The Expression of Potato Expansin A3 (StEXPA3) and Extensin4 (StEXT4) Genes with Distribution of StEXPAs and HRGPs-Extensin Changes as an Effect of Cell Wall Rebuilding in Two Types of PVYNTN–Solanum tuberosum Interactions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Virus Inoculation
2.2. Isolation of RNA and Genomic DNA (gDNA) for Analysis of the Expression of StEXPA3 and StEXT4 in Potato Plants with Differing Resistances Against PVY
2.3. Real-Time Quantitative Polymerase Chain Reaction (qPCR) Expression Analysis of StEXPA3 and StEXT4 in Potato Plants with Different Resistances to PVYNTN
2.4. Immunofluorescence Localization and Assessment of the Quantitative Fluorescence Signal Using the Corrected Total Cell Fluorescence Method (CTCF) in Potato Plants with Different Resistances for PVY
2.5. Electron Microscopy Material Preparation, Immunogold Localization, and Statistical Quantification of the Cell Distribution of EXPAs and Extensins in Potato Plants with Different Resistances for PVY
3. Results
3.1. Expression Analysis of Expansin A3 (StEXPA3) in Compatible and Incompatible PVYNTN–Potato Interactions
3.2. Immunofluorescence Expansins Localization and Assessment of the Quantitative Fluorescence Signal in PVYNTN–Potato Compatible and Incompatible Interactions
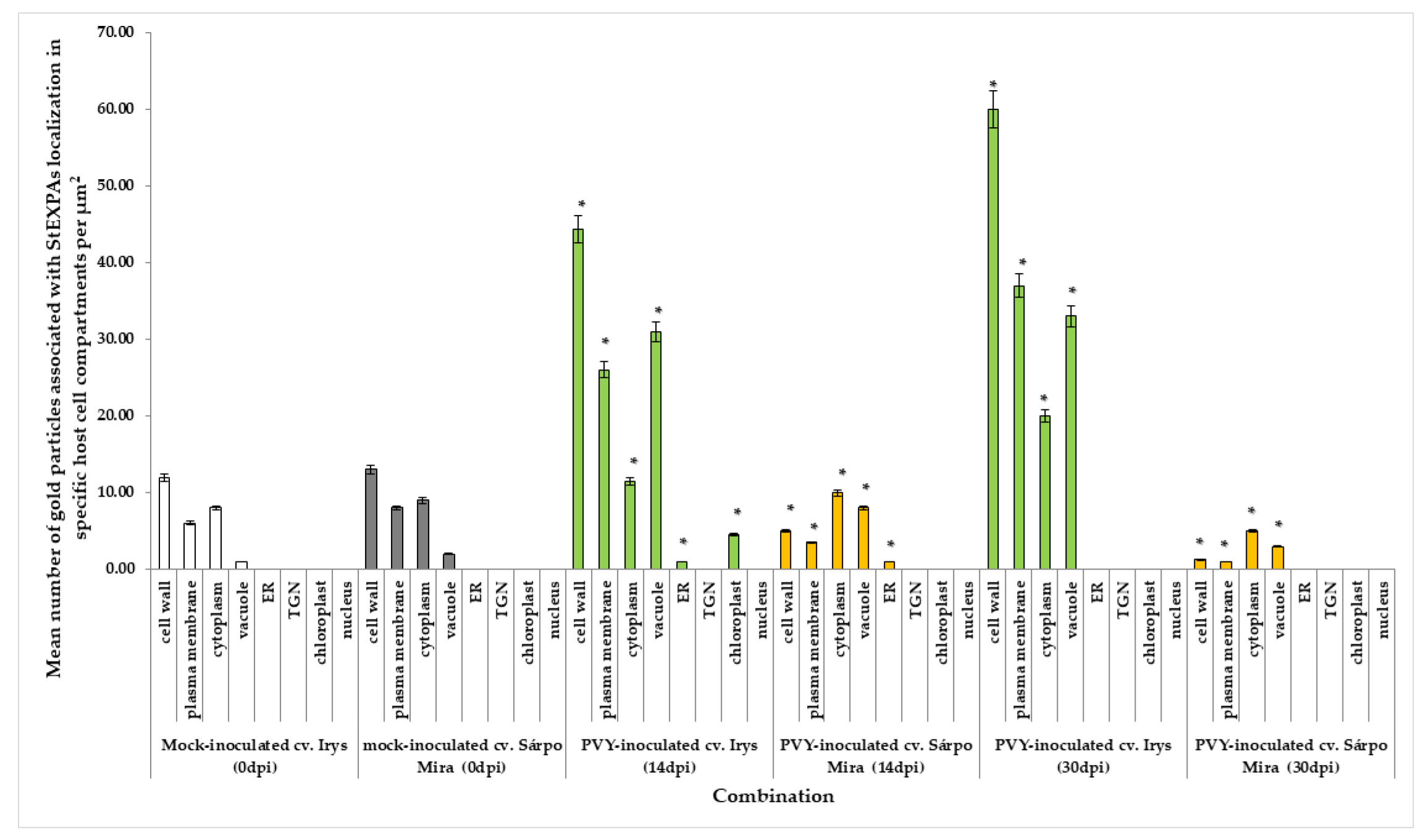
3.3. Quantitative Immunogold Localization of StEXPAs in Compatible and Incompatible PVYNTN–Potato Interactions
3.4. Expression Analysis of Extensin 4 (StEXT4) in Compatible and Incompatible PVYNTN–Potato Interactions
3.5. Immunofluorescence Localization of Hydroxyproline-Rich Glycoprotein (HRGP) Extensins with the Assessment of the Quantitative Fluorescence Signal in Compatible and Incompatible PVYNTN–Potato Interactions
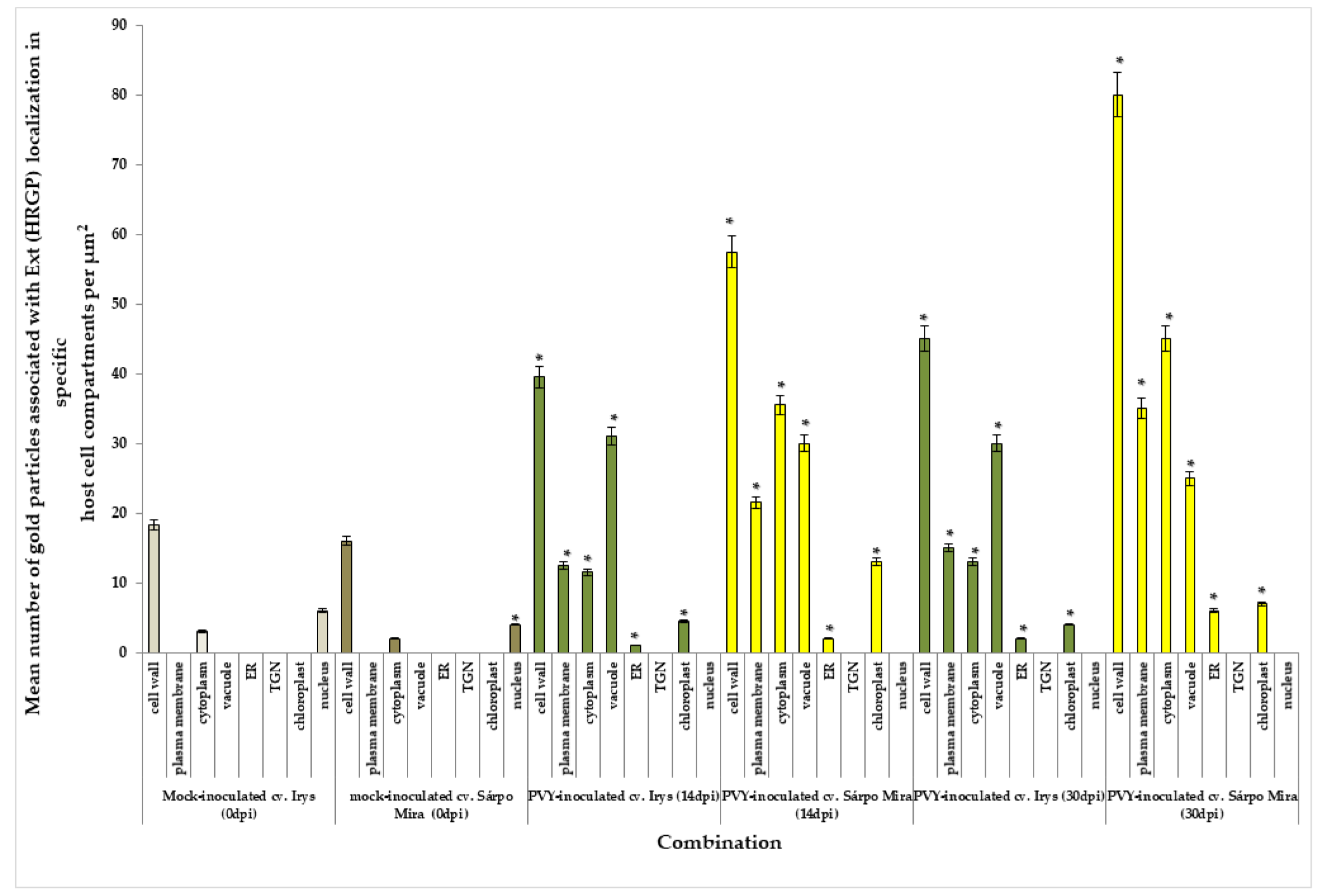
3.6. Immunogold Localization of Extensins (HGRPs) with Quantification in Compatible and Incompatible PVYNTN–Potato Interactions
4. Discussion
4.1. StEXPA3 Gene Expression and StEXPAs Localization in PVYNTN–Potato Interactions
4.2. StEXT4 Gene Expression and Hydroxyproline-Rich Glycoprotein Extensins (HRGPs) Localization in PVYNTN–Potato Interactions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 945–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, R.; Shinohara, N.; Asaoka, R.; Narukawa, H.; Nishitani, K. The biosynthesis and function of polysaccharide components of the plant cell wall. In Plant Cell Wall Patterning and Cell Shape, 1st ed.; Fukuda, H., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2014; Volume 1, pp. 3–34. [Google Scholar]
- Tucker, M.R.; Koltunow, A.M. Traffic monitors at the cell periphery: The role of cell walls during early female reproductive cell differentiation in plants. Curr. Opin. Plant Biol. 2014, 17, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Houston, K.; Tucker, M.R.; Chowdhury, J.; Shirley, N.; Little, A. The plant cell wall: A complex and dynamic structure as revealed by the responses of genes under stress conditions. Front. Plant Sci. 2016, 7, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Loosening of plant cell walls by expansions. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Rayle, D.L.; Cleland, R.E. The acid growth theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef] [Green Version]
- McQueen-Mason, S.J.; Cosgrove, D.J. Expansin mode of action on cell walls. Analysis of wall hydrolysis, stress relaxation, and binding. Plant Physiol. 1995, 107, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Kende, H.; Bradford, K.; Brummell, D.; Cho, H.T.; Cosgrove, D.J.; Fleming, A.; Gehring, C.; Lee, Y.; McQueen-Mason, S.J.; Rose, J.; et al. Nomenclature for members of the expansin superfamily of genes and proteins. Plant Mol. Biol. 2004, 55, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Choi, D.; Kende, H. Expansins: Everexpanding numbers and functions. Curr. Opin. Plant Biol. 2001, 4, 527–532. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.; Zhang, C.; Jiang, X.; Kang, M.; Yin, X.; Lu, P.; Zhang, X.; Zheng, Y.; Gao, J. RhNAC2 and RhEXPA4 are involved in the regulation of dehydration tolerance during the expansion of rose petals. Plant Physiol. 2012, 160, 2064–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 15, 1254–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Xu, X.; Shi, Y.; Xu, J.; Huang, B. Transgenic tobacco plants overexpressing a grass PpEXP1 gene exhibit enhanced tolerance to heat stress. PLoS ONE 2014, 9, e100792. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Li, F.; Renaud, J.; Shen, W.; Li, Y.; Guo, L.; Cui, H.; Sumarah, M.; Wang, A. NbEXPA1, an α-expansin, is plasmodesmata-specific and a novel host factor for potyviral infection. Plant J. 2017, 92, 846–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuqamar, S.; Ajeb, S.; Sham, A.; Enan, M.R.; Iratni, R. A mutation in the expansin-like A2 gene enhances resistance to necrotrophic fungi and hypersensitivity to abiotic stress in Arabidopsis thaliana. Mol. Plant Pathol. 2013, 14, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Zou, W.S.; Wu, G.; Lin, H.H.; Xi, D.H. Tobacco alpha-expansin EXPA4 plays a role in Nicotiana benthamiana defence against Tobacco mosaic virus. Planta 2018, 247, 355–368. [Google Scholar] [CrossRef]
- Chen, L.J.; Zou, W.S.; Fei, C.Y.; Wu, G.; Li, X.Y.; Lin, H.H.; Xi, D.H. α-Expansin EXPA4 positively regulates abiotic stress tolerance but negatively regulates pathogen resistance in Nicotiana tabacum. Plant Cell Physiol. 2018, 59, 2317–2330. [Google Scholar] [CrossRef]
- Wieczorek, K.; Golecki, B.; Gerdes, L.; Heinen, P.; Szakasits, D.; Durachko, D.M.; Cosgrove, D.J.; Kreil, D.P.; Puzio, P.S.; Bohlmann, H.; et al. Expansins are involved in the formation of nematode-induced syncytia in roots of Arabidopsis thaliana. Plant J. 2006, 48, 98–112. [Google Scholar] [CrossRef]
- Ding, X.; Cao, Y.; Huang, L.; Zhao, J.; Xu, C.; Li, X.; Wang, S. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Deepak, S.; Shailasree, S.; Kini, R.K.; Muck, A.; Mithofer, A.; Shetty, S.H. Hydroxyproline-rich glycoproteins and plant defense. J. Phytopathol. 2010, 158, 585–593. [Google Scholar]
- Velasquez, S.M.; Salgado-Salter, J.; Petersen, B.L.; Estevez, J.M. Recent advances on the post-translational modifications of EXTs and their roles in plant cell walls. Front. Plant Sci. 2012, 3, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, P.M.; Brownleader, M.D.; Pantelides, A.T.; Trevan, M.; Smith, J.J.; Saddler, G. Extensin from suspension-cultured potato cells: A hydroxyproline-rich glycoprotein, devoid of agglutinin activity. Planta 1997, 202, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Raggi, V. Hydroxyproline-rich glycoprotein accumulation in tobacco leaves protected against Erysiphe cichoracearum by potato virus Y infection. Plant Pathol. 2000, 49, 179–186. [Google Scholar] [CrossRef]
- Wei, G.; Shirsat, A.H. Extensin over-expression in Arabidopsis limits pathogen invasiveness. Mol. Plant Pathol. 2006, 7, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Wycoff, K.L.; Powell, P.A.; Gonzales, R.A.; Corbin, D.R.; Lamb, C.; Dixon, R.A. Stress activation of a bean hydroxyproline-rich glycoprotein promoter is superimposed on a pattern of tissue-specific developmental expression. Plant Physiol. 1995, 109, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otulak-Kozieł, K.; Kozieł, E.; Lockhart, B.E.L. Plant cell wall dynamics in compatible and incompatible potato response to infection caused by Potato virus Y (PVYNTN). Int. J. Mol. Sci. 2018, 19, 862. [Google Scholar] [CrossRef] [Green Version]
- Otulak-Kozieł, K.; Kozieł, E.; Bujarski, J.J. Spatiotemporal changes in xylan-1/xyloglucan and xyloglucan xyloglucosyl transferase (Xth-Xet5) as a step-in of ultrastructural cell wall remodelling in potato–Potato virus y (PVYntn) hypersensitive and susceptible reaction. Int. J. Mol. Sci. 2018, 19, 2287. [Google Scholar] [CrossRef] [Green Version]
- Zimmoch-Guzowska, E.; Yin, Z.; Chrzanowska, M.; Flis, B. Sources and effectiveness of potato PVY resistance in IHAR’s breeding research. Am. Potato J. 2013, 90, 21–27. [Google Scholar] [CrossRef] [Green Version]
- The European Cultivated Potato Database. Available online: https://www.europotato.org/quick_search.php (accessed on 19 October 2019).
- Otulak, K.; Kozieł, E.; Garbaczewska, G. Ultastructural impact of tobacco rattle virus on tobacco and pepper ovary and anther tissues. J. Phytopatol. 2016, 164, 226–241. [Google Scholar] [CrossRef]
- Otulak, K.; Garbaczewska, G. Ultrastructural events during hypersensitive response of potato cv. Rywal infected with necrotic strains of Potato virus Y. Acta Physiol. Plant. 2010, 32, 635–644. [Google Scholar] [CrossRef]
- Chrzanowska, M.; Doroszewska, T. Comparison between PVY isolates obtained from potato and tobacco plants grown in Poland. Phytopathol. Pol. 1997, 13, 63–71. [Google Scholar]
- Tomczyńska, I.; Jupe, F.; Hein, I.; Marczewski, W.; Śliwka, J. Hypersensitive response to Potato virus Y in potato cultivar Sárpo Mira is conferred by the Ny-Smira gene located on the long arm of chromosome IX. Mol. Breed. 2014, 34, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zhang, N.; Si, H.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information GeneBank Database with Sequence of Expansin-A3. Available online: https://www.ncbi.nlm.nih.gov/nuccore/XM_006340959.2/ (accessed on 19 October 2019).
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information GeneBank Database with Sequence of Extensin 4. Available online: https://www.ncbi.nlm.nih.gov/nuccore/971585705 (accessed on 19 October 2019).
- Gubler, F. Immunofluorescence localisation of microtubules in plant root tips embedded in butyl-methyl methacrylate. Cell Biol. Int. Rep. 1989, 13, 137–145. [Google Scholar] [CrossRef]
- The University of Sydney Official Website of the Bosh Institute. Available online: https://sydney.edu.au/medicine/bosch/facilities/advanced-microscopy/user-support/ImageJ_FL_Image_Analysis.pdf (accessed on 6 October 2019).
- Burgess, A.; Vigneron, S.; Brioudes, E.; Labbé, J.C.; Lorca, T.; Castro, A. Loss of human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of the cyclin B-Cdc2/PP2A balance. Proc. Natl. Acad. Sci. USA 2010, 107, 12564–12569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otulak-Kozieł, K.; Kozieł, E.; Valverde, R.A. The respiratory burst oxidase homolog d (rbohd) cell and tissue distribution in potato–potato virus y (pvyntn) hypersensitive and susceptible reactions. Int. J. Mol. Sci. 2019, 20, 2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otulak, K.; Garbaczewska, G. The participation of plant cell organelles in compatible and incompatible Potato virus Y-tobacco and -potato plant interaction. Acta Physiol. Plant. 2013, 36, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Luschin-Ebengreuth, N.; Zechmann, B. Compartment-specific investigations of antioxidants and hydrogen peroxide in leaves of Arabidopsis thaliana during dark-induced senescence. Acta Physiol. Plant. 2016, 38, 133. [Google Scholar] [CrossRef] [Green Version]
- Underwood, W. The plant cell wall: A dynamic barrier against pathogen invasion. Front. Plant Sci. 2012, 3, 85. [Google Scholar] [CrossRef] [Green Version]
- Büttner, D.; He, S.Y. Type III protein secretion in plant pathogenic bacteria. Plant Physiol. 2009, 150, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Plant cell wall extensibility: Connecting plant cell growth with cell wall structure, mechanics, and the action of wall-modifying enzymes. J. Exp. Bot. 2015, 67, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Han, Y.Y.; Chen, Y.; Kong, X.; Wang, W. The involvement of expansins in response to water stress during leaf development in wheat. J. Plant Physiol. 2015, 183, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Pien, S.; Wyrzykowska, J.; McQueen-Mason, S.; Smart, C.; Fleming, A. Local expression of expansin induces the entire process of leaf development and modifies leaf shape. Proc. Natl. Acad. Sci. USA 2001, 98, 11812–11817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.T.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuluev, B.R.; Knyazev, A.V.; Lebedev, Y.P.; Chemaris, A.V. Morphological and physiological characteristics of transgenic tobacco plants expressing expansin genes: AtEXP10 from Arabidopsis and PnEXPA1 from poplar. Russ. J. Plant Physiol. 2012, 59, 97–104. [Google Scholar] [CrossRef]
- Kuluev, B.R.; Safiullina, M.G.; Knyazev, A.V.; Chemaris, A.V. Effect of ectopic expression of NtEXPA5 gene on cell size and growth of organs of transgenic tobacco plants. Russ. J. Plant Physiol. 2013, 44, 28–34. [Google Scholar] [CrossRef]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Guo, R.; Jie, F.; Nettleton, D.; Peng, J.; Carr, T.; Yeakley, J.M.; Fan, J.B.; Whitham, S.A. Spatial Analysis of Arabidopsis thaliana Gene Expression in Response to Turnip mosaic virus Infection. Mol. Plant Microbe Interact. 2007, 20, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Showalter, A.M.; Bell, J.N.; Cramer, C.L.; Bailey, J.A.; Varner, J.E.; Lamb, C.J. Accumulation of hydroxyproline-rich glycoprotein mRNAs in response to fungal elicitor and infection. Proc. Natl. Acad. Sci. USA 1985, 82, 6551–6555. [Google Scholar] [CrossRef] [Green Version]
- Shetty, N.P.; Jensen, J.D.; Knudsen, A.; Finnie, C.; Geshi, N.; Blennow, A.; Collinge, D.B.; Jørgensen, H.J. Effects of beta-1,3-glucan from Septoria tritici on structural defense responses in wheat. J. Exp. Bot. 2009, 60, 4287–4300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Satoh, K.; Kikuchi, S.; Omura, T. The repression of cell wall and plastid-related genes and the induction of defense-related genes in rice plants infected with Rice dwarf virus. Mol. Plant-Microbe Interact. 2007, 20, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Ma, L.; Zhao, J.; Li, Z.; Sun, F.; Lu, X. Comparative Transcriptome Analysis of Two Rice Varieties in Response to Rice Stripe Virus and Small Brown Planthoppers during Early Interaction. PLoS ONE 2013, 8, e82126. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, N.; Mazau, D.; Esquerré-Tugayé, M.-T.; Asselin, A. Immunogold localization of hydroxyproline-rich glycoproteins in necrotic tissue of Nicotiuna tabucum L. cv. Xunthi-nc infected by tobacco mosaic virus. Physiol. Mol. Plant Pathol. 1990, 36, 129–145. [Google Scholar] [CrossRef]
- Chakraborty, N.; Basak, J. Comparative transcriptome profiling of a resistant vs. susceptible Vigna mungo cultivar in response to Mungbean yellow mosaic India virus infection reveals new insight into MYMIV resistance. Curr. Plant Biol. 2018, 15, 8–24. [Google Scholar] [CrossRef]
- Xie, D.; Ma, L.; Samaj, J.; Xu, C. Immunohistochemical analysis of cell wall hydroxyproline-rich glycoproteins in the roots of resistant and susceptible wax gourd cultivars in response to Fusarium oxysporum f. sp. Benincasae infection and fusaric acid treatment. Plant Cell Rep. 2011, 30, 1555–1569. [Google Scholar] [CrossRef]
- Basavaraju, P.; Shailasree, S.; Shetty, N.P.; Ramachandra, K.K.; Jorgensen, H.J.L.; de Neergaard, E.; Shetty, H.S. Infection induced oxidative cross-linking of hydroxyproline-rich glycoproteins (HRGPs) is associated with restriction of Colletotrichum sublineolum in sorghum. J. Plant Interact. 2009, 4, 179–186. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Brown, I.R.; Mansfield, J.W.; Bailey, J.A.; Mazau, D.; Rumeau, D.; Esquerré-Tugayé, M.T. Immunocytochemical localization of hydroxyproline-rich glycoproteins accumulating in melon and bean at sites of resistance to bacteria and fungi. Mol. Plant-Microbe Interact. 1990, 3, 33–40. [Google Scholar] [CrossRef]
- Jeffree, C.E.; Yeoman, M.M. A study of the intracellular and intercellular distribution of the Datura stramonium lectin using an immunofluorescent technique. New Phytol. 1981, 87, 463–471. [Google Scholar] [CrossRef]
- Jose-Estanyol, M.; Puigdomènech, P. Cellular localization of the embryo-specific hybrid PRP from Zea mays, and characterization of promoter regulatory elements of its gene. Plant Mol. Biol. 2012, 80, 325–335. [Google Scholar] [CrossRef]
- Carpita, N.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.J.; Swords, K.M.; Lynch, M.A.; Staehelin, L.A. Spatial organization of the assembly pathways of glycoproteins and complex polysaccharides in the Golgi apparatus of plants. J. Cell Biol. 1991, 112, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fan, W.; Li, X.; Chen, H.; Takáč, T.; Šamajová, O.; Fabrice, M.R.; Xie, L.; Ma, J.; Šamaj, J.; et al. Expression and distribution of extensins and AGPs in susceptible and resistant banana cultivars in response to wounding and Fusarium oxysporum. Sci. Rep. 2017, 20, 42400. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, M.; Beven, A.; Donovan, N.; Neill, S.J.; Peart, J.; Roberts, K.; Knox, J.P. Localization of cell wall proteins in relation to the developmental anatomy of the carrot root apex. Plant J. 1994, 5, 237–246. [Google Scholar] [CrossRef]
- Kovalenko, A.G.; Kovalenko, E.A.; Telegeeva, T.A. Localization of Viral Infection and hemagglutination Activity in Plants. Mikrobiol. Zhurnal 1991, 53, 69–76. [Google Scholar]
- Kovalenko, O.H.; Kyrychenko, A.M.; Teleheieva, T.A. Effect of TMV-infection on protein and carbohydrate content in hypersensitive tobacco plants and their antiviral and hemagglutinating activity. Ukr. Biokhim Zhurnal 2003, 75, 103–108. [Google Scholar]
- Babosha, A.V. Inducible lectins and plant resistance to pathogens and abiotic stress. Biochemistry 2008, 73, 812–825. [Google Scholar] [CrossRef]
- Ponstein, A.S.; Bres-Vloemans, S.A.; Sela-Buurlage, M.B.; van den Elzen, P.J.M.; Melchers, L.S.; Cornelissen, B.J.C. A novel pathogen- and wound-inducible tobacco (Nicotiana tabacum) protein with antifungal activity. Plant Physiol. 1994, 104, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Babosha, A.V.; Ladygina, M.E. Determination of phytohemagglutinins as related to virus resistance of potato. In Physiological-Biochemical and Biophysical Methods of Diagnostics of Plant Resistance to Pathogens and Other Factors, 1st ed.; Ladygina, M.E., Ed.; Moscow State University Publishing House: Moscow, Russia, 1992; Volume 1, pp. 43–52. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene ID | Forward Primer | Reverse Primer | Temperature of Primer Annealing (°C) | Concentration in Reaction (μM) | Product Length (bp) |
|---|---|---|---|---|---|---|
| Investigated | ||||||
| StEXPA3 | XM_006340959.2 | 5′-TGCCGTCAATGCCAGAATCC-3′ | 3′-CACCGTAGAACGTAGCGTGG-5′ | 58 | 0.5 | 74 |
| StEXT4 | XM_006366812.2 | 5′-GATAAGCCATTAGACGCCATT-3′ | 3′-TCGCCAGAACTATCACAGAA-5′ | 58 | 0.5 | 100 |
| Reference | ||||||
| EF1a | AB061263 | 5′-GGTGATGCTGGTATGGTTAAG-3′ | 3′-GGTCCTTCTTGTCAACATTCTT-5′ | 58 | 0.5 | 148 |
| Program | Parameters |
|---|---|
| Preliminary denaturation | 95 °C for 5 min |
| Amplification (35 cycles) | 95 °C for10 s 58 °C for 10 s 72 °C for 20 s * |
| Melting curve | 65–95 °C; 0.5 °C/s |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otulak-Kozieł, K.; Kozieł, E.; Lockhart, B.E.L.; Bujarski, J.J. The Expression of Potato Expansin A3 (StEXPA3) and Extensin4 (StEXT4) Genes with Distribution of StEXPAs and HRGPs-Extensin Changes as an Effect of Cell Wall Rebuilding in Two Types of PVYNTN–Solanum tuberosum Interactions. Viruses 2020, 12, 66. https://doi.org/10.3390/v12010066
Otulak-Kozieł K, Kozieł E, Lockhart BEL, Bujarski JJ. The Expression of Potato Expansin A3 (StEXPA3) and Extensin4 (StEXT4) Genes with Distribution of StEXPAs and HRGPs-Extensin Changes as an Effect of Cell Wall Rebuilding in Two Types of PVYNTN–Solanum tuberosum Interactions. Viruses. 2020; 12(1):66. https://doi.org/10.3390/v12010066
Chicago/Turabian StyleOtulak-Kozieł, Katarzyna, Edmund Kozieł, Benham E. L. Lockhart, and Józef J. Bujarski. 2020. "The Expression of Potato Expansin A3 (StEXPA3) and Extensin4 (StEXT4) Genes with Distribution of StEXPAs and HRGPs-Extensin Changes as an Effect of Cell Wall Rebuilding in Two Types of PVYNTN–Solanum tuberosum Interactions" Viruses 12, no. 1: 66. https://doi.org/10.3390/v12010066