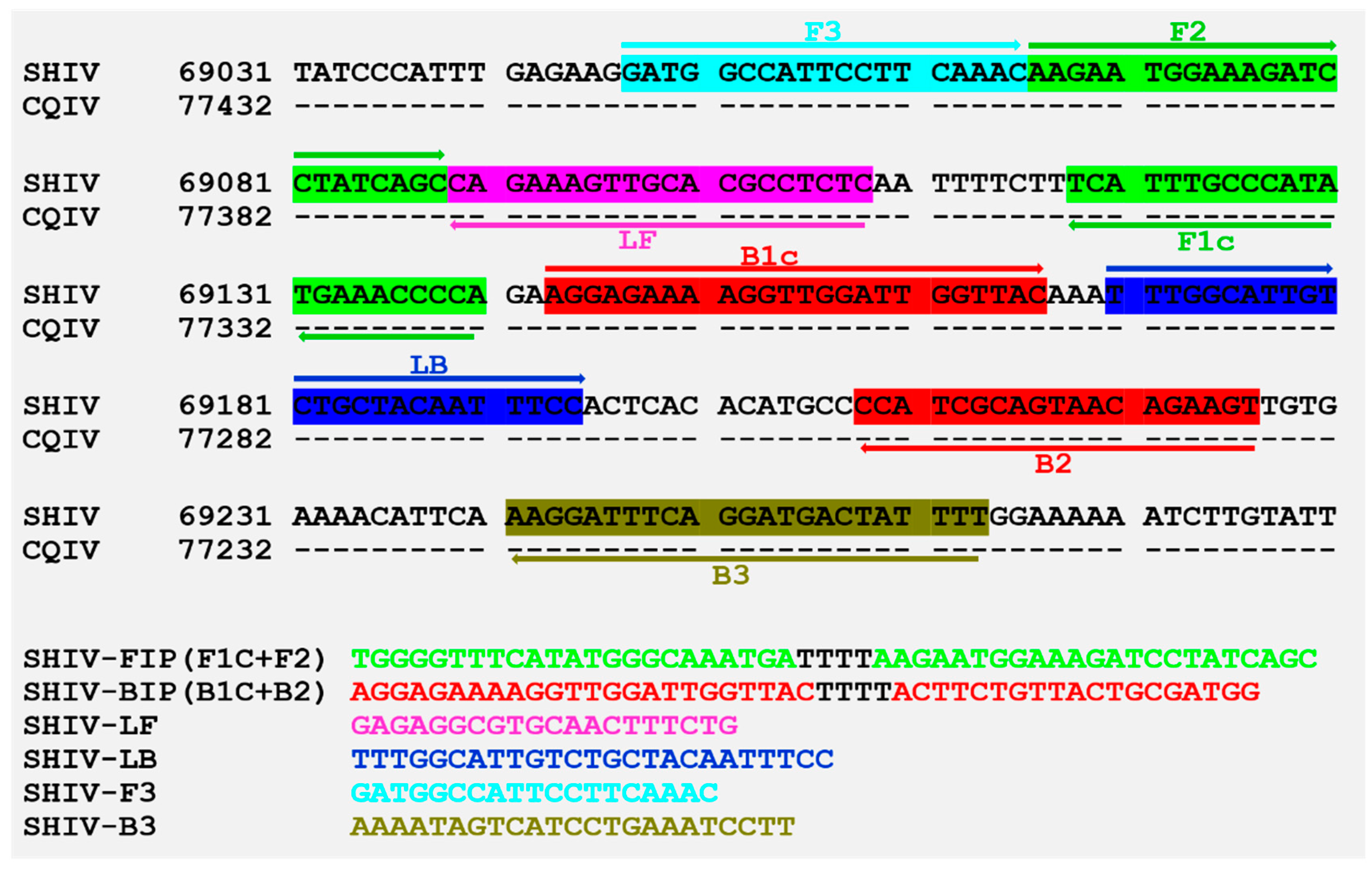
2.6. Loop-Mediated Isothermal Amplification (LAMP)
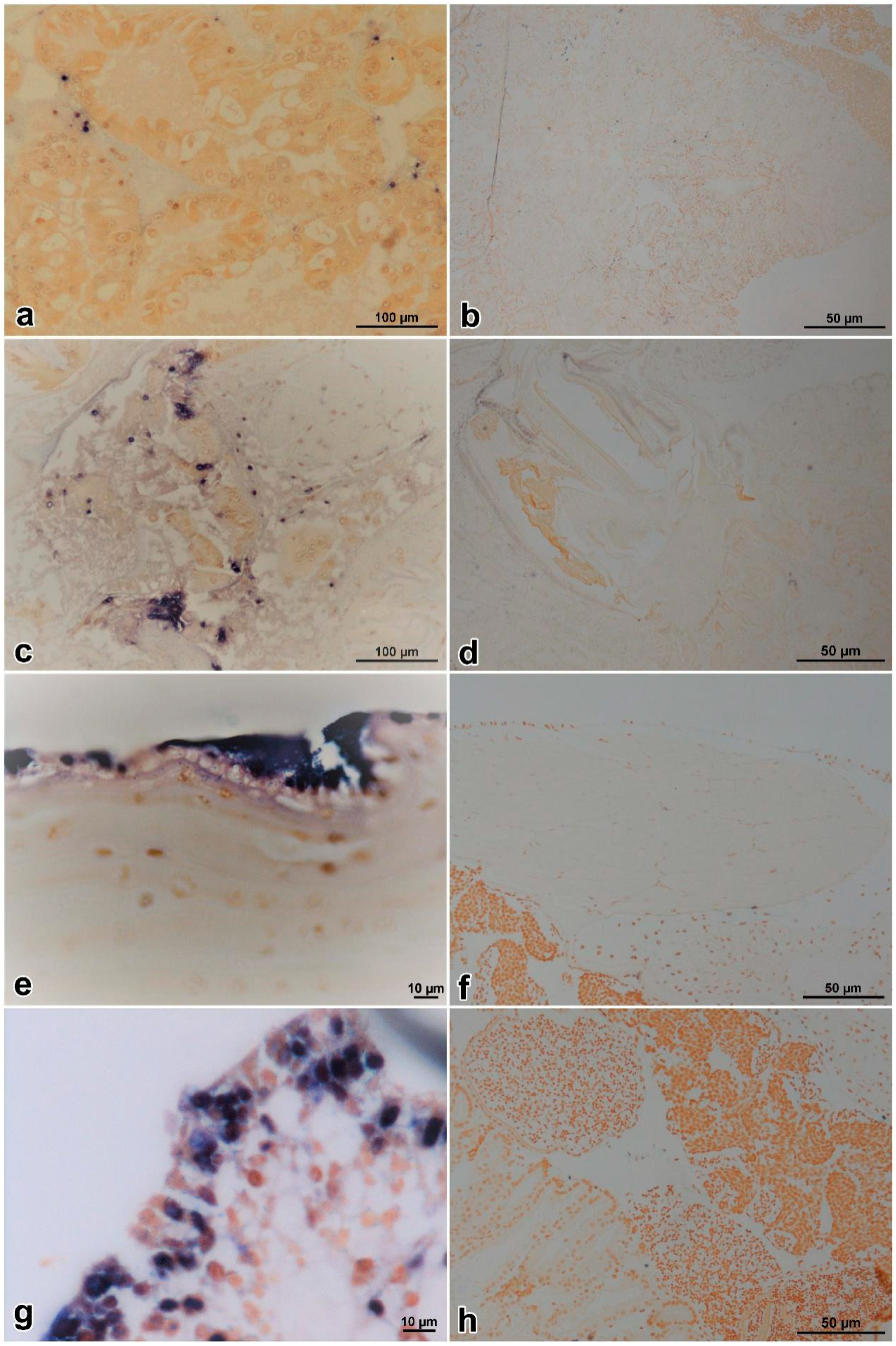
2.7. In Situ DIG-Labeling-Loop-Mediated DNA Amplification (ISDL)
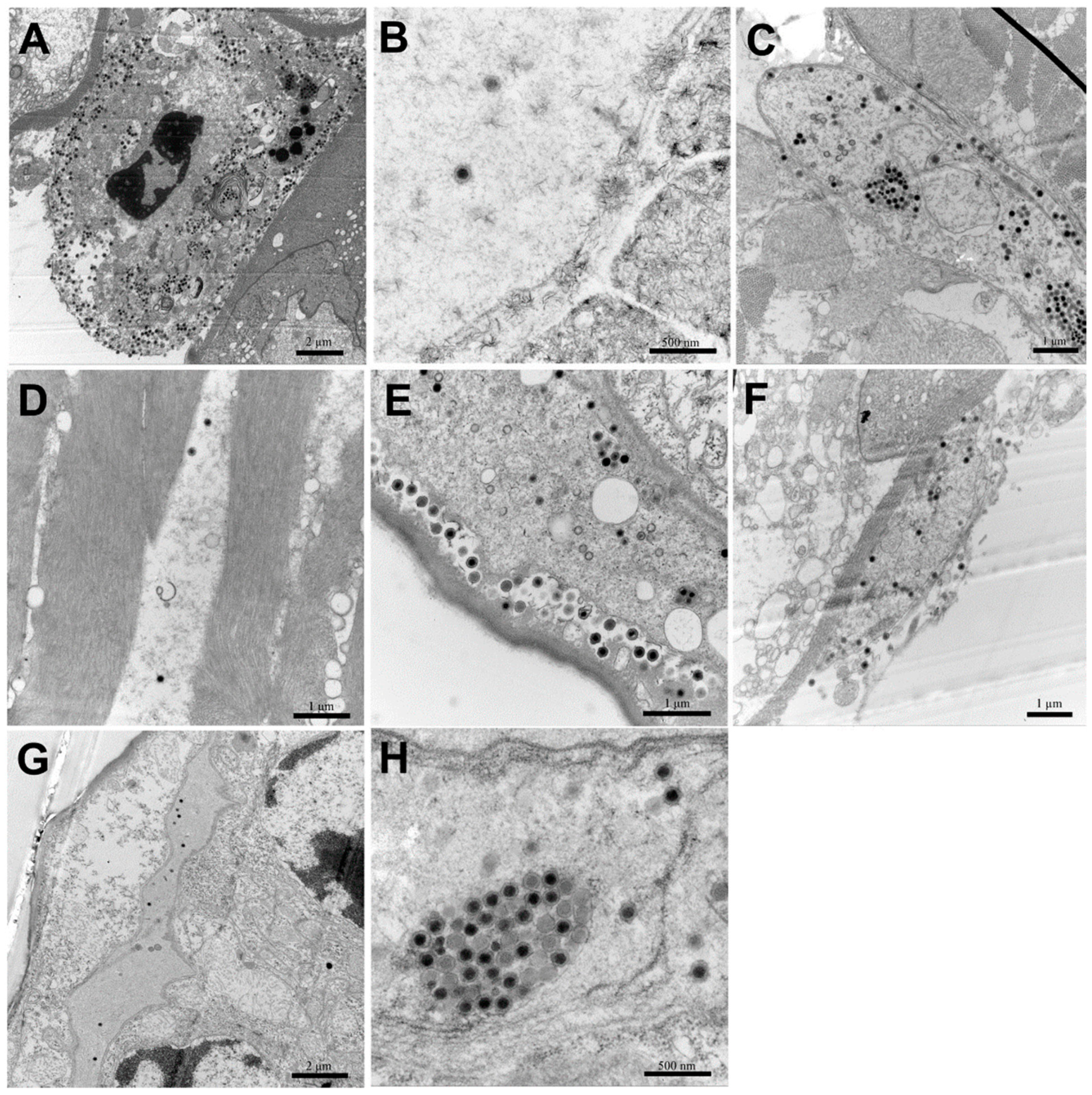
2.8. Transmission Electron Microscopy with Ultrathin Sections
3. Results
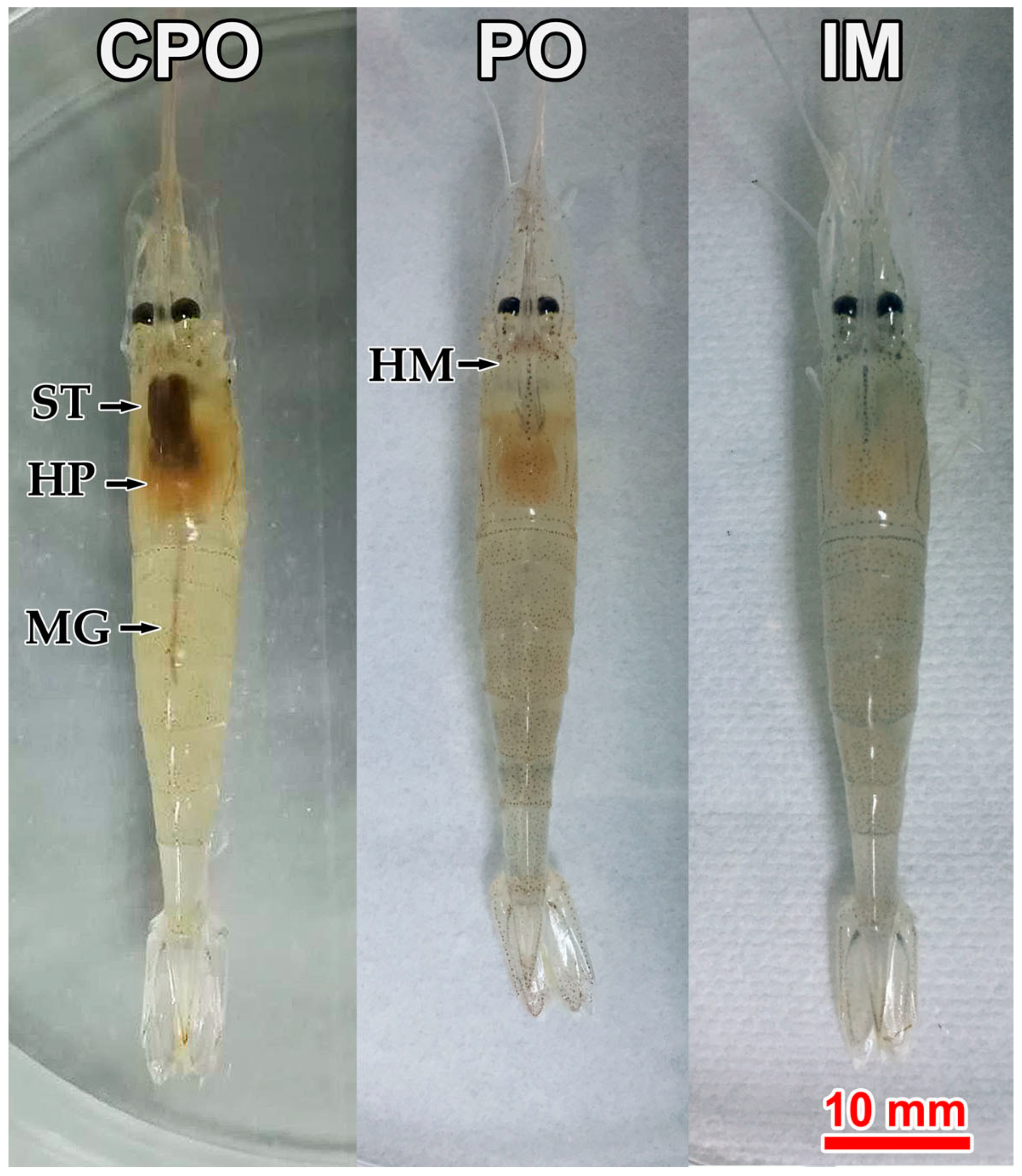
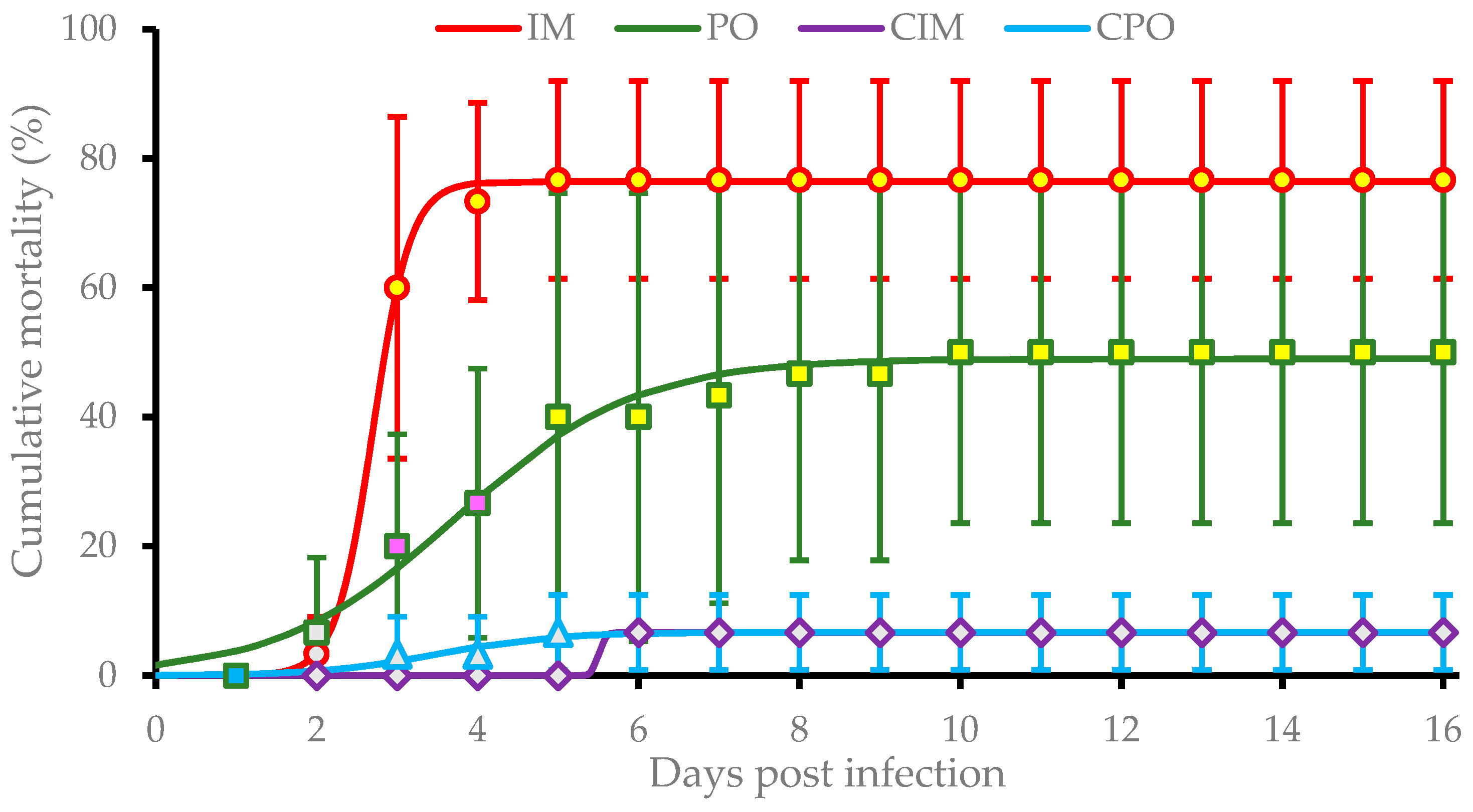
3.1. Clinical Signs and Cumulative Mortality
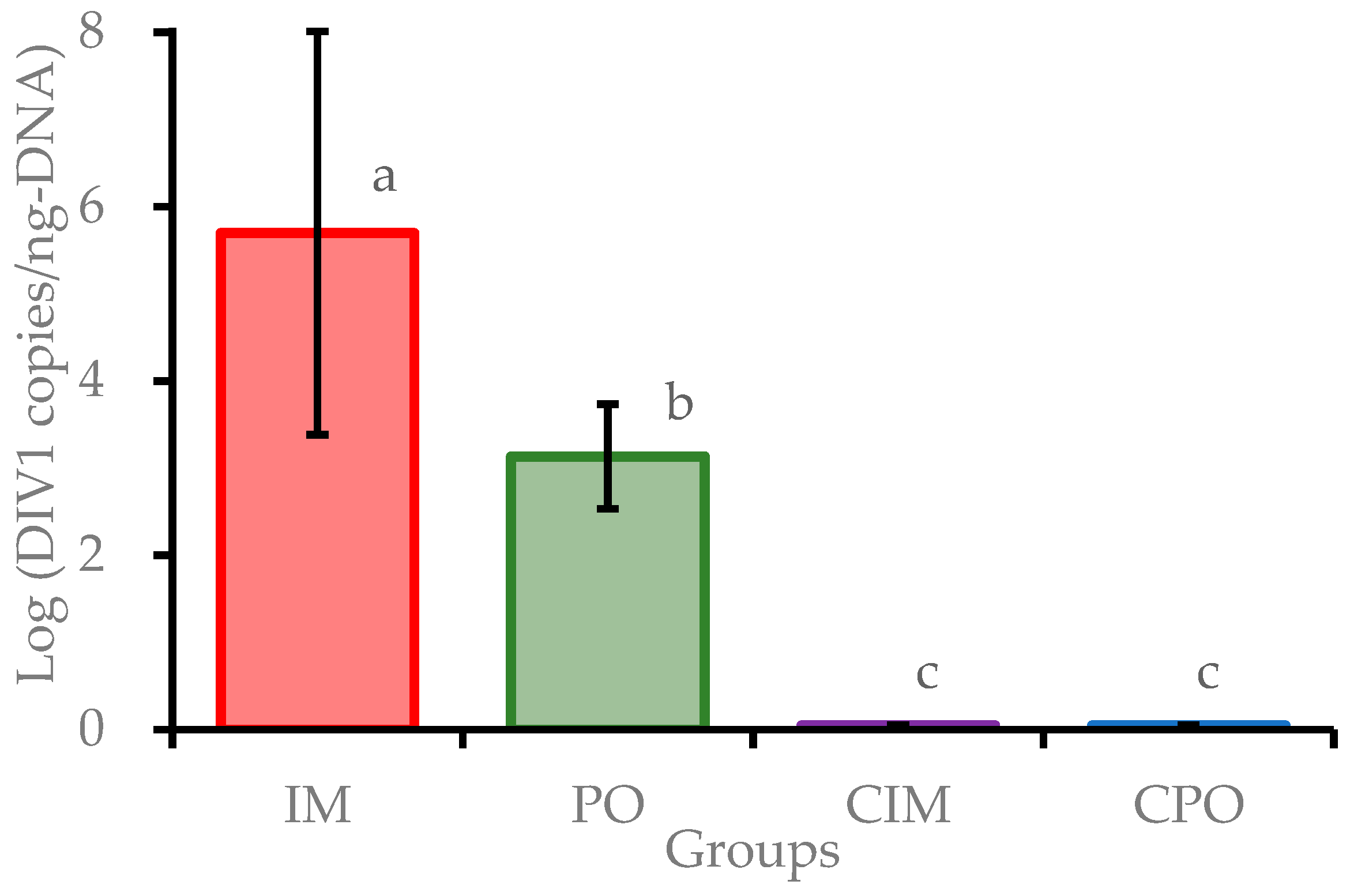
3.2. TaqMan qPCR Detection
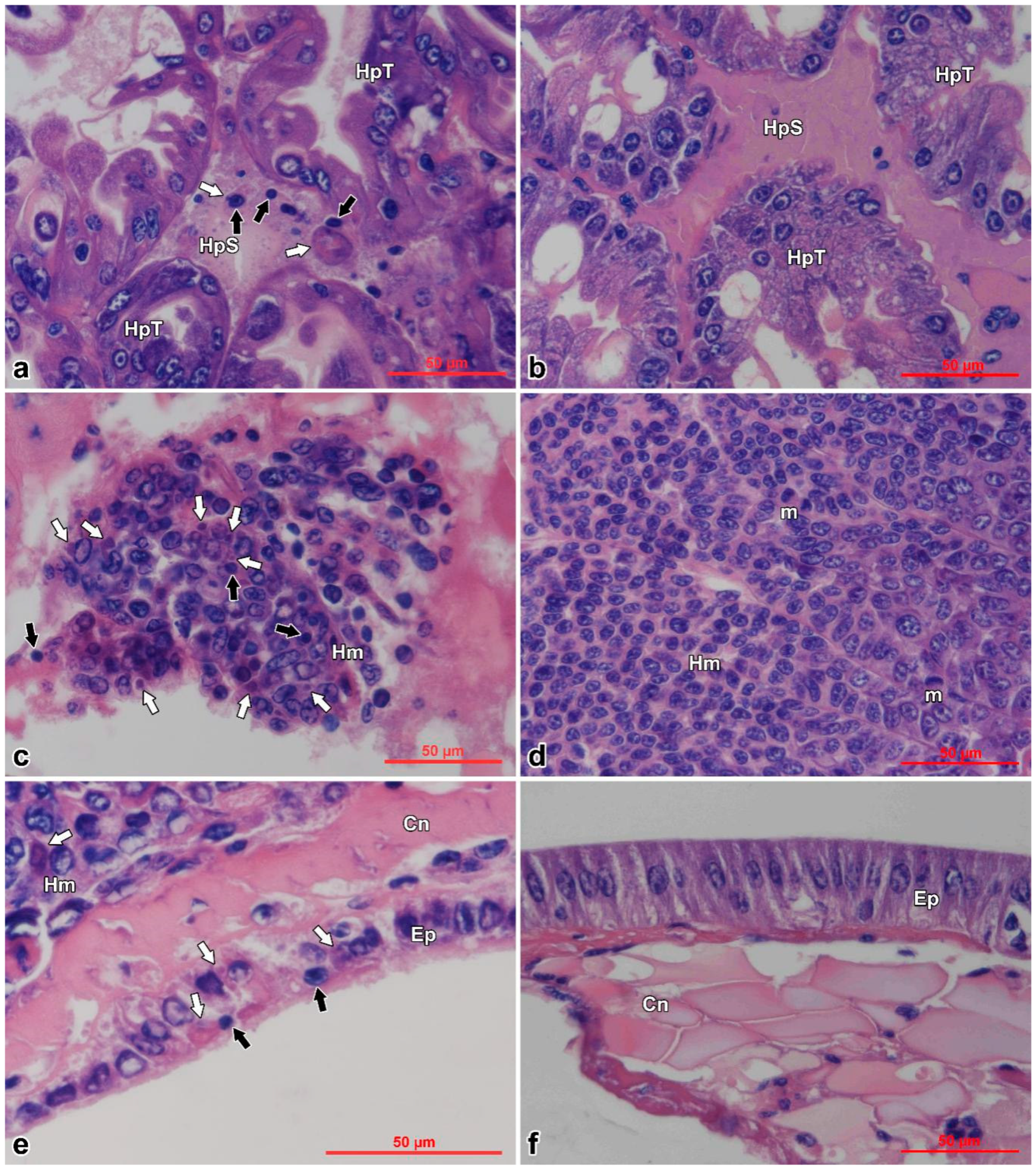
3.3. Histopathology
3.4. Primer Design and LAMP Reaction
3.5. ISDL
3.6. TEM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jun, K.; Kazuhiro, N. Megalocytiviruses. Viruses 2012, 4, 521–538. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Yu, K.H.; Jancovich, J.K. The Molecular biology of Frog virus 3 and other Iridoviruses infecting cold-blooded vertebrates. Viruses 2011, 3, 1959–1985. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.W.; Subramaniam, K.; Waltzek, T.B.; Whittington, R.; Williams, T.; et al. ICTV virus taxonomy profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef] [PubMed]
- Montanie, H.; Bonami, J.R.; Comps, M. Irido-like virus infection in the crab Macropipus depurator L. (Crustacea, Decapoda). J. Invertebr. Pathol. 1993, 61, 320–322. [Google Scholar] [CrossRef]
- Lightner, D.V.; Redman, R.M. A putative iridovirus from the penaeid shrimp Protrachypene precipua Burkenroad (Crustacea Decapoda). J. Invertebr. Pathol. 1993, 1, 107–109. [Google Scholar] [CrossRef]
- Miao, H.Z.; Tong, S.L.; Xu, B.; Jiang, M.; Liu, X.Y. Viral and Pathological observation in cultured lymphoid tissues of shrimp Penaeus chinensis. J. Fish. China 1999, 2, 169–173. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Redman, R.M.; Pantoja, C.R.; Groumellec, M.L.; Duraisamy, P.; Lightner, D.V. Identification of an iridovirus in Acetes erythraeus (Sergestidae) and the development of in situ hybridization and PCR method for its detection. J. Invertebr. Pathol. 2007, 96, 255–260. [Google Scholar] [CrossRef]
- Xu, L.M.; Wang, T.; Li, F.; Yang, F. Isolation and preliminary characterization of a new pathogenic iridovirus from redclaw crayfish Cherax quadricarinatus. Dis. Aquat. Organ. 2016, 120, 17–26. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.M.; Wan, X.Y.; Li, C.; Zhang, Q.L.; Wang, R.Y.; Cheng, D.Y.; Dong, X.; Yang, B.; Wang, X.H.; et al. Characterization of a new member of Iridoviridae, Shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Yang, F.; Huang, J.; Williams, T.; Whittington, R.; Jancovich, J.; Subramaniam, K.; Waltzek, T.; Hick, P.; Ince, I.A.; et al. One new genus with one new species in the subfamily Betairidovirinae. ICTV taxonomic proposal to the Iridoviridae Study Group of International Committee for Taxonomy of Viruses, 2018.004D. ICTV 2018. Available online: https://talk.ictvonline.org/files/ictv_official_taxonomy_updates_since_the_8th_report/m/animal-dna-viruses-and-retroviruses/8051 (accessed on 10 March 2019).
- OIE. Chapter 1.5 Criteria for listing species as susceptible to infection with a specific pathogen. In Aquatic Animal Health Code, 21nd ed.; World Organisation for Animal Health: Paris, France, 2018; ISBN 978-92-95108-69-1. Available online: http://www.oie.int/index.php?id=171&L=0&htmfile=chapitre_criteria_species.htm (accessed on 10 March 2019).
- Xu, W.; Xie, J.; Shi, H.; Li, C. Hematodinium infections in cultured ridgetail white prawns, Exopalaemon carinicauda, in eastern China. Aquaculture 2010, 300, 25–31. [Google Scholar] [CrossRef]
- Wang, X.Q.; Yan, B.L.; Ma, S.; Dong, S.L. Study on the biology and cultural ecology of Exopalaemon carinicauda. Shandong Fish. 2005, 8, 21–23. [Google Scholar]
- OIE. Manual of Diagnostic Tests for Aquatic Animals, 7th Edition. World Organisation for Animal Health. 2016; ISBN 978-92-9044-887-7. Available online: http://www.oie.int/standard-setting/aquatic-manual/access-online/ (accessed on 10 January 2019).
- Li, X.P.; Wan, X.Y.; Xu, T.T.; Huang, J.; Zhang, Q.L. Development and validation of a TaqMan RT-qPCR for the detection of convert mortality nodavirus (CMNV). J. Virol. Methods 2018, 62, 65–71. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.M.; Wan, X.Y.; Zhang, Q.L.; Li, C.; Dong, X.; Yang, B.; Huang, J. Detection and quantification of shrimp hemocyte iridescent virus by TaqMan probe based real-time PCR. J. Invertebr. Pathol. 2018, 154, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Song, X.L.; Yu, J.; Zhang, L.J. The components of an inorganic physiological buffer for Penaeus chinensis. Methods Cell Sci. 1999, 21, 225–230. [Google Scholar] [CrossRef]
- Bell, T.A.; Lightner, D.V. A Handbook of Normal Penaeid Shrimp Histology; World Aquaculture Society: Baton Rouge, LA, USA, 1988; ISBN 0-935868-37-2. [Google Scholar]
- Jitrakorn, S.; Arunrut, N.; Sanguanrut, P.; Flegel, T.W.; Kiatpathomchai, W.; Saksmerprome, V. In situ DIG-labeling, loop-mediated DNA amplification (ISDL) for highly sensitive detection of infectious hypodermal and hematopoietic necrosis virus (IHHNV). Aquaculture 2016, 456, 36–43. [Google Scholar] [CrossRef]
- Lightner, D.V. A Handbook of Shrimp Pathology and Diagnostic Procedures for Disease of Cultured Penaeid Shrimp; World Aquaculture Society: Baton Rouge, LA, USA, 1996; ISBN 0-962452-99-8. [Google Scholar]
- Graham, L.; Orenstein, J.M. Processing tissue and cells for transmission electron microscopy in diagnostic pathology and research. Nat. Protoc. 2007, 2, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Panphut, W.; Senapin, S.; Sriurairatana, S.; Withyachumnarnkul, B.; Flegel, T.W. A novel integrase-containing element may interact with Laem-Singh virus (LSNV) to cause slow growth in giant tiger shrimp. BMC Vet. Res. 2011, 7, 18. [Google Scholar] [CrossRef]
- Qiu, L.; Dong, X.; Wan, X.Y.; Huang, J. Analysis of iridescent viral disease of shrimp (SHID) in 2017. In Analysis of Important Diseases of Aquatic Animals in China in 2017; Fishery and Fishery Administration Bureau under the Ministry of Agriculture and Rural Affairs, National Fishery Technical Extension Center, Ed.; China Agriculture Press: Beijing, China, 2018; pp. 187–204. ISBN 978-7-109-24522-8. (In Chinese) [Google Scholar]
- Qiu, L.; Chen, X.; Zhao, R.H.; Li, C.; Gao, W.; Zhang, Q.L.; Huang, J. Description of a natural infection with decapod iridescent virus 1 in farmed giant freshwater prawn, Macrobrachium rosenbergii. Viruses 2019, 11, 354. [Google Scholar] [CrossRef]
- Kun, P.C.; Fang, Y.H.; Tian, W.T.; Feng, Y.; Ming, C.J. Study of Cherax quadricarinatus iridescent virus in two crabs. J. Appl. Oceanogr. 2017, 1, 82–86. [Google Scholar] [CrossRef]
- Lei, Z.W.; Huang, J.; Shi, C.Y.; Zhang, L.J.; Yu, K.K. Investigation into the hosts of White spot syndrome virus (WSSV). Oceanologia et Limnologia Sinica 2002, 33, 250–258. [Google Scholar]
- Liu, S.; Li, J.T.; Tian, Y.; Wang, C.; Li, X.P.; Xu, T.T.; Li, J.; Zhang, Q.L. Experimental vertical transmission of covert mortality nodavirus in Exopalaemon carinicauda. J. Gen. Virol. 2017, 98, 652–661. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Gui, T.; Sun, Z.; Xiang, J. A copper-induced metallothionein gene from Exopalaemon carinicauda and its response to heavy metal ions. Int J. Biol. Macromol. 2014, 70, 246–250. [Google Scholar] [CrossRef]
- Huang, J.; Song, X.L.; Yu, J.; Yang, C.H. Baculoviral hypodermal and hematopoietic necrosis-study on the pathogen and pathology of the explosive epidemic disease of shrimp. Mar. Fish. Res. 1995, 16, 1–10, (In Chinese with English abstract). [Google Scholar]
- Huang, J.; Zeng, L.B.; Dong, X.; Liang, Y.; Xie, G.S.; Zhang, Q.L. Trend Analysis and Policy Recommendation on Aqutic Biosecurity in China. Eng. Sci. 2016, 18, 15–21, (In Chinese with English abstract). [Google Scholar]
- Huang, J.; Dong, X.; Zhang, Q.L.; Wan, X.Y.; Xie, G.S.; Yang, B.; Wang, H.L.; Yang, Q.; Xu, H.; Wang, X.H. Emerging diseases and biosecurity strategies for shrimp farming. In #We R Aquaculture, AQUA 2018; The World Aquaculture Society: Monpellier, France, 25–29 August 2018; p. 356. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Qiu, L.; Wang, H.; Zou, P.; Dong, X.; Li, F.; Huang, J. Susceptibility of Exopalaemon carinicauda to the Infection with Shrimp Hemocyte Iridescent Virus (SHIV 20141215), a Strain of Decapod Iridescent Virus 1 (DIV1). Viruses 2019, 11, 387. https://doi.org/10.3390/v11040387
Chen X, Qiu L, Wang H, Zou P, Dong X, Li F, Huang J. Susceptibility of Exopalaemon carinicauda to the Infection with Shrimp Hemocyte Iridescent Virus (SHIV 20141215), a Strain of Decapod Iridescent Virus 1 (DIV1). Viruses. 2019; 11(4):387. https://doi.org/10.3390/v11040387
Chicago/Turabian StyleChen, Xing, Liang Qiu, Hailiang Wang, Peizhuo Zou, Xuan Dong, Fuhua Li, and Jie Huang. 2019. "Susceptibility of Exopalaemon carinicauda to the Infection with Shrimp Hemocyte Iridescent Virus (SHIV 20141215), a Strain of Decapod Iridescent Virus 1 (DIV1)" Viruses 11, no. 4: 387. https://doi.org/10.3390/v11040387
APA StyleChen, X., Qiu, L., Wang, H., Zou, P., Dong, X., Li, F., & Huang, J. (2019). Susceptibility of Exopalaemon carinicauda to the Infection with Shrimp Hemocyte Iridescent Virus (SHIV 20141215), a Strain of Decapod Iridescent Virus 1 (DIV1). Viruses, 11(4), 387. https://doi.org/10.3390/v11040387