Metagenomic Next-Generation Sequencing Reveal Presence of a Novel Ungulate Bocaparvovirus in Alpacas

 ,
,
Abstract
:Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Zhao, J.; Zheng, M.; Liu, Z.; Yuan, J.; Zhao, J.; Shen, Q.; Fan, Z.; Jiang, L. Genome Sequence of a Porcine Bocavirus Detected in Feces of Domestic Minks in China. Genome Announc. 2017, 5, e01170-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.K.; Ahmed, S.S.; Yeung, H.C.; Li, K.S.; Fan, R.Y.; Cheng, T.Y.; Cai, J.P.; Wang, M.; Zheng, B.J.; Samson, S.Y.W.; et al. Identification and Interspecies Transmission of a Novel Bocaparvovirus among Different Bat Species in China. J. Gen. Virol. 2016, 97, 3345–3358. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Tsoi, H.-W.; Patteril, N.G.; Yeung, H.C.; Joseph, S.; Wong, E.Y.M.; Muhammed, R.; Chow, F.W.N. Two novel dromedary camel bocaparvoviruses from dromedaries in the Middle East with unique genomic features. J. Gen. Virol. 2017, 98, 1349–1359. [Google Scholar]
- Kapoor, A.; Mehta, N.; Esper, F.; Poljsak-Prijatelj, M.; Quan, P.L.; Qaisar, N.; Delwart, E.; Lipkin, W.L. Identification and characterization of a new Bocavirus species in Gorillas. PLoS ONE 2010, 5, e11948. [Google Scholar] [CrossRef] [PubMed]
- Ao, Y.; Li, X.; Li, L.; Xie, X.; Jin, D.; Yu, J.; Lu, S.; Duan, Z. Two novel bocaparvovirus species identified in wild Himalayan marmots. Sci. China Life Sci. 2017, 60, 1348–1356. [Google Scholar] [CrossRef]
- Gunn, L.; Collins, P.J.; Fanning, S.; McKillen, J.; Morgan, J.; Staines, A.; O’Shea, H. Detection and characterisation of novel bocavirus (genus Bocaparvovirus) and gastroenteritis viruses from asymptomatic pigs in Ireland. Infect. Ecol. Epidemiol. 2015, 5, 27270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Song, F.; Xiu, L.; Liu, Y.; Yang, J.; Yao, L.; Peng, J. Identification and characterization of a novel rodent bocavirus from different rodent species in China. Emerg. Microbes Infect. 2018, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef]
- Cheng, W.-X.; Li, J.-S.; Huang, C.-P.; Yao, D.-P.; Liu, N.; Cui, S.-X.; Jin, Y.; Duan, Z.-J. Identification and nearly full-lengthg characterization of novel porcine Bocaviruses. PLoS ONE 2010, 5, e13583. [Google Scholar] [CrossRef]
- Principi, N.; Piralla, A.; Zampiero, A.; Bianchini, S.; Umbrello, G.; Scala, A.; Bosis, S.; Fossali, E.; Baldanti, F. Bocavirus infection in otherwise healthy children with respiratory disease. PLoS ONE 2015, 10, e0135640. [Google Scholar] [CrossRef]
- Kailasan, S.; Halder, S.; Gurda, B.; Bladek, H.; Chipman, P.R.; McKenna, R.; Brown, K.; Agbandje-McKenna, M. Structure of an enteric pathogen, bovine parvovirus. J. Virol. 2015, 89, 2603–2614. [Google Scholar] [CrossRef]
- Binn, L.N.; Lazar, E.C.; Eddy, G.A.; Kajima, M. Recovery and characterization of a minute virus of canines. Infect. Immun. 1970, 1, 503–508. [Google Scholar]
- McClenahan, S.D.; Scherba, G.; Borst, L.; Fredrickson, R.L.; Krause, P.R.; Uhlenhaut, C. Discovery of a Bovine Enterovirus in Alpaca. PLoS ONE 2013, 8, e68777. [Google Scholar] [CrossRef]
- Alpaca Owners Association Inc Homepage. Available online: https://www.alpacainfo.com/about/statistics/alpacas-us (accessed on 22 May 2019).
- Crossley, B.M.; Mock, R.E.; Callison, S.A.; Hietala, S.K. Identification and characterization of a novel alpaca respiratory coronavirus most closely related to the human coronavirus 229E. Viruses 2012, 4, 3689–3700. [Google Scholar] [CrossRef]
- Brito, B.P.; Gardner, I.A.; Hietala, S.K.; Crossley, B.M. Variation in Bluetongue virus real-time reverse transcription polymerase chain reaction assay results in blood samples of sheep, cattle, and alpaca. J. Vet. Diagn. Investig. 2011, 23, 753–757. [Google Scholar] [CrossRef]
- Kapil, S.; Yeary, T.; Evermann, J.F. Viral diseases of new world camelids. Vet. Clin. N. Am. Food Anim. Pract. 2009, 25, 323–337. [Google Scholar] [CrossRef]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Barrington, G.M.; Allen, A.J.; Parish, S.M.; Tibary, A. Biosecurity and biocontainment in alpaca operations. Small Rum. Res. 2006, 61, 217–225. [Google Scholar] [CrossRef]
- Wernery, U.; Kaaden, O.R. Foot-and-mouth disease in camelids: A review. Vet. J. 2004, 168, 134–142. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Anderson, J.; Hesse, R.A.; Anderson, G. Bovine rhinitis viruses are common in U.S. cattle with bovine respiratory disease. PLoS ONE 2015, 10, e0121998. [Google Scholar] [CrossRef]
- Neill, J.D.; Bayles, D.O.; Ridpath, J.F. Simultaneous rapid sequencing of multiple RNA virus genomes. J. Virol. Methods 2014, 201, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Knutson, T.P.; Velayudhan, B.T.; Marthaler, D.G. A porcine enterovirus G associated with enteric disease contains a novel papain-like cysteine protease. J. Gen. Virol. 2017, 98, 1305–1310. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014. [Google Scholar] [CrossRef]
- Joshi, N.; Fass, J. Sickle—A Windowed Adaptive Trimming Tool for FASTQ Files Using Quality. 2011. Available online: http://github.com/najoshi/sickle (accessed on 2 July 2019).
- Buffalo, V. Scythe—A Bayesian Adapter Trimmer. 2011. Available online: http://github.com/vsbuffalo/scythe (accessed on 2 July 2019).
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Shackelton, L.A.; Parrish, C.R.; Truyen, U.; Holmes, E.C. High rate of viral evolution associated with the emergence of carnivore parvovirus. Proc. Natl. Acad. Sci. USA 2005, 102, 379–384. [Google Scholar] [CrossRef]
- Fasina, O.O.; Dong, Y.; Pintel, D.J. NP1 protein of the bocaparvovirus minute virus of canines controls access to the viral capsid genes via its role in RNA processing. J. Virol. 2015, 90, 1718–1728. [Google Scholar] [CrossRef]
- Zádori, Z.; Szelei, J.; Lacoste, M.-C.; Li, Y.; Gariépy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A Viral Phospholipase A2 Is Required for Parvovirus Infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.F.; Dong, Y.M.; Yi, Q.H.; Huang, Y.; Zhao, D.; Yang, Y.B.; Tijssen, P.; Qiu, J.M.; Liu, K.Y.; Li, Y. The determinants for the enzyme activity of human parvovirus b19 phospholipase a2 (pla2) and its influence on cultured cells. PLoS ONE 2013, 8, e61440. [Google Scholar] [CrossRef]
- Castrignano, S.B.; Nagasse-Sugahara, T.K. The metagenomic approach and causality in virology. Rev. Saúde Pública. 2015, 49, 21. [Google Scholar] [CrossRef]





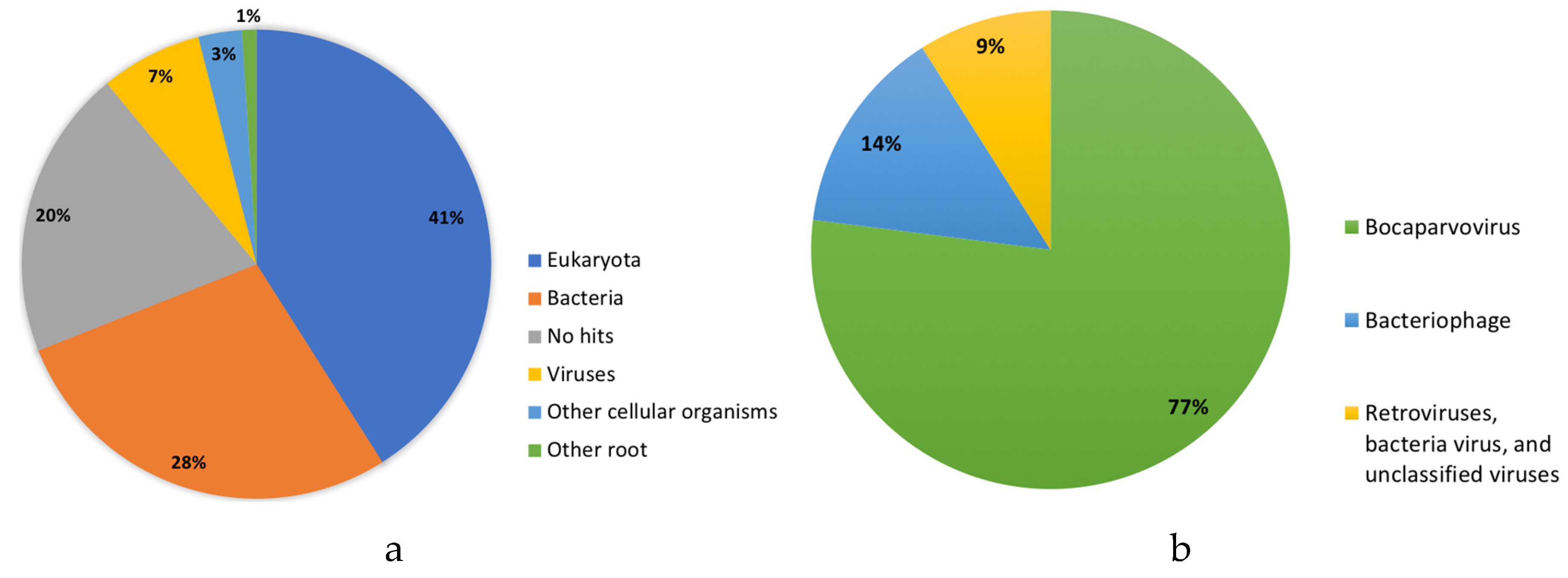{kind=link}
{kind=link}
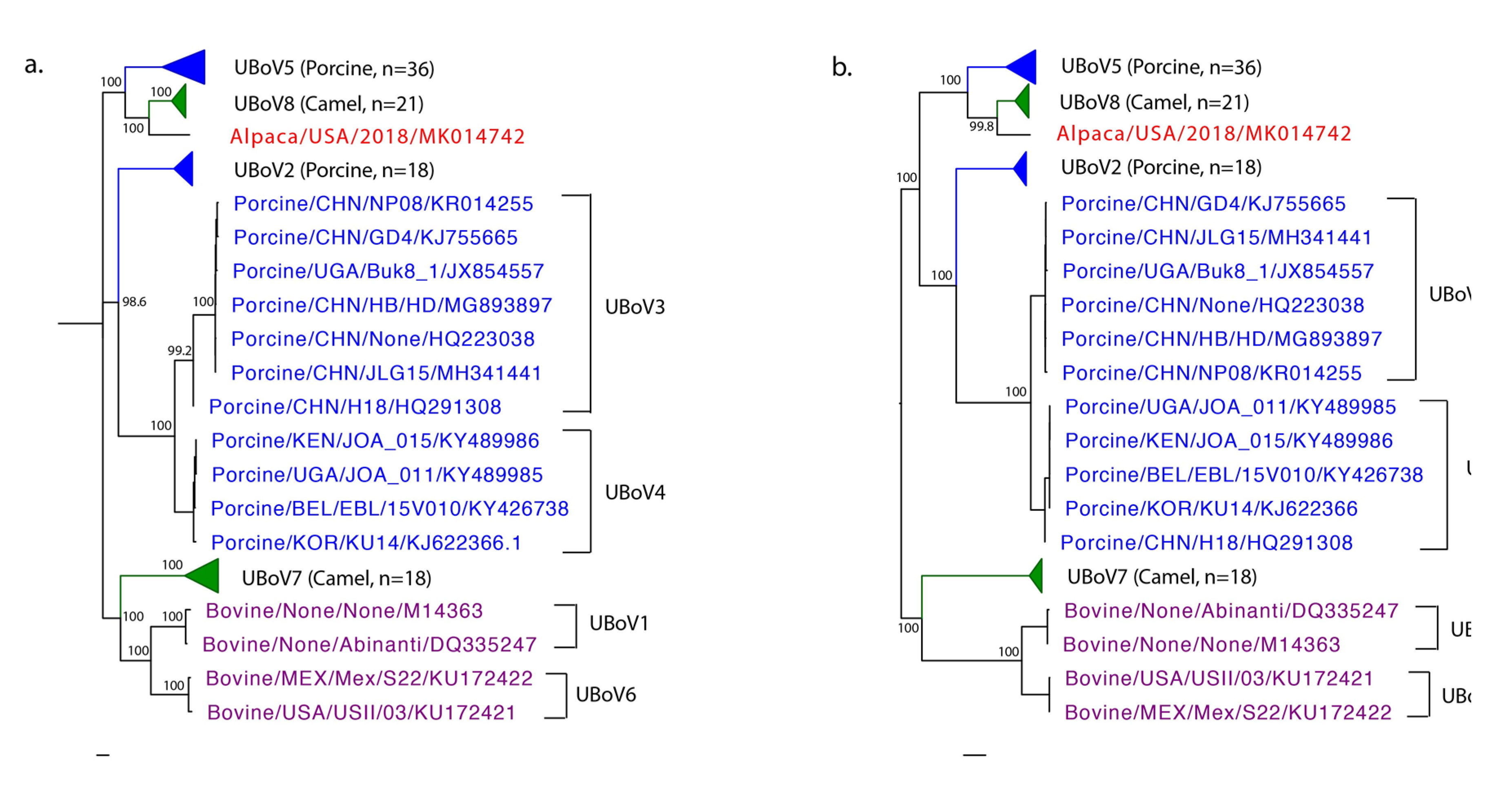{kind=link}
{kind=link}
| UBoV Virus | Species | Whole Genome (nt) | NS1 | NP1 | VP1 | |||
|---|---|---|---|---|---|---|---|---|
| nt | aa | nt | aa | nt | aa | |||
| UBoV1 (n = 2) | Bovine | 36.22–38.98 | 33.32–37.70 | 26.75–30.08 | 35.14–35.42 | 40.44–42.62 | 36.51–36.72 | 28.49–31.45 |
| UBoV2 (n = 18) | Porcine | 37.20–39.61 | 43.40–49.64 | 36.18–40.34 | 34.58–36.94 | 31.89–37.84 | 33.42–34.79 | 27.03–28.65 |
| UBoV3 (n = 6) | Porcine | 33.73–36.17 | 45.64–45.83 | 36.23–36.61 | 34.13–34.49 | 35.96 | 25.96–28.63 | 20.10–20.83 |
| UBoV4 (n = 5) | Porcine | 35.74–36.07 | 45.66–45.95 | 37.06–37.66 | 34.89–35.61 | 31.52–31.87 | 27.20–27.64 | 21.31–21.59 |
| UBoV5 (n = 36) | Porcine | 44.37–47.32 | 52.83–59.39 | 46.58–52.8 | 37.59–40.40 | 34.78–40.53 | 39.02–40.54 | 30.76–35.81 |
| UBoV6 (n = 2) | Bovine | 38.58–38.62 | 37.60–38.01 | 29.64 | 33.02 | 40.56–40.88 | 36.17–36.32 | 29.45–29.88 |
| UBoV7 (n = 18) | Camel | 36.49–36.76 | 39.01–39.64 | 33.08–33.98 | 36.91–38.93 | 32.24–33.88 | 26.05–27.11 | 21.08–21.84 |
| UBoV8 (n = 21) | Camel | 57.77–58.58 | 67.64–69.26 | 57.89–67.85 | 46.99–52.47 | 53.51–56.76 | 45.05–46.36 | 42.15–43.60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, D.; Chaudhary, S.; Lu, N.; Duff, M.; Heffel, M.; McKinney, C.A.; Bedenice, D.; Marthaler, D. Metagenomic Next-Generation Sequencing Reveal Presence of a Novel Ungulate Bocaparvovirus in Alpacas. Viruses 2019, 11, 701. https://doi.org/10.3390/v11080701
Kumar D, Chaudhary S, Lu N, Duff M, Heffel M, McKinney CA, Bedenice D, Marthaler D. Metagenomic Next-Generation Sequencing Reveal Presence of a Novel Ungulate Bocaparvovirus in Alpacas. Viruses. 2019; 11(8):701. https://doi.org/10.3390/v11080701
Chicago/Turabian StyleKumar, Deepak, Suman Chaudhary, Nanyan Lu, Michael Duff, Mathew Heffel, Caroline A. McKinney, Daniela Bedenice, and Douglas Marthaler. 2019. "Metagenomic Next-Generation Sequencing Reveal Presence of a Novel Ungulate Bocaparvovirus in Alpacas" Viruses 11, no. 8: 701. https://doi.org/10.3390/v11080701