Peste des Petits Ruminants at the Wildlife–Livestock Interface in the Northern Albertine Rift and Nile Basin, East Africa

 , , , , , , and add
Show full author list
, , , , , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
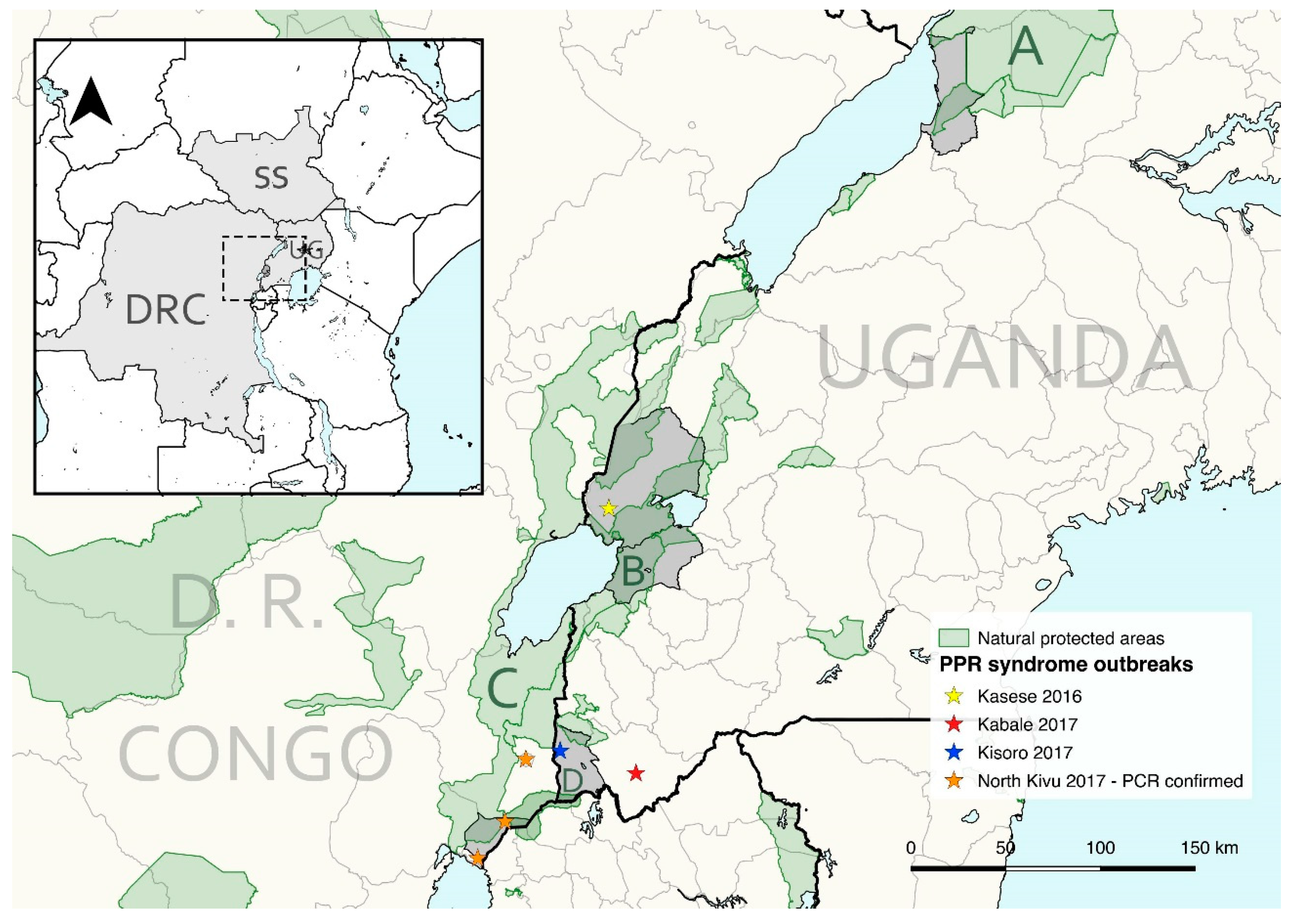
2.1. Study Areas
2.2. Sampling and Data
2.2.1. Western Uganda
2.2.2. South Sudan
2.2.3. Democratic Republic of the Congo
2.3. Laboratory analyses
2.4. Data analyses
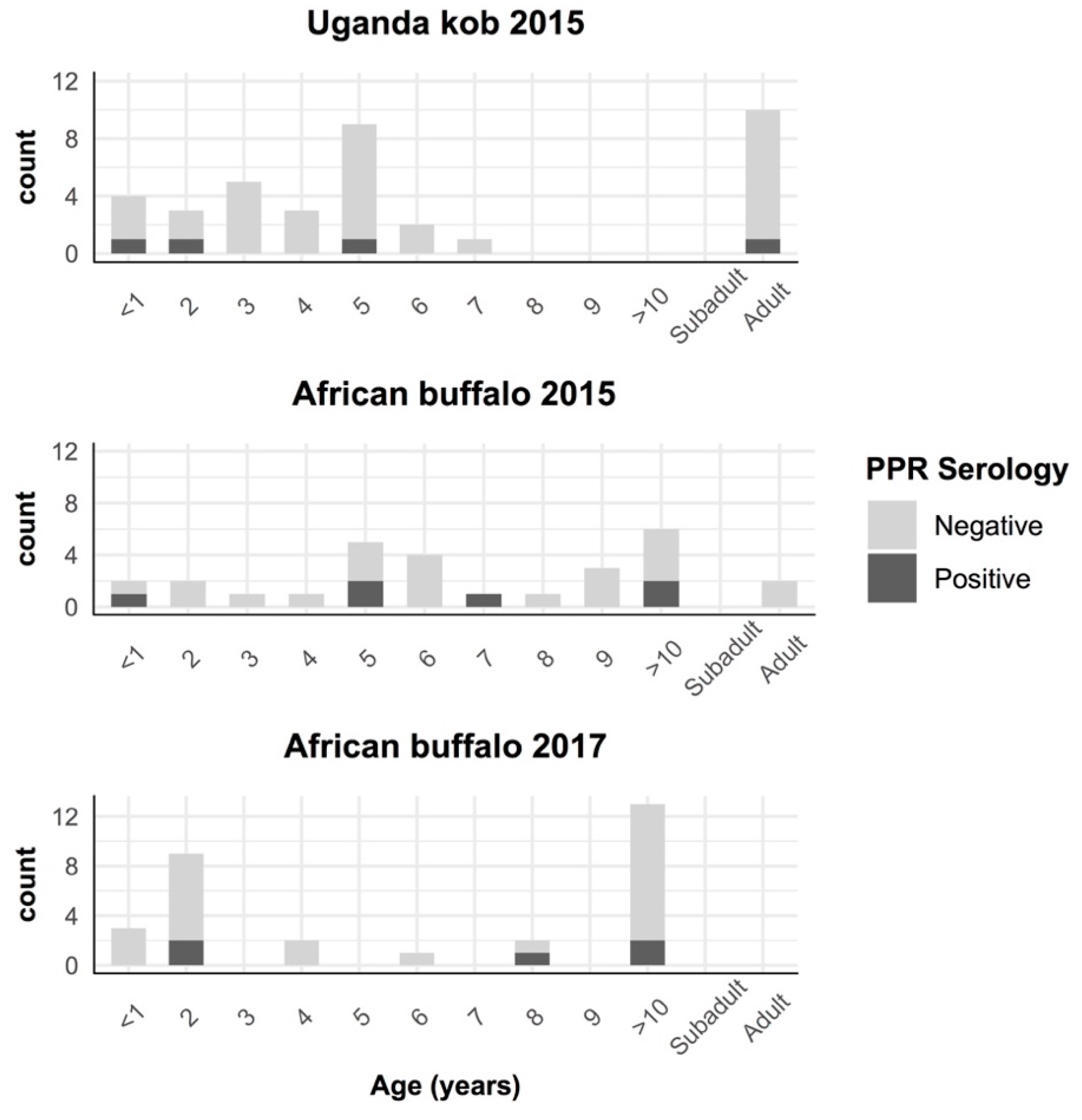
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parida, S.; Muniraju, M.; Mahapatra, M.; Muthuchelvan, D.; Buczkowski, H.; Banyard, A.C. Peste des petits ruminants. Vet. Microbiol. 2015, 181, 90–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE-FAO. Global Control and Eradication of Peste des Petits Ruminants. Investing in Veterinary Systems, Food Security and Poverty Alleviation. World Organization for Animal Health, United Nations Food and Agriculture Organisation, 2015. Available online: http://www.fao.org/3/a-i4477e.pdf (accessed on 30 May 2018).
- Jones, B.A.; Rich, K.M.; Mariner, J.C.; Anderson, J.; Jeggo, M.; Thevasagayam, S.; Cai, Y.; Peters, A.R.; Roeder, P. The economic impact of eradicating peste des petits ruminants: A benefit-cost analysis. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, X.F.; Fine, A.E.; Pruvot, M.; Njeumi, F.; Walzer, C.; Kock, R.; Shiilegdamba, E. PPR virus threatens wildlife conservation. Science 2018, 362, 165–166. [Google Scholar]
- Pruvot, M.; Strindberg, S.; Shiilegdamba, E.; Ganzorig, K.; Damdinjav, B.; Buuveibaatar, B.; Chimeddorj, B.; Bayandonoi, G.; Jargalsaikhan, T.; Shatar, M.; et al. Outbreak of Peste des Petits Ruminants among Critically Endangered Mongolian Saiga and Other Wild Ungulates, Mongolia, 2016–2017. Emerg. Infect. Dis. 2020, 26, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kock, R. Mongolia. Investigation of Peste des Petits Ruminants (PPR) among Wild Animals and Its Potential Impact on the Current PPR Situation in Livestock; Crisis Management Centre for Animal Health, Mission Report, Ed.; FAO: Rome, Italy, 2017; pp. 1–42. [Google Scholar]
- Parida, S.; Muniraju, M.; Altan, E.; Baazizi, R.; Raj, G.D.; Mahapatra, M. Emergence of PPR and its threat to Europe. Small Rumin. Res. 2016, 142, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Pope, R.A.; Parida, S.; Bailey, D.; Brownlie, J.; Barrett, T.; Banyard, A.C. Early Events following Experimental Infection with Peste-Des-Petits Ruminants Virus Suggest Immune Cell Targeting. PLoS ONE 2013, 8, e55830. [Google Scholar] [CrossRef]
- Truong, T.; Boshra, H.; Embury-Hyatt, C.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Chetty, T.; Mather, A.; et al. Peste des petits ruminants virus tissue tropism and pathogenesis in sheep and goats following experimental infection. PLoS ONE 2014, 9, e87145. [Google Scholar] [CrossRef]
- Parida, S.; Couacy-Hymann, E.; Pope, R.A.; Mahapatra, M.; El Harrak, M.; Brownlie, J.; Banyard, A.C. Pathology of Peste des Petits Ruminants. In Peste Des Petits Ruminants Virus; Munir, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 51–67. ISBN 9783662451656. [Google Scholar]
- World Organization for Animal Health (OIE). Peste des petits ruminants (infection with peste des petits ruminants virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019; OIE: Paris, France, 2013. [Google Scholar]
- Banyard, A.C.; Wang, Z.; Parida, S. Peste des petits ruminants virus, Eastern Asia. Emerg. Infect. Dis. 2014, 20, 2176–2178. [Google Scholar] [CrossRef] [Green Version]
- OIE-FAO (2015) Global Strategy for the Control and Eradication of PPR, World Organization for Animal Health, United Nations Food and Agriculture Organisation. Available online: http://www.oie.int/eng/ppr2015/doc/PPR-Global-Strategy-2015-03-28.pdf (accessed on 1 October 2018).
- Mahapatra, M.; Sayalel, K.; Muniraju, M.; Eblate, E.; Fyumagwa, R.; Shilinde, L.; Mdaki, M.; Keyyu, J.; Parida, S.; Kock, R. Spillover of Peste des Petits Ruminants Virus from Domestic to Wild Ruminants in the Serengeti Ecosystem, Tanzania. Emerg. Infect. Dis. 2015, 21, 2230–2234. [Google Scholar] [CrossRef] [Green Version]
- El Hag Ali, B.; Taylor, W. Isolation of peste des petits ruminants virus from the Sudan. Res. Vet. Sci. 1984, 36, 1–4. [Google Scholar] [CrossRef]
- Roeder, P.L.; Abraham, G.; Kenfe, G.; Barretf, T. Peste des Petits Ruminants in Ethiopian Goats. Trop. Anim. Health Prod. 1994, 26, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Wamwayi, H.M.; Rossiter, P.B.; Kariuki, D.P.; Wafula, J.S.; Barrett, T.; Anderson, J. Peste des petits ruminants antibodies in East Africa. Vet. Rec. 1995, 136, 199–200. [Google Scholar] [CrossRef] [PubMed]
- Karimuribo, E.D.; Loomu, P.M.; Mellau, L.S.B.; Swai, E.S. Retrospective study on sero-epidemiology of peste des petits ruminants before its official confirmation in northern Tanzania in 2008. Res. Opin. Anim. Vet. Sci. 2011, 1, 184–187. [Google Scholar]
- AU-IBAR Pan African Animal Health Yearbook 2003; Interafrican Bureau for Animal Resources and African Union: Nairobi, Kenya, 2004; pp. 1–73.
- Luka, P.D.; Erume, J.; Mwiine, F.N.; Ayebazibwe, C. Molecular characterization of peste des petits ruminants virus from the Karamoja region of Uganda (2007–2008). Arch. Virol. 2012, 157, 29–35. [Google Scholar] [CrossRef]
- Banyard, A.C.; Parida, S.; Batten, C.; Oura, C.; Kwiatek, O.; Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J. Gen. Virol. 2010, 91, 2885–2897. [Google Scholar] [CrossRef] [Green Version]
- Torsson, E.; Kgotlele, T.; Berg, M.; Mtui-Malamsha, N.; Swai, E.S.; Wensman, J.J.; Misinzo, G. History and current status of peste des petits ruminants virus in Tanzania. Infect. Ecol. Epidemiol. 2016, 6, 1–7. [Google Scholar] [CrossRef]
- Nkamwesiga, J.; Coffin-Schmitt, J.; Ochwo, S.; Mwiine, F.N.; Palopoli, A.; Ndekezi, C.; Isingoma, E.; Nantima, N.; Nsamba, P.; Adiba, R.; et al. Identification of peste des petits ruminants transmission hotspots in the Karamoja subregion of Uganda for targeting of eradication interventions. Front. Vet. Sci. 2019, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dundon, W.G.; Kihu, S.M.; Gitao, G.C.; Bebora, L.C.; John, N.M.; Oyugi, J.O.; Loitsch, A.; Diallo, A. Detection and Genome Analysis of a Lineage III Peste Des Petits Ruminants Virus in Kenya in 2011. Transbound. Emerg. Dis. 2015, 64, 644–650. [Google Scholar] [CrossRef] [Green Version]
- Kihu, S.M.; Gachohi, J.M.; Ndungu, E.K.; Gitao, G.C.; Bebora, L.C.; John, N.M.; Wairire, G.G.; Maingi, N.; Wahome, R.G.; Ireri, R. Sero-epidemiology of Peste des petits ruminants virus infection in Turkana County, Kenya. BMC Vet. Res. 2015, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Munir, M. Role of Wild Small Ruminants in the Epidemiology of Peste Des Petits Ruminants. Transbound. Emerg. Dis. 2014, 61, 411–424. [Google Scholar] [CrossRef]
- Wensman, J.J.; Abubakar, M.; Shabbir, M.Z.; Rossiter, P. Peste des petits ruminants in wild ungulates. Trop. Anim. Health Prod. 2018, 50, 1815–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, L.; Wu, X.; Liu, F.; Zou, Y.; Wang, Q.; Liu, C.; Bao, J.; Wang, W.; Ma, W.; et al. Diagnosis of Peste des Petits Ruminants in Wild and Domestic Animals in Xinjiang, China, 2013–2016. Transbound. Emerg. Dis. 2017, 64, e43–e47. [Google Scholar] [CrossRef] [PubMed]
- Marashi, M.; Masoudi, S.; Moghadam, M.K.; Modirrousta, H.; Marashi, M.; Parvizifar, M.; Dargi, M.; Saljooghian, M.; Homan, F.; Hoffmann, B.; et al. Peste des petits ruminants virus in vulnerable wild small ruminants, Iran, 2014–2016. Emerg. Infect. Dis. 2017, 23, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Couacy-Hymann, E.; Bodjo, C.; Danho, T.; Libeau, G.; Diallo, A. Surveillance of wildlife as a tool for monitoring rinderpest and peste des petits ruminants in West Africa. Rev. Sci. Tech. 2005, 24, 869–877. [Google Scholar]
- Kock, R.A.; Wamwayi, H.M.; Rossiter, P.B.; Libeau, G.; Wambwa, E.; Okori, J.; Shiferaw, F.S.; Mlengeya, T.D. Re-infection of wildlife populations with rinderpest virus on the periphery of the Somali ecosystem in East Africa. Prev. Vet. Med. 2006, 75, 63–80. [Google Scholar] [CrossRef]
- Asil, R.M.; Ludlow, M.; Ballal, A.; Alsarraj, S.; Ali, W.H.; Mohamed, B.A.; Mutwakil, S.M.; Osman, N.A. First detection and genetic characterization of peste des petits ruminants virus from dorcas gazelles “Gazella dorcas” in the Sudan, 2016-2017. Arch. Virol. 2019, 164, 2537–2543. [Google Scholar] [CrossRef]
- Meunier, N.V.; Sebulime, P.; White, R.G.; Kock, R. Wildlife-livestock interactions and risk areas for cross-species spread of bovine tuberculosis. Onderstepoort J. Vet. Res. 2017, 84, 1–10. [Google Scholar] [CrossRef]
- Miguel, E.; Grosbois, V.; Caron, A.; Boulinier, T.; Fritz, H.; Cornelis, D.; Foggin, C.; Makaya, P.V.; Tshabalala, P.T.; De Garine-Wichatitsky, M. Contacts and foot and mouth disease transmission from wild to domestic bovines in Africa. Ecosphere 2013, 4, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Fenton, A.; Pedersen, A.B. Community epidemiology framework for classifying disease threats. Emerg. Infect. Dis. 2005, 11, 1815–1821. [Google Scholar] [CrossRef]
- Plumptre, A.J.; Davenport, T.R.B.; Behangana, M.; Kityo, R.; Eilu, G.; Ssegawa, P.; Ewango, C.; Meirte, D.; Kahindo, C.; Herremans, M.; et al. The biodiversity of the Albertine Rift. Biol. Conserv. 2007, 134, 178–194. [Google Scholar] [CrossRef]
- Olupot, W.; Mcneilage, A.J.; Plumptre, A.J. An Analysis of Socioeconomics of Bushmeat Hunting at Major Hunting Sites in Uganda; Wildlife Conservation Society: Kampala, Uganda, 2009; pp. 1–108. [Google Scholar]
- Kock, R.A. Rinderpest and wildlife. In Rinderpest and Peste des Petits Ruminants Virus. Plagues of large and small ruminants; Barrett, T., Pastoret, P.-P., Taylor, W., Eds.; Elsevier Publications, Academic Press: London, UK, 2005; pp. 144–162. [Google Scholar]
- Libeau, G.; Lancelot, R.; Colas, F.; Guerre, L.; Bishop, D.H.L.; Diallo, A. Development of competitive ELISA for the peste des petits ruminants virus using a recombinant nucleoprotein. Res. Vet. Sci. 1995, 58, 50–55. [Google Scholar] [CrossRef]
- Clarke, B.; Mahapatra, M.; Friedgut, O.; Bumbarov, V.; Parida, S. Persistence of Lineage IV Peste-des-petits ruminants virus within Israel since 1993: An evolutionary perspective. PLoS ONE 2017, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Batten, C.A.; Banyard, A.C.; King, D.P.; Henstock, M.R.; Edwards, E.; Sanders, A.; Buczkowski, H.; Chris Oura, C.L.; Barrett, T. A real time RT-PCR assay for the specific detection of Peste des petits ruminants virus. J. Virol. Methods 2011, 171, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Muniraju, M.; Munir, M.; Parthiban, A.R.; Banyard, A.C.; Bao, J.; Wang, Z.; Ayebazibwe, C.; Ayelet, G.; El Harrak, M.; Mahapatra, M.; et al. Molecular Evolution of Peste des Petits Ruminants Virus. Emerg. Infect. Dis. 2014, 20, 2023–2033. [Google Scholar] [CrossRef] [Green Version]
- Baazizi, R.; Mahapatra, M.; Clarke, B.D.; Ait-Oudhia, K.; Khelef, D.; Parida, S. Peste des petits ruminants (PPR): A neglected tropical disease in maghreb region of North Africa and its threat to Europe. PLoS ONE 2017, 12, 1–16. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, M.; Nunes, T.; Heuer, C.; Marshall, J.; Sanchez, J.; Thornton, R.; Reiczigel, J.; Robison-Cox, J.; Sebastiani, P.; Solymos, P.; et al. Package epiR: An R Package for the Analysis of Epidemiological Data. R package version 1.0-11. Available online: https://cran.r-project.org/web/packages/epiR/epiR.pdf (accessed on 1 February 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- QGIS Development Team QGIS 2016. Las Palmas 2.18. Geogaphic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 1 February 2017).
- Tshilenge, M.G.; Walandila, J.S.; Byakya, D.; Masumu, J.; Katshay, L.; Cattoli, G.; Bushu, E.; Mpiana, S.; Dundon, W.G. Peste des petits ruminants viruses of lineages II and III identified in the Democratic Republic of the Congo. Vet. Microbiol. 2019, 239, 108493. [Google Scholar] [CrossRef]
- Niyokwishimira, A.; de D Baziki, J.; Dundon, W.G.; Nwankpa, N.; Njoroge, C.; Boussini, H.; Wamwayi, H.; Jaw, B.; Cattoli, G.; Nkundwanayo, C.; et al. Detection and molecular characterization of Peste des Petits Ruminants virus from outbreaks in Burundi, December 2017–January 2018. Transbound. Emerg. Dis. 2019, 66, 2067–2073. [Google Scholar] [CrossRef]
- Misinzo, G.; Kgotlele, T.; Muse, E.A.; Van Doorsselaere, J.; Berg, M.; Munir, M. Peste des Petits Ruminants Virus Lineage II and IV from Goats in Southern Tanzania During an Outbreak in 2011. Br. J. Virol. 2015, 2, 1–4. [Google Scholar]
- Tounkara, K.; Kwiatek, O.; Niang, M.; Abou Kounta Sidibe, C.; Sery, A.; Dakouo, M.; Salami, H.; Lo, M.M.; Ba, A.; Diop, M.; et al. Genetic Evidence for Transboundary Circulation of Peste Des Petits Ruminants Across West Africa. Front. Vet. Sci. 2019, 6, 1–7. [Google Scholar]
- Kock, R.A. The Role of Wildlife in the Epidemiology of Rinderpest in East and Central Africa 1994-2004: A Study Based on Serological Surveillance and Disease Investigation. Ph.D. Thesis, University of Cambridge, Cambridge, UK, 2008. [Google Scholar]
- Nakayima, J.; Nanfuka, M.; Kidega, E.; Kajuna, Y.; Ndumu, D. Epidemiological Insights into the Occurrence of Peste des Petits Ruminants Virus (PPRV) Among Sheep, Goats and Cattle in Western Uganda. Int. J. Livest. Res. 2018, 8, 73. [Google Scholar] [CrossRef]
- Field, C.R.; Laws, R.M. The distribution of the larger herbivores in the Queen Elizabeth National Park, Uganda. J. Appl. Ecol. 1970, 7, 273–294. [Google Scholar] [CrossRef]
- Lembo, T.; Oura, C.; Parida, S.; Hoare, R.; Frost, L.; Fyumagwa, R.; Kivaria, F.; Chubwa, C.; Kock, R.; Cleaveland, S.; et al. Peste des petits ruminants infection among cattle and wildlife in northern Tanzania. Emerg. Infect. Dis. 2013, 19, 2037–2040. [Google Scholar] [CrossRef] [Green Version]
- Marjan, M.D. Movements and Conservation of the Migratory white-eared kob (Kobus kob leucotis) in South Sudan. Ph.D. Thesis, University of Massachusetts Amherst, Amherst, MA, USA, September 2014. [Google Scholar]
- Sen, A.; Saravanan, P.; Balamurugan, V.; Bhanuprakash, V.; Venkatesan, G.; Sarkar, J.; Rajak, K.K.; Ahuja, A.; Yadav, V.; Sudhakar, S.B.; et al. Detection of subclinical peste des petits ruminants virus infection in experimental cattle. Virus Dis. 2014, 25, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Agga, G.E.; Raboisson, D.; Walch, L.; Alemayehu, F. Epidemiological Survey of Peste des Petits Ruminants in Ethiopia: Cattle as Potential Sentinel for Surveillance. Front. Vet. Sci. 2019, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Fakri, F.Z.; Elhajjam, A.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Tadlaoui, K.; Fassi-Fihri, O.; Elharrak, M. Susceptibility of Moroccan sheep and goat breeds to peste des petits ruminants virus. Acta Vet. Scand. 2017, 59, 1–6. [Google Scholar] [CrossRef]
- Hoffmann, B.; Wiesner, H.; Maltzan, J.; Mustefa, R.; Eschbaumer, M.; Arif, F.A.; Beer, M. Fatalities in Wild Goats in Kurdistan Associated with Peste Des Petits Ruminants Virus. Transbound. Emerg. Dis. 2011, 59, 173–176. [Google Scholar] [CrossRef]
- Abubakar, M.; Rajput, Z.I.; Arshed, M.J.; Sarwar, G.; Ali, Q. Evidence of peste des petits ruminants virus (PPRV) infection in Sindh Ibex (Capra aegagrus blythi) in Pakistan as confirmed by detection of antigen and antibody. Trop. Anim. Health Prod. 2011, 43, 745–747. [Google Scholar] [CrossRef]
- Caron, A.; Cappelle, J.; Cumming, G.S.; De Garine-Wichatitsky, M.; Gaidet, N. Bridge hosts, a missing link for disease ecology in multi-host systems. Vet. Res. 2015, 46, 1–11. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Year | Individuals | Herds | |||||
|---|---|---|---|---|---|---|---|---|
| n | Positives | Prev.% (CI 95%) | n | Positives | Prev.% (CI 95%) | |||
| Uganda—Queen Elizabeth National Park | ||||||||
| Buffalo | Syncerus caffer | 2015 | 28 | 6 | 21.4 (10.2–39.5) | 14 | 4 | 28.6 (11.7–54.6) |
| 2017 | 30 | 5 | 16.7 (7.3–33.6) | 14 | 5 | 35.7 (16.3–61.2) | ||
| Total | 58 | 11 | 19.0 (10.9–30.8) | 28 | 9 | 32.1 (17.9–50.7) | ||
| Uganda kob | Kobus kob thomasi | 2015 | 39 | 4 | 10.3 (4.1–23.6) | 20 | 3 | 15.0 (5.2–36.0) |
| All species | 97 | 15 | 15.5 (9.6–24.0) | 48 | 13 | 27.1 (16.6–41.0) | ||
| South Sudan—Nile basin and Boma National Park | ||||||||
| Eland | Taurotragus oryx | 2013 | 9 | 0 | 0.0 (0.0–29.9) | NA | NA | NA |
| Elephant | Loxodonta africana | 2013 | 40 | 1 | 2.5 (0.1–12.9) | NA | NA | NA |
| Tiang | Damaliscus lunatus tiang | 2013 | 21 | 15 | 71.4 (50.0–86.2) | NA | NA | NA |
| White eared Kob | Kobus kob leucotis | 2013 | 17 | 0 | 0.0 (0.0–18.4) | NA | NA | NA |
| All species | 87 | 16 | 18.4 (11.6–27.8) | |||||
| 2015 January–February | 2016 June–July | 2017 June–August | Total | |||||
|---|---|---|---|---|---|---|---|---|
| n | Prev.% (CI 95%) | n | Prev.% (CI 95%) | n | Prev.% (CI 95%) | n | Prev.% (CI 95%) | |
| Goats | ||||||||
| MFNP—Bulisa district | 15 | 0.0 (0.0–20.7) | NA | NA | NA | NA | 15 | 0.0 (0.0–20.7) |
| QENP—Kasese district | 15 | 0.0 (0.0–20.7) | 23 | 0.0 (0.0–14.2) | 160 | 9.6 (5.6–15.6) | 198 | 7.6 (4.3–12.5) |
| QENP—Rubirizi district | 72 | 0.0 (0.0–4.3) | 20 | 4.2 (0.0–24.2) | 0 | NA | 92 | 0.1 (0.0–5.2) |
| MGNP—Kisoro district | 59 | 0.0 (0.0–5.5) | 52 | 1.0 (0.0–9.7) | 139 | 0.0 (0.0–1.8) | 250 | 0.0 (0.0–1.3) |
| Sheep | ||||||||
| MFNP—Bulisa district | 5 | 0.0 (0.0–45.4) | 0 | NA | 0 | NA | 5 | 0.0 (0.0–45.4) |
| QENP—Kasese district | 0 | NA | 7 | 29.5 (7.7–67.5) | 20 | 20.3 (7.6–43.4) | 27 | 22.2 (10.3–42.5) |
| QENP—Rubirizi district | 8 | 0.0 (0.0–33.6) | 6 | 0.0 (0.0–40.7) | 0 | NA | 14 | 0.0 (0.0–22.0) |
| MGNP—Kisoro district | 53 | 0.0 (0.0–6.2) | 47 | 3.5 (0.2–14.2) | 107 | 0.0 (0.0–4.4) | 207 | 0.5 (0.0–3.4) |
| Cattle | ||||||||
| MFNP—Bulisa district | 0 | NA | 0 | NA | 0 | NA | 0 | NA |
| QENP—Kasese district | 137 | 2.8 (0.6–7.8) | 60 | 39.9 (27.9–53.5) | 50 | 39.6 (26.6–54.4) | 247 | 19.3 (14.6–25.0) |
| QENP—Rubirizi district | 3 | 0.0 (0.0–60.0) | 0 | NA | 0 | NA | 3 | 0.0 (0.0–60.0) |
| MGNP—Kisoro district | 39 | 0.0 (0.0–8.5) | 51 | 3.1 (0.1–13.1) | 146 | 0.0 (0.0–3.0) | 236 | 0.3 (0.0–2.9) |
| Sample No. | Species | Sex | Age | CT-Value |
|---|---|---|---|---|
| DRC-1 | Goat | F | 10 months | 18.16 |
| DRC-2 | Goat | F | 4 years | 29.10 |
| DRC-3 | Goat | F | 4 years | 32.79 |
| DRC-4 | Goat | F | 3 years | 28.06 |
| DRC-5 | Goat | F | 1.5 years | 24.79 |
| DRC-6 | Goat | F | 1 year | 34.11 |
| DRC-7 | Goat | F | 1 year | 29.68 |
| DRC-8 | Sheep | F | 2 years | UD |
| DRC-9 | Sheep | F | 2 years | 32.11 |
| DRC-10 | Sheep | F | 3 years | 25.09 |
| DRC-11 | Sheep | M | 1.5 years | UD |
| DRC-12 | Sheep | F | 4 years | 27.67 |
| DRC-13 | Goat | F | 3 years | 22.17 |
| DRC-14 | Goat | F | 1 year | 20.23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez Aguilar, X.; Mahapatra, M.; Begovoeva, M.; Kalema-Zikusoka, G.; Driciru, M.; Ayebazibwe, C.; Adwok, D.S.; Kock, M.; Lukusa, J.-P.K.; Muro, J.; et al. Peste des Petits Ruminants at the Wildlife–Livestock Interface in the Northern Albertine Rift and Nile Basin, East Africa. Viruses 2020, 12, 293. https://doi.org/10.3390/v12030293
Fernandez Aguilar X, Mahapatra M, Begovoeva M, Kalema-Zikusoka G, Driciru M, Ayebazibwe C, Adwok DS, Kock M, Lukusa J-PK, Muro J, et al. Peste des Petits Ruminants at the Wildlife–Livestock Interface in the Northern Albertine Rift and Nile Basin, East Africa. Viruses. 2020; 12(3):293. https://doi.org/10.3390/v12030293
Chicago/Turabian StyleFernandez Aguilar, Xavier, Mana Mahapatra, Mattia Begovoeva, Gladys Kalema-Zikusoka, Margaret Driciru, Chrisostom Ayebazibwe, David Solomon Adwok, Michael Kock, Jean-Paul Kabemba Lukusa, Jesus Muro, and et al. 2020. "Peste des Petits Ruminants at the Wildlife–Livestock Interface in the Northern Albertine Rift and Nile Basin, East Africa" Viruses 12, no. 3: 293. https://doi.org/10.3390/v12030293