
Dengue Virus Infection: A Tale of Viral Exploitations and Host Responses

 and
and 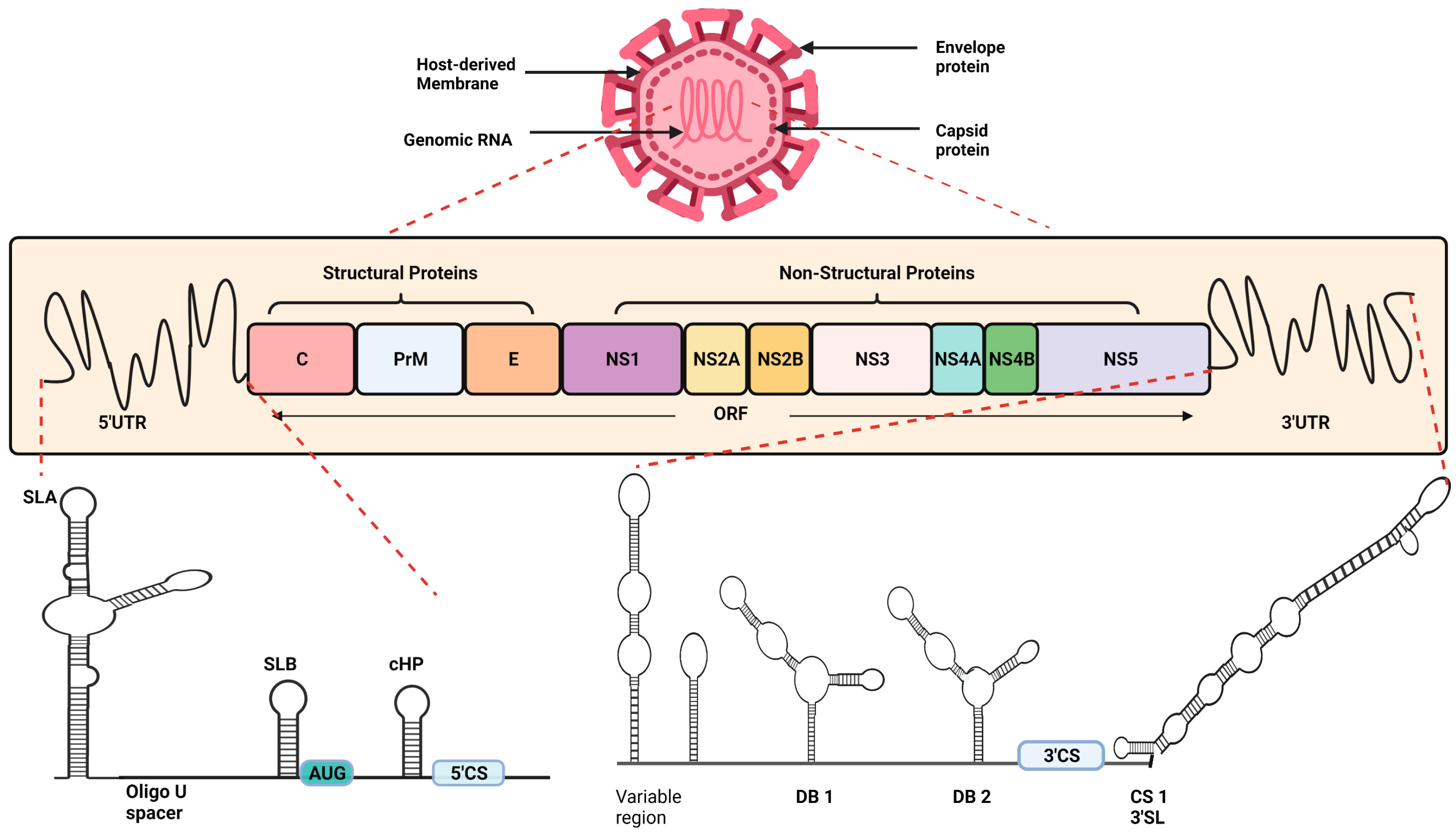{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
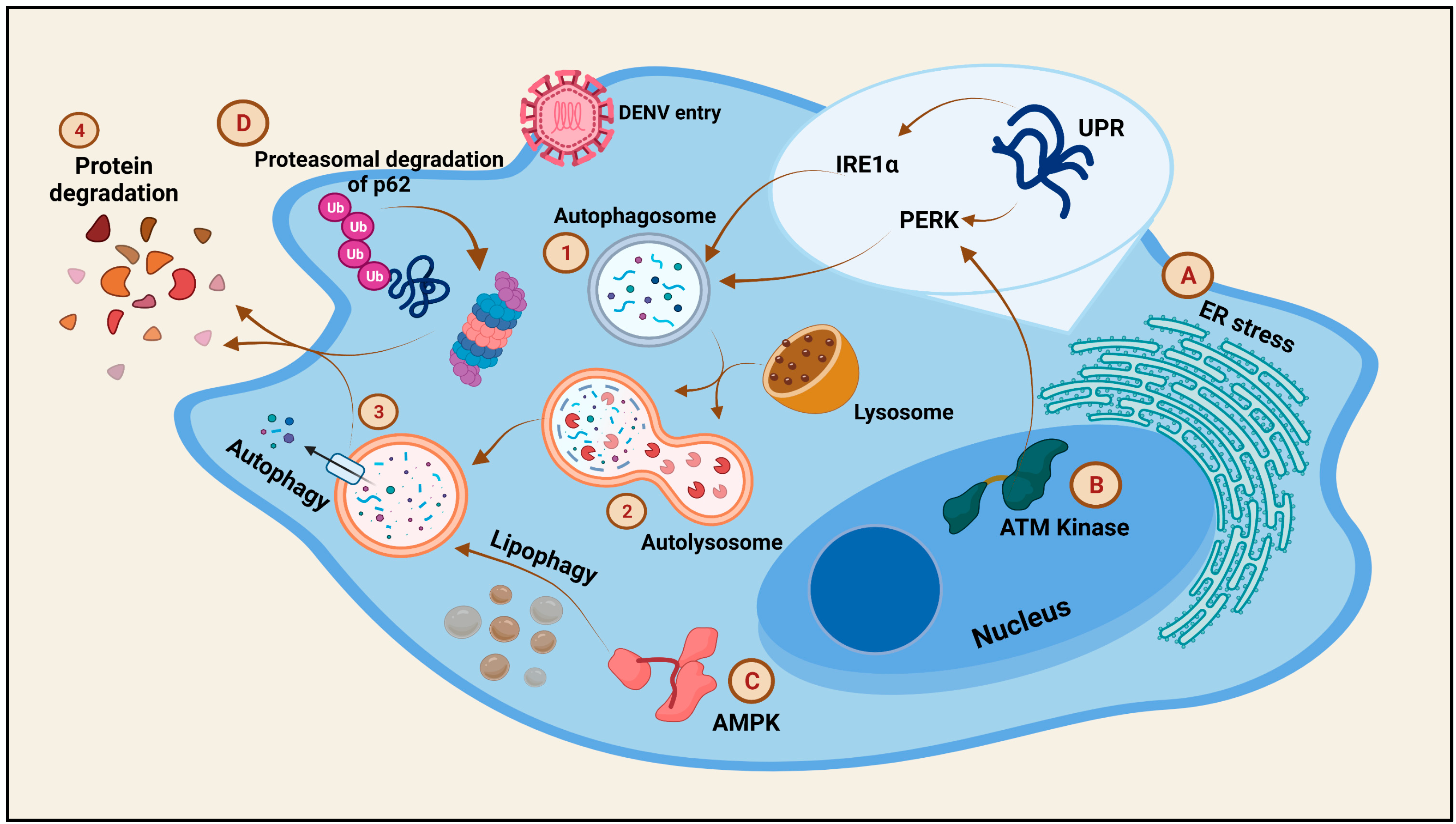{kind=link}
Abstract
:1. Introduction
2. Structural Details of Dengue Virus
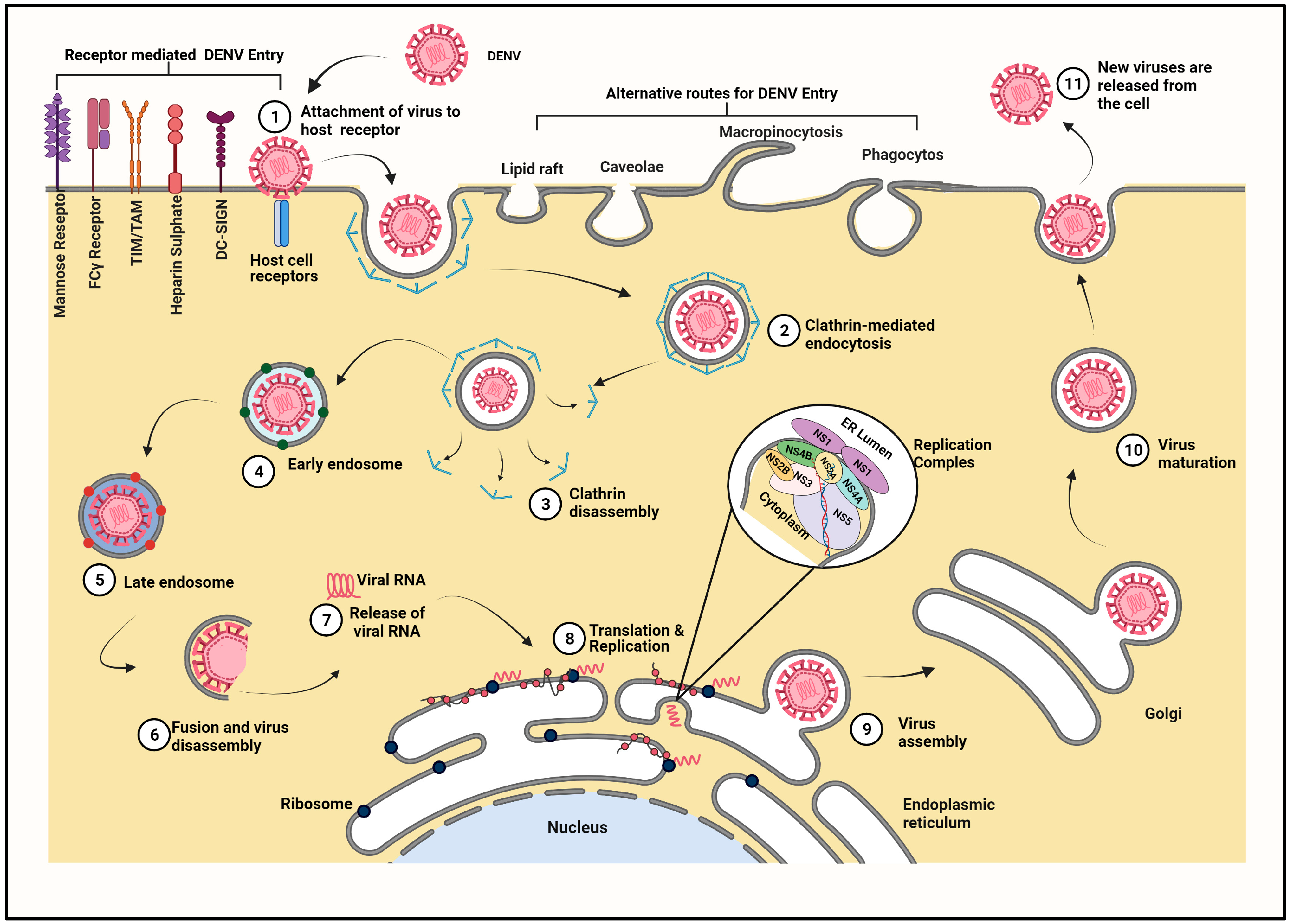
3. Transmission and Replication Cycle of Dengue Virus inside Mammalian Cell
3.1. Entry of Virus Particle and Role of Cell Surface Receptors
3.2. Fusion and Release of Viral Genome
3.3. Transport of Viral Genome
3.4. Viral Replication
3.5. Assembly of Viral Proteins
3.6. Maturation and Egress of Virus
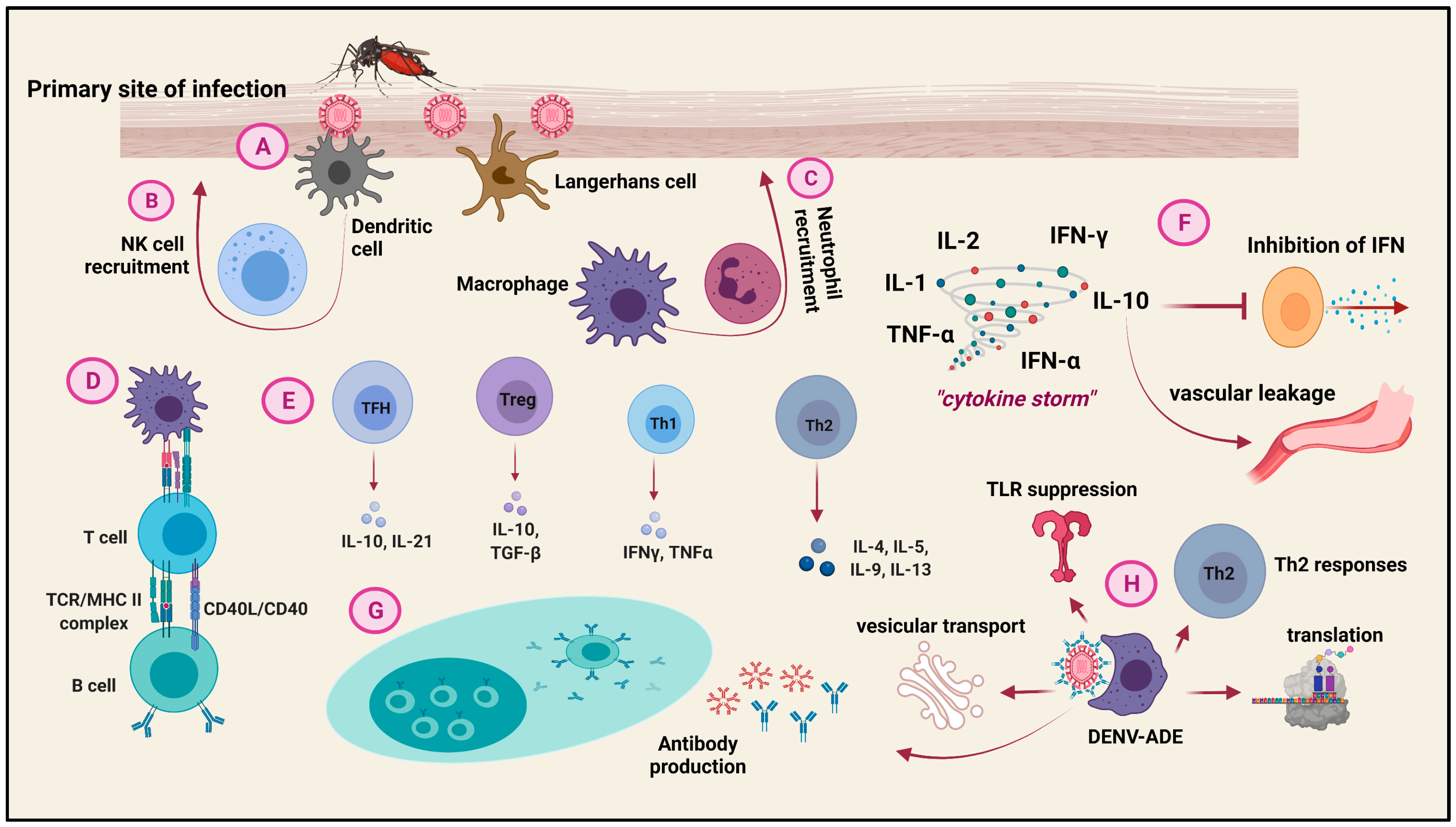
4. Pathogenesis of Dengue Virus inside the Host
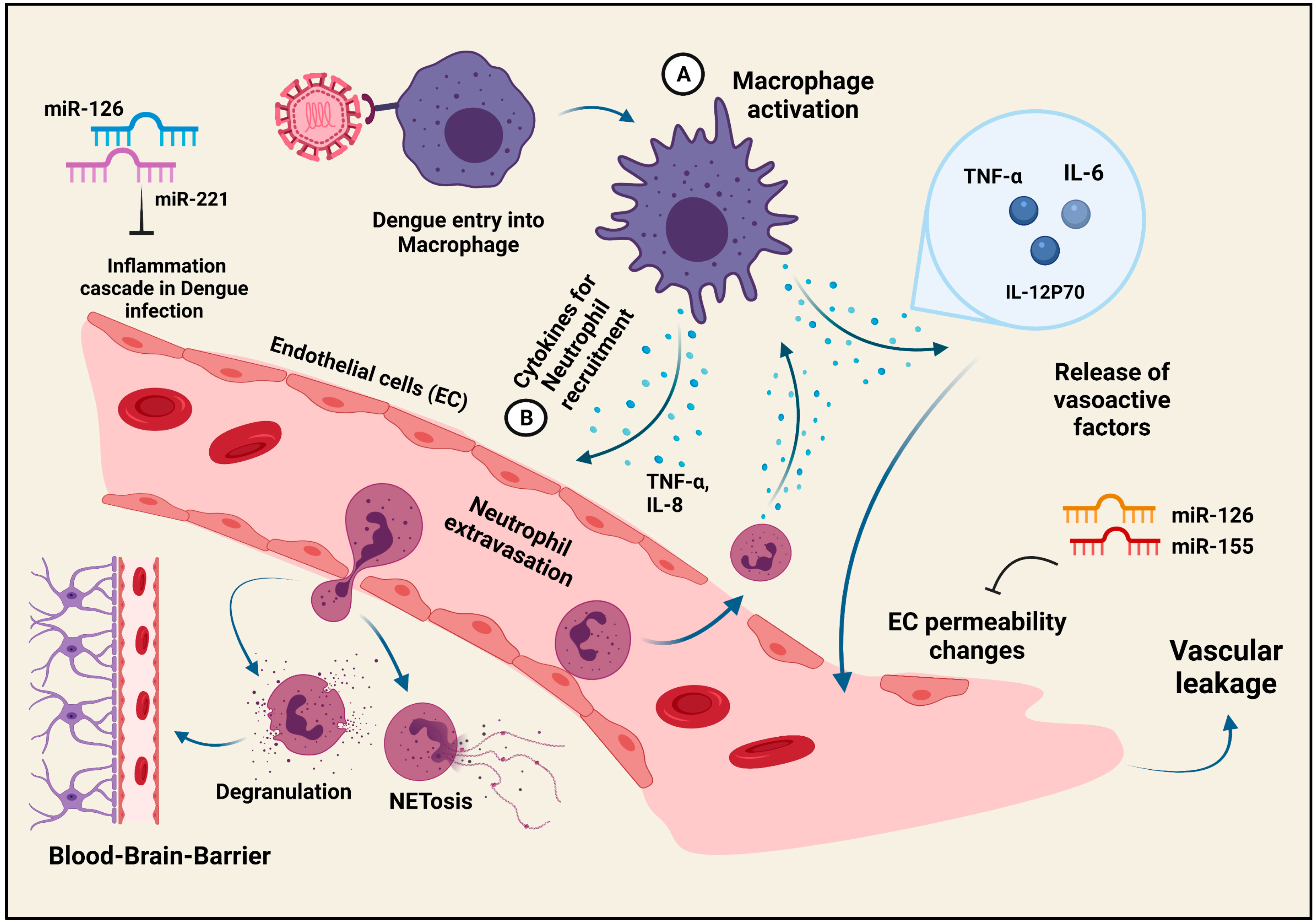
5. Inflammation during Dengue Virus Infection
6. Host Immune Cell Responses to Dengue Virus Infection
7. Cytokine Storm during Dengue Infection
8. Intrinsic Antibody Dependent Enhancement (ADE) during Dengue Infection
9. Dengue Virus Induced Cell Death
9.1. Regulation of Apoptosis during Dengue Virus Infection
9.2. Regulation of Autophagy during Dengue Virus Infection
10. Antivirals against Dengue Infection
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Dengue and Severe Dengue; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Roy, S.K.; Bhattacharjee, S. Dengue virus: Epidemiology, biology, and disease aetiology. Can. J. Microbiol. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Züst, R.; Toh, Y.X.; Pfaff, J.M.; Kahle, K.M.; Davidson, E.; Doranz, B.J.; Velumani, S.; Tukijan, F.; Wang, C.-I.; et al. Protective Capacity of the Human Anamnestic Antibody Response during Acute Dengue Virus Infection. J. Virol. 2016, 90, 11122–11131. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, M.; Rasotgi, V.; Jain, S.; Gupta, V. Discovery of fifth serotype of dengue virus (DENV-5): A new public health dilemma in dengue control. Med. J. Armed Forces India 2015, 71, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Harapan, H.; Michie, A.; Sasmono, R.T.; Imrie, A. Dengue: A Minireview. Viruses 2020, 12, 829. [Google Scholar] [CrossRef]
- Murugesan, A.; Manoharan, M. Emerging and Reemerging Viral Pathogens; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 978-0-12-819400-3. [Google Scholar]
- O’Connor, O.; Ou, T.P.; Aubry, F.; Dabo, S.; Russet, S.; Girault, D.; In, S.; Minier, M.; Hoem, T.; Boyer, S.; et al. Potential role of vector-mediated natural selection in dengue virus genotype/lineage replacements in two epidemiologically contrasted settings. Emerg. Microbes Infect. 2021, 10, 1346–1357. [Google Scholar] [CrossRef]
- Urcuqui-Inchima, S.; Patiño, C.; Torres, S.; Haenni, A.-L.; Díaz, F.J. Recent Developments in Understanding Dengue Virus Replication. Adv. Virus Res. 2010, 77, 1–39. [Google Scholar] [CrossRef]
- Cruz-Oliveira, C.; Freire, J.M.; Conceição, T.M.; Higa, L.M.; Castanho, M.; Da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol. Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begum, F.; Das, S.; Mukherjee, D.; Ray, U. Hijacking the Host Immune Cells by Dengue Virus: Molecular Interplay of Receptors and Dengue Virus Envelope. Microorganisms 2019, 7, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, E.G.; Kumar, A.; Bartenschlager, R. Revisiting Dengue Virus–Host Cell Interaction: New insights into molecular and cellular virology. Adv. Virus Res. 2014, 88, 1–109. [Google Scholar] [CrossRef] [PubMed]
- Carro, S.D.; Cherry, S. Beyond the Surface: Endocytosis of Mosquito-Borne Flaviviruses. Viruses 2020, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lok, S.-M.; Yu, I.-M.; Zhang, Y.; Kuhn, R.J.; Chen, J.; Rossmann, M.G. The Flavivirus Precursor Membrane-Envelope Protein Complex: Structure and Maturation. Science 2008, 319, 1830–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaitseva, E.; Yang, S.-T.; Melikov, K.; Pourmal, S.; Chernomordik, L.V. Dengue Virus Ensures Its Fusion in Late Endosomes Using Compartment-Specific Lipids. PLOS Pathog. 2010, 6, e1001131. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, C.M.; Sevvana, M.; Kuhn, R.J. Structure-guided paradigm shifts in flavivirus assembly and maturation mechanisms. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2020; Volume 108, pp. 33–83. ISBN 978-0-12-820761-1. [Google Scholar]
- Marianneau, P.; Cardona, A.; Edelman, L.; Deubel, V.; Desprès, P. Dengue virus replication in human hepatoma cells activates NF-kappaB which in turn induces apoptotic cell death. J. Virol. 1997, 71, 3244–3249. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.L.; Inoue, T.; Chen, Y.-J.; Chang, A.; Tsai, B.; Tai, A.W. The ER Membrane Protein Complex Promotes Biogenesis of Dengue and Zika Virus Non-structural Multi-pass Transmembrane Proteins to Support Infection. Cell Rep. 2019, 27, 1666–1674.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, A.M.; Shurtleff, M.J.; Popova, K.D.; Kulsuptrakul, J.; Weissman, J.S.; Puschnik, A.S. The ER membrane protein complex is required to ensure correct topology and stable expression of flavivirus polyproteins. eLife 2019, 8, e48469. [Google Scholar] [CrossRef]
- Barrows, N.J.; Anglero-Rodriguez, Y.; Kim, B.; Jamison, S.F.; Le Sommer, C.; McGee, C.E.; Pearson, J.L.; Dimopoulos, G.; Ascano, M.; Bradrick, S.S.; et al. Dual roles for the ER membrane protein complex in flavivirus infection: Viral entry and protein biogenesis. Sci. Rep. 2019, 9, 9711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, D.W.; Campos, R.K.; Child, J.R.; Zheng, T.; Chan, K.W.K.; Bradrick, S.S.; Vasudevan, S.; Garcia-Blanco, M.A.; Nicchitta, C.V. Dengue Virus Selectively Annexes Endoplasmic Reticulum-Associated Translation Machinery as a Strategy for Co-opting Host Cell Protein Synthesis. J. Virol. 2018, 92, e01766-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamarnik, A.V.; Andino, R. Switch from translation to RNA replication in a positive-stranded RNA virus. Genes Dev. 1998, 12, 2293–2304. [Google Scholar] [CrossRef] [Green Version]
- Van den Elsen, K.; Quek, J.; Luo, D. Molecular Insights into the Flavivirus Replication Complex. Viruses 2021, 13, 956. [Google Scholar] [CrossRef]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.E.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and Three-Dimensional Architecture of the Dengue Virus Replication and Assembly Sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Bujalowski, P.J.; Teramoto, T.; Gottipati, K.; Scott, S.D.; Padmanabhan, R.; Choi, K.H. Structures of flavivirus RNA promoters suggest two binding modes with NS5 polymerase. Nat. Commun. 2021, 12, 2530. [Google Scholar] [CrossRef]
- Iglesias, N.G.; Filomatori, C.V.; Gamarnik, A.V. The F1 Motif of Dengue Virus Polymerase NS5 Is Involved in Promoter-Dependent RNA Synthesis. J. Virol. 2011, 85, 5745–5756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, D.E.; Lodeiro, M.F.; Ludueña, S.J.; Pietrasanta, L.I.; Gamarnik, A.V. Long-Range RNA-RNA Interactions Circularize the Dengue Virus Genome. J. Virol. 2005, 79, 6631–6643. [Google Scholar] [CrossRef] [Green Version]
- Choi, K. The Role of the Stem-Loop A RNA Promoter in Flavivirus Replication. Viruses 2021, 13, 1107. [Google Scholar] [CrossRef]
- Barnard, T.R.; Abram, Q.H.; Lin, Q.F.; Wang, A.B.; Sagan, S.M. Molecular Determinants of Flavivirus Virion Assembly. Trends Biochem. Sci. 2021, 46, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zou, J.; Zhang, X.; Zhou, Y.; Routh, A.; Kang, C.; Popov, V.L.; Chen, X.; Wang, Q.-Y.; Dong, H.; et al. Dengue NS2A Protein Orchestrates Virus Assembly. Cell Host Microbe 2019, 26, 606–622.e8. [Google Scholar] [CrossRef] [PubMed]
- Patkar, C.G.; Kuhn, R.J. Yellow Fever Virus NS3 Plays an Essential Role in Virus Assembly Independent of Its Known Enzymatic Functions. J. Virol. 2008, 82, 3342–3352. [Google Scholar] [CrossRef] [Green Version]
- Voßmann, S.; Wieseler, J.; Kerber, R.; Kümmerer, B.M. A Basic Cluster in the N Terminus of Yellow Fever Virus NS2A Contributes to Infectious Particle Production. J. Virol. 2015, 89, 4951–4965. [Google Scholar] [CrossRef] [Green Version]
- Byk, L.A.; Gamarnik, A.V. Properties and Functions of the Dengue Virus Capsid Protein. Annu. Rev. Virol. 2016, 3, 263–281. [Google Scholar] [CrossRef] [Green Version]
- Bressanelli, S.; Stiasny, K.; Allison, S.L.; Stura, E.; Duquerroy, S.; Lescar, J.; Heinz, F.X.; Rey, F. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. EMBO J. 2004, 23, 728–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, K.; Allison, S.L.; Schalich, J.; Heinz, F.X. Proteolytic activation of tick-borne encephalitis virus by furin. J. Virol. 1997, 71, 8475–8481. [Google Scholar] [CrossRef] [Green Version]
- Wengler, G. Cell-associated West Nile flavivirus is covered with E+pre-M protein heterodimers which are destroyed and reorganized by proteolytic cleavage during virus release. J. Virol. 1989, 63, 2521–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, I.-M.; Holdaway, H.A.; Chipman, P.R.; Kuhn, R.J.; Rossmann, M.G.; Chen, J. Association of the pr Peptides with Dengue Virus at Acidic pH Blocks Membrane Fusion. J. Virol. 2009, 83, 12101–12107. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Lu, X.; Li, J.; Liu, Y. Novel binding between pre-membrane protein and vacuolar ATPase is required for efficient dengue virus secretion. Biochem. Biophys. Res. Commun. 2008, 373, 319–324. [Google Scholar] [CrossRef]
- Limjindaporn, T.; Wongwiwat, W.; Noisakran, S.; Srisawat, C.; Netsawang, J.; Puttikhunt, C.; Kasinrerk, W.; Avirutnan, P.; Thiemmeca, S.; Sriburi, R.; et al. Interaction of dengue virus envelope protein with endoplasmic reticulum-resident chaperones facilitates dengue virus production. Biochem. Biophys. Res. Commun. 2009, 379, 196–200. [Google Scholar] [CrossRef]
- Perera, R.; Kuhn, R.J. Structural proteomics of dengue virus. Curr. Opin. Microbiol. 2008, 11, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, I.-M.; Zhang, W.; Holdaway, H.A.; Li, L.; Kostyuchenko, V.A.; Chipman, P.R.; Kuhn, R.J.; Rossmann, M.G.; Chen, J. Structure of the Immature Dengue Virus at Low pH Primes Proteolytic Maturation. Science 2008, 319, 1834–1837. [Google Scholar] [CrossRef]
- Mason, P.W.; Pincus, S.; Fournier, M.J.; Mason, T.L.; Shope, R.E.; Paoletti, E. Japanese encephalitis virus-vaccinia recombinants produce particulate forms of the structural membrane proteins and induce high levels of protection against lethal JEV infection. Virology 1991, 180, 294–305. [Google Scholar] [CrossRef]
- John, A.S.; Abraham, S.N.; Gubler, D.J. Barriers to preclinical investigations of anti-dengue immunity and dengue pathogenesis. Nat. Rev. Genet. 2013, 11, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Marchette, N.J.; Halstead, S.B.; Falkler, W.A.; Stenhouse, A.; Nash, D. Studies on the Pathogenesis of Dengue Infection in Monkeys. III. Sequential Distribution of Virus in Primary and Heterologous Infections. J. Infect. Dis. 1973, 128, 23–30. [Google Scholar] [CrossRef]
- Whitehorn, J.; Simmons, C.P. The pathogenesis of dengue. Vaccine 2011, 29, 7221–7228. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Kyle, J.L.; Harris, E. Global Spread and Persistence of Dengue. Annu. Rev. Microbiol. 2008, 62, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloia, A.L.; Abraham, A.M.; Bonder, C.S.; Pitson, S.; Carr, J. Dengue Virus-Induced Inflammation of the Endothelium and the Potential Roles of Sphingosine Kinase-1 and MicroRNAs. Mediat. Inflamm. 2015, 2015, 509306. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Wang, S.-Y. Activation of Terminally Differentiated Human Monocytes/Macrophages by Dengue Virus: Productive Infection, Hierarchical Production of Innate Cytokines and Chemokines, and the Synergistic Effect of Lipopolysaccharide. J. Virol. 2002, 76, 9877–9887. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.M.; Hocking, H.; Bunting, K.; Wright, P.J.; Davidson, A.; Gamble, J.; Burrell, C.J.; Li, P. Supernatants from dengue virus type-2 infected macrophages induce permeability changes in endothelial cell monolayers. J. Med Virol. 2003, 69, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.-J.; Wang, J.-J.; Shaio, M.-F.; Kao, C.-L.; Chang, D.-M.; Han, S.-W.; Lai, J.-H. Infection of Human Dendritic Cells by Dengue Virus Causes Cell Maturation and Cytokine Production. J. Immunol. 2001, 166, 1499–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luplerdlop, N.; Missé, D.; Bray, D.; Deleuze-Marquès, V.; Gonzalez, J.-P.; Leardkamolkarn, V.; Yssel, H.; Veas, F. Dengue-virus-infected dendritic cells trigger vascular leakage through metalloproteinase overproduction. EMBO Rep. 2006, 7, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Da Conceição, T.M.; Rust, N.M.; Berbel, A.C.E.R.; Martins, N.B.; Santos, C.A.D.N.; Da Poian, A.; de Arruda, L.B. Essential role of RIG-I in the activation of endothelial cells by dengue virus. Virology 2013, 435, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvert, J.K.; Helbig, K.J.; Dimasi, D.; Cockshell, M.; Beard, M.R.; Pitson, S.M.; Bonder, C.S.; Carr, J.M. Dengue Virus Infection of Primary Endothelial Cells Induces Innate Immune Responses, Changes in Endothelial Cells Function and Is Restricted by Interferon-Stimulated Responses. J. Interf. Cytokine Res. 2015, 35, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Wati, S.; Rawlinson, S.; Ivanov, R.A.; Dorstyn, L.; Beard, M.R.; Jans, D.; Pitson, S.; Burrell, C.J.; Li, P.; Carr, J. Tumour necrosis factor alpha (TNF-α) stimulation of cells with established dengue virus type 2 infection induces cell death that is accompanied by a reduced ability of TNF-α to activate nuclear factor B and reduced sphingosine kinase-1 activity. J. Gen. Virol. 2010, 92, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Sindi, N. Dengue Virus: Infection, Immunological Response, and Vaccine Development. J. Pharm. Res. Int. 2021, 33, 1–19. [Google Scholar] [CrossRef]
- Opasawatchai, A.; Amornsupawat, P.; Jiravejchakul, N.; Chan-In, W.; Spoerk, N.J.; Manopwisedjaroen, K.; Singhasivanon, P.; Yingtaweesak, T.; Suraamornkul, S.; Mongkolsapaya, J.; et al. Neutrophil Activation and Early Features of NET Formation Are Associated with Dengue Virus Infection in Human. Front. Immunol. 2019, 9, 3007. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.L.; Paoletti, C.; Campisi, M.; Osaki, T.; Adriani, G.; Kamm, R.D.; Mattu, C.; Chiono, V. MicroRNA delivery through nanoparticles. J. Control. Release 2019, 313, 80–95. [Google Scholar] [CrossRef]
- Qi, Y.; Li, Y.; Zhang, L.; Huang, J. microRNA expression profiling and bioinformatic analysis of dengue virus-infected peripheral blood mononuclear cells. Mol. Med. Rep. 2013, 7, 791–798. [Google Scholar] [CrossRef] [Green Version]
- Tambyah, P.A.; Ching, C.S.; Sepramaniam, S.; Ali, J.M.; Armugam, A.; Jeyaseelan, K. microRNA expression in blood of dengue patients. Ann. Clin. Biochem. Int. J. Lab. Med. 2016, 53, 466–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; He, Z.; Hu, Y.; Wen, W.; Lin, C.; Yu, J.; Pan, J.; Li, R.; Deng, H.; Liao, S.; et al. MicroRNA-30e* Suppresses Dengue Virus Replication by Promoting NF-κB–Dependent IFN Production. PLoS Neglected Trop. Dis. 2014, 8, e3088. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.-F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.R.; Srivastava, D. miR-126 Regulates Angiogenic Signaling and Vascular Integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.-X.; Zeng, D.-Y.; Li, R.-T.; Pang, R.-P.; Yang, H.; Hu, Y.-L.; Zhang, Q.; Jiang, Y.; Huang, L.-Y.; Tang, Y.-B.; et al. Essential Role of MicroRNA-155 in Regulating Endothelium-Dependent Vasorelaxation by Targeting Endothelial Nitric Oxide Synthase. Hypertension 2012, 60, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-F.; Huang, J.; Li, H.; Zhang, C.; Huang, X.; Tong, G.; Xu, Y.-Z. MicroRNA-221 regulates endothelial nitric oxide production and inflammatory response by targeting adiponectin receptor 1. Gene 2015, 565, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, R.; Muthukumaravel, S.; Jambulingam, P. The Involvement of Neuroinflammation in Dengue Viral Disease: Importance of Innate and Adaptive Immunity. Neuroimmunomodulation 2019, 26, 111–118. [Google Scholar] [CrossRef]
- Tsai, T.-T.; Chen, C.-L.; Lin, Y.-S.; Chang, C.-P.; Tsai, C.-C.; Cheng, Y.-L.; Huang, C.-C.; Ho, C.-J.; Lee, Y.-C.; Lin, L.-T.; et al. Microglia retard dengue virus-induced acute viral encephalitis. Sci. Rep. 2016, 6, 27670. [Google Scholar] [CrossRef]
- Al-Shujairi, W.H.; Clarke, J.N.; Davies, L.T.; Alsharifi, M.; Pitson, S.; Carr, J. Intracranial Injection of Dengue Virus Induces Interferon Stimulated Genes and CD8+ T Cell Infiltration by Sphingosine Kinase 1 Independent Pathways. PLoS ONE 2017, 12, e0169814. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Maniakis-Grivas, G.; Singh, U.K.; Asher, R.M.; Mauri, F.; Elkington, P.; Friedland, J.S. Interleukin-17 regulates matrix metalloproteinase activity in human pulmonary tuberculosis. J. Pathol. 2018, 244, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A.L. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef]
- Liu, Y.-J. Dendritic Cell Subsets and Lineages, and Their Functions in Innate and Adaptive Immunity. Cell 2001, 106, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Cerny, D.; Haniffa, M.; Shin, A.; Bigliardi, P.L.; Tan, B.K.; Lee, B.; Poidinger, M.; Tan, E.Y.; Ginhoux, F.; Fink, K. Selective Susceptibility of Human Skin Antigen Presenting Cells to Productive Dengue Virus Infection. PLoS Pathog. 2014, 10, e1004548. [Google Scholar] [CrossRef] [Green Version]
- Shresta, S.; Kyle, J.L.; Snider, H.M.; Basavapatna, M.; Beatty, P.R.; Harris, E. Interferon-Dependent Immunity Is Essential for Resistance to Primary Dengue Virus Infection in Mice, Whereas T- and B-Cell-Dependent Immunity Are Less Critical. J. Virol. 2004, 78, 2701–2710. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, E.J.M.; Silva, A.M.; Cordeiro, M.T.; Brito, C.A.; Gil, L.H.V.G.; Braga-Neto, U.; Marques, E.T.A. Alternative Complement Pathway Deregulation Is Correlated with Dengue Severity. PLoS ONE 2009, 4, e6782. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [Green Version]
- Green, A.M.; Beatty, P.R.; Hadjilaou, A.; Harris, E. Innate Immunity to Dengue Virus Infection and Subversion of Antiviral Responses. J. Mol. Biol. 2014, 426, 1148–1160. [Google Scholar] [CrossRef] [Green Version]
- Nasirudeen, A.M.A.; Wong, H.H.; Thien, P.; Xu, S.; Lam, K.-P.; Liu, D.X. RIG-I, MDA5 and TLR3 Synergistically Play an Important Role in Restriction of Dengue Virus Infection. PLoS Neglected Trop. Dis. 2011, 5, e926. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; West, K.; Kalayanarooj, S.; Gibbons, R.V.; Srikiatkhachorn, A.; Green, S.; Libraty, D.; Jaiswal, S.; Rothman, A. B-Cell Responses During Primary and Secondary Dengue Virus Infections in Humans. J. Infect. Dis. 2011, 204, 1514–1522. [Google Scholar] [CrossRef]
- Ubol, S.; Phuklia, W.; Kalayanarooj, S.; Modhiran, N. Mechanisms of Immune Evasion Induced by a Complex of Dengue Virus and Preexisting Enhancing Antibodies. J. Infect. Dis. 2010, 201, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Sette, A.; Weiskopf, D. Cytotoxic CD4 T Cells: Differentiation, Function, and Application to Dengue Virus Infection. Front. Immunol. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Grifoni, A.; Sette, A.; Weiskopf, D. Human T Cell Response to Dengue Virus Infection. Front. Immunol. 2019, 10, 2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suharti, C.; Van Gorp, E.C.M.; Dolmans, W.M.V.; Setiati, E.T.; Hack, C.E.; Djokomoeljanto, R.; Van Der Meer, J.W.M. Cytokine patterns during dengue shock syndrome. Eur. Cytokine Netw. 2003, 14, 172–177. [Google Scholar] [PubMed]
- Chakravarti, A.; Kumaria, R. Circulating levels of tumour necrosis factor-alpha & interferon-gamma in patients with dengue & dengue haemorrhagic fever during an outbreak. Indian J. Med Res. 2006, 123, 25–30. [Google Scholar] [PubMed]
- Bozza, F.A.; Cruz, O.G.; Zagne, S.M.; Azeredo, E.L.; Nogueira, R.M.; Assis, E.F.; Bozza, P.T.; Kubelka, C.F. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 2008, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Srikiatkhachorn, A.; Mathew, A.; Rothman, A.L. Immune-mediated cytokine storm and its role in severe dengue. Semin. Immunopathol. 2017, 39, 563–574. [Google Scholar] [CrossRef]
- Tsai, T.-T.; Chuang, Y.-J.; Lin, Y.-S.; Wan, S.-W.; Chen, C.-L.; Lin, C.-F. An emerging role for the anti-inflammatory cytokine interleukin-10 in dengue virus infection. J. Biomed. Sci. 2013, 20, 40. [Google Scholar] [CrossRef] [Green Version]
- Abhishek, K.S.; Chakravarti, A.; Baveja, C.P.; Kumar, N.; Siddiqui, O.; Kumar, S. Association of interleukin-2, -4 and -10 with dengue severity. Indian J. Pathol. Microbiol. 2017, 60, 66–69. [Google Scholar] [CrossRef]
- Kuczera, D.; Assolini, J.P.; Tomiotto-Pellissier, F.; Pavanelli, W.R.; Silveira, G.F. Highlights for Dengue Immunopathogenesis: Antibody-Dependent Enhancement, Cytokine Storm, and Beyond. J. Interf. Cytokine Res. 2018, 38, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Nunez, N.V.A.; Hoornweg, T.E.; Van De Pol, D.P.I.; Sjollema, K.A.; Flipse, J.; Van Der Schaar, H.M.; Smit, J.M. How antibodies alter the cell entry pathway of dengue virus particles in macrophages. Sci. Rep. 2016, 6, 28768. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.R.; Ong, E.; Tan, H.C.; Zhang, S.L.; Zhang, Q.; Tang, K.F.; Kaliaperumal, N.; Lim, A.P.C.; Hibberd, M.; Chan, S.H.; et al. Leukocyte immunoglobulin-like receptor B1 is critical for antibody-dependent dengue. Proc. Natl. Acad. Sci. USA 2014, 111, 2722–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viktorovskaya, O.V.; Greco, T.M.; Cristea, I.M.; Thompson, S.R. Identification of RNA Binding Proteins Associated with Dengue Virus RNA in Infected Cells Reveals Temporally Distinct Host Factor Requirements. PLoS Neglected Trop. Dis. 2016, 10, e0004921. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.Y.Y.; Low, J.Z.H.; Gan, E.S.; Ong, E.Z.; Zhang, S.L.-X.; Tan, H.C.; Chai, X.; Ghosh, S.; Ooi, E.E.; Chan, K.R. Antibody-Dependent Dengue Virus Entry Modulates Cell Intrinsic Responses for Enhanced Infection. mSphere 2019, 4, e00528-19. [Google Scholar] [CrossRef] [Green Version]
- Narayan, R.; Tripathi, S. Intrinsic ADE: The Dark Side of Antibody Dependent Enhancement During Dengue Infection. Front. Cell. Infect. Microbiol. 2020, 10, 580096. [Google Scholar] [CrossRef]
- Kawagoe, T.; Takeuchi, O.; Takabatake, Y.; Kato, H.; Isaka, Y.; Tsujimura, T.; Akira, S. TANK is a negative regulator of Toll-like receptor signaling and is critical for the prevention of autoimmune nephritis. Nat. Immunol. 2009, 10, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Flipse, J.; Wilschut, J.; Smit, J.M. Molecular Mechanisms Involved in Antibody-Dependent Enhancement of Dengue Virus Infection in Humans. Traffic 2012, 14, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Malavige, G.; Huang, L.-C.; Salimi, M.; Gomes, L.; Jayaratne, S.D.; Ogg, G.S. Cellular and Cytokine Correlates of Severe Dengue Infection. PLoS ONE 2012, 7, e50387. [Google Scholar] [CrossRef] [Green Version]
- Flipse, J.; Diosa-Toro, M.; Hoornweg, T.E.; Van De Pol, D.P.I.; Urcuqui-Inchima, S.; Smit, J.M. Antibody-Dependent Enhancement of Dengue Virus Infection in Primary Human Macrophages; Balancing Higher Fusion against Antiviral Responses. Sci. Rep. 2016, 6, 29201. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Cheng, A.; Wang, M.; Yin, Z.; Jia, R. The Dual Regulation of Apoptosis by Flavivirus. Front. Microbiol. 2021, 12, 654494. [Google Scholar] [CrossRef]
- Netsawang, J.; Noisakran, S.; Puttikhunt, C.; Kasinrerk, W.; Wongwiwat, W.; Malasit, P.; Yenchitsomanus, P.-T.; Limjindaporn, T. Nuclear localization of dengue virus capsid protein is required for DAXX interaction and apoptosis. Virus Res. 2010, 147, 275–283. [Google Scholar] [CrossRef]
- Morchang, A.; Yasamut, U.; Netsawang, J.; Noisakran, S.; Wongwiwat, W.; Songprakhon, P.; Srisawat, C.; Puttikhunt, C.; Kasinrerk, W.; Malasit, P.; et al. Cell death gene expression profile: Role of RIPK2 in dengue virus-mediated apoptosis. Virus Res. 2011, 156, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Nasirudeen, A.; Wang, L.; Liu, D.X. Induction of p53-dependent and mitochondria-mediated cell death pathway by dengue virus infection of human and animal cells. Microbes Infect. 2008, 10, 1124–1132. [Google Scholar] [CrossRef]
- Long, X.; Li, Y.; Qi, Y.; Xu, J.; Wang, Z.; Zhang, X.; Zhang, D.; Zhang, L.; Huang, J. XAF1 contributes to dengue virus-induced apoptosis in vascular endothelial cells. FASEB J. 2013, 27, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- El-Bacha, T.; Midlej, V.D.V.P.; da Silva, A.P.P.; da Costa, L.S.; Benchimol, M.; Galina, A.; Da Poian, A. Mitochondrial and bioenergetic dysfunction in human hepatic cells infected with dengue 2 virus. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 1158–1166. [Google Scholar] [CrossRef] [Green Version]
- Torrentes-Carvalho, A.; Azeredo, E.L.; Reis, S.R.; Miranda, A.S.; Gandini, M.; Barbosa, L.S.; Kubelka, C.F. Dengue-2 infection and the induction of apoptosis in human primary monocytes. Mem. Inst. Oswaldo Cruz 2009, 104, 1091–1099. [Google Scholar] [CrossRef] [Green Version]
- Suwanmanee, S.; Luplertlop, N. Immunopathogenesis of Dengue Virus-Induced Redundant Cell Death: Apoptosis and Pyroptosis. Viral Immunol. 2017, 30, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Thepparit, C.; Khakpoor, A.; Khongwichit, S.; Wikan, N.; Fongsaran, C.; Chingsuwanrote, P.; Panraksa, P.; Smith, D.R. Dengue 2 infection of HepG2 liver cells results in endoplasmic reticulum stress and induction of multiple pathways of cell death. BMC Res. Notes 2013, 6, 372. [Google Scholar] [CrossRef] [Green Version]
- Netsawang, J.; Panaampon, J.; Khunchai, S.; Kooptiwut, S.; Nagila, A.; Puttikhunt, C.; Yenchitsomanus, P.-T.; Limjindaporn, T. Dengue virus disrupts Daxx and NF-κB interaction to induce CD137-mediated apoptosis. Biochem. Biophys. Res. Commun. 2014, 450, 1485–1491. [Google Scholar] [CrossRef]
- Xiao, C.; Ghosh, S. NF-κB, an Evolutionarily Conserved Mediator of Immune and Inflammatory Responses. Mech. Lymph. Act. Immune Regul. X 2005, 560, 41–45. [Google Scholar] [CrossRef]
- Olagnier, D.; Peri, S.; Steel, C.; Van Montfoort, N.; Chiang, C.; Beljanski, V.; Slifker, M.; He, Z.; Nichols, C.N.; Lin, R.; et al. Cellular Oxidative Stress Response Controls the Antiviral and Apoptotic Programs in Dengue Virus-Infected Dendritic Cells. PLoS Pathog. 2014, 10, e1004566. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-F.; Lei, H.-Y.; Shiau, A.-L.; Liu, H.-S.; Yeh, T.-M.; Chen, S.-H.; Liu, C.-C.; Chiu, S.-C.; Lin, Y.-S. Endothelial Cell Apoptosis Induced by Antibodies Against Dengue Virus Nonstructural Protein 1 Via Production of Nitric Oxide. J. Immunol. 2002, 169, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morchang, A.; Lee, R.C.H.; Yenchitsomanus, P.-T.; Sreekanth, G.P.; Noisakran, S.; Chu, J.J.H.; Limjindaporn, T. RNAi screen reveals a role of SPHK2 in dengue virus–mediated apoptosis in hepatic cell lines. PLoS ONE 2017, 12, e0188121. [Google Scholar] [CrossRef] [Green Version]
- Hart, J.R.; Vogt, P.K. Phosphorylation of AKT: A Mutational Analysis. Oncotarget 2011, 2, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Airo, A.M.; Urbanowski, M.D.; Lopez-Orozco, J.; You, J.H.; Skene-Arnold, T.D.; Holmes, C.; Yamshchikov, V.; Malik-Soni, N.; Frappier, L.; Hobman, T.C. Expression of flavivirus capsids enhance the cellular environment for viral replication by activating Akt-signalling pathways. Virology 2018, 516, 147–157. [Google Scholar] [CrossRef]
- Li, J.; Huang, R.; Liao, W.; Chen, Z.; Zhang, S.; Huang, R. Dengue virus utilizes calcium modulating cyclophilin-binding ligand to subvert apoptosis. Biochem. Biophys. Res. Commun. 2012, 418, 622–627. [Google Scholar] [CrossRef]
- Panyasrivanit, M.; Greenwood, M.; Murphy, D.; Isidoro, C.; Auewarakul, P.; Smith, D.R. Induced autophagy reduces virus output in dengue infected monocytic cells. Virology 2011, 418, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-R.; Lei, H.-Y.; Liu, M.-T.; Wang, J.-R.; Chen, S.-H.; Jiang-Shieh, Y.-F.; Lin, Y.-S.; Yeh, T.-M.; Liu, C.-C.; Liu, H.-S. Autophagic machinery activated by dengue virus enhances virus replication. Virology 2008, 374, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Margariti, A.; Li, H.; Chen, T.; Martin, D.; Vizcay-Barrena, G.; Alam, S.; Karamariti, E.; Xiao, Q.; Zampetaki, A.; Zhang, Z.; et al. XBP1 mRNA Splicing Triggers an Autophagic Response in Endothelial Cells through BECLIN-1 Transcriptional Activation. J. Biol. Chem. 2013, 288, 859–872. [Google Scholar] [CrossRef] [Green Version]
- Acharya, B.; Gyeltshen, S.; Chaijaroenkul, W.; Na-Bangchang, K. Significance of Autophagy in Dengue Virus Infection: A Brief Review. Am. J. Trop. Med. Hyg. 2019, 100, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Datan, E.; Roy, S.G.; Germain, G.; Zali, N.; McLean, E.J.; Golshan, G.; Harbajan, S.; Lockshin, A.R.; Zakeri, Z. Dengue-induced autophagy, virus replication and protection from cell death require ER stress (PERK) pathway activation. Cell Death Dis. 2016, 7, e2127. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-R.; Kuo, S.-H.; Lin, C.-Y.; Fu, P.-J.; Lin, Y.-S.; Yeh, T.-M.; Liu, H.-S. Dengue virus-induced ER stress is required for autophagy activation, viral replication, and pathogenesis both in vitro and in vivo. Sci. Rep. 2018, 8, 489. [Google Scholar] [CrossRef] [PubMed]
- Metz, P.; Chiramel, A.; Chatel-Chaix, L.; Alvisi, G.; Bankhead, P.; Mora-Rodríguez, R.; Long, G.; Hamacher-Brady, A.; Brady, N.R.; Bartenschlager, R. Dengue Virus Inhibition of Autophagic Flux and Dependency of Viral Replication on Proteasomal Degradation of the Autophagy Receptor p62. J. Virol. 2015, 89, 8026–8041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.; Rozera, R.; Kumar, R.; Haque, A.; Attri, A. Herbal remedies, vaccines and drugs for dengue fever: Emerging prevention and treatment strategies. Asian Pac. J. Trop. Med. 2019, 12, 147. [Google Scholar] [CrossRef]
- Ali, F.; Chorsiya, A.; Anjum, V.; Khasimbi, S.; Ali, A. A systematic review on phytochemicals for the treatment of dengue. Phytotherapy Res. 2020, 35, 1782–1816. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Yoon, I.-K. A review of Dengvaxia®: Development to deployment. Hum. Vaccines Immunother. 2019, 15, 2295–2314. [Google Scholar] [CrossRef] [Green Version]
- Martinez, D.R.; Metz, S.W.; Baric, R.S. Dengue Vaccines: The Promise and Pitfalls of Antibody-Mediated Protection. Cell Host Microbe 2021, 29, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Nivarthi, U.K.; Swanstrom, J.; Delacruz, M.J.; Patel, B.; Durbin, A.P.; Whitehead, S.S.; Kirkpatrick, B.D.; Pierce, K.K.; Diehl, S.A.; Katzelnick, L.; et al. A tetravalent live attenuated dengue virus vaccine stimulates balanced immunity to multiple serotypes in humans. Nat. Commun. 2021, 12, 1102. [Google Scholar] [CrossRef] [PubMed]
- Wollner, C.J.; Richner, M.; Hassert, M.A.; Pinto, A.K.; Brien, J.D.; Richner, J.M. A Dengue Virus Serotype 1 mRNA-LNP Vaccine Elicits Protective Immune Responses. J. Virol. 2021, 95, e02482-20. [Google Scholar] [CrossRef]
- Idrees, S.; Ashfaq, U.A. RNAi: Antiviral therapy against dengue virus. Asian Pac. J. Trop. Biomed. 2013, 3, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.A.A.; Palanisamy, U.D.; Khoo, J.J.; Dhanoa, A.; Hassan, S.S. Efficacy of geraniin on dengue virus type-2 infected BALB/c mice. Virol. J. 2019, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Jasso-Miranda, C.; Herrera-Camacho, I.; Flores-Mendoza, L.K.; Dominguez, F.; Vallejo-Ruiz, V.; Sanchez-Burgos, G.G.; Pando-Robles, V.; Santos-Lopez, G.; Reyes-Leyva, J. Antiviral and immunomodulatory effects of polyphenols on macrophages infected with dengue virus serotypes 2 and 3 enhanced or not with antibodies. Infect. Drug Resist. 2019, 12, 1833–1852. [Google Scholar] [CrossRef]
- Islam, M.T.; Mubarak, M.S. Diterpenes and their derivatives as promising agents against dengue virus and dengue vectors: A literature-based review. Phytotherapy Res. 2019, 34, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Shen, L.; Benyoucef, S.; De Groote, D.; Deubel, V.; Wattré, P. Enhanced TNFα production by monocytic-like cells exposed to dengue virus antigens. Immunol. Lett. 1996, 53, 115–120. [Google Scholar] [CrossRef]
- Keivan, Z.; Boon-Teong, T.; Sing-Sin, S.; Pooi-Fong, W.; Mohd, R.M.; Sazaly, A.; Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; et al. In vitro antiviral activity of fisetin, rutin and naringenin against dengue virus type-2. J. Med. Plants Res. 2014, 8, 307–312. [Google Scholar] [CrossRef]
- Trujillo-Correa, A.I.; Quintero-Gil, D.C.; Diaz-Castillo, F.; Quiñones, W.; Robledo, S.M.; Martinez-Gutierrez, M. In vitro and in silico anti-dengue activity of compounds obtained from Psidium guajava through bioprospecting. BMC Complement. Altern. Med. 2019, 19, 298–316. [Google Scholar] [CrossRef]
- Raekiansyah, M.; Buerano, C.C.; Luz, M.A.D.; Morita, K. Inhibitory effect of the green tea molecule EGCG against dengue virus infection. Arch. Virol. 2018, 163, 1649–1655. [Google Scholar] [CrossRef]
- Loaiza-Cano, V.; Monsalve-Escudero, L.M.; Filho, C.d.S.M.B.; Martinez-Gutierrez, M.; de Sousa, D.P. Antiviral Role of Phenolic Compounds against Dengue Virus: A Review. Biomolecules 2020, 11, 11. [Google Scholar] [CrossRef]
- Han, Y.-S.; Penthala, N.R.; Oliveira, M.; Mesplède, T.; Xu, H.; Quan, Y.; Crooks, P.A.; Wainberg, M.A. Identification of resveratrol analogs as potent anti-dengue agents using a cell-based assay. J. Med Virol. 2017, 89, 397–407. [Google Scholar] [CrossRef]
- Soto-Acosta, R.; Bautista-Carbajal, P.; Syed, G.H.; Siddiqui, A.; Del Angel, R.M. Nordihydroguaiaretic acid (NDGA) inhibits replication and viral morphogenesis of dengue virus. Antivir. Res. 2014, 109, 132–140. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Chen, D.-Y.; Wen, H.-W.; Ou, J.-L.; Chiou, S.-S.; Chen, J.-M.; Wong, M.-L.; Hsu, W.-L. Inhibition of Enveloped Viruses Infectivity by Curcumin. PLoS ONE 2013, 8, e62482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; Mishra, K.P.; Ganju, L. Salidroside exhibits anti-dengue virus activity by upregulating host innate immune factors. Arch. Virol. 2016, 161, 3331–3344. [Google Scholar] [CrossRef]
- Wu, D.-W.; Mao, F.; Ye, Y.; Li, J.; Xu, C.-L.; Luo, X.-M.; Chen, J.; Shen, X. Policresulen, a novel NS2B/NS3 protease inhibitor, effectively inhibits the replication of DENV2 virus in BHK-21 cells. Acta Pharmacol. Sin. 2015, 36, 1126–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, C.-Y.; Chen, S.-J.; Wu, H.-N.; Ping, Y.-H.; Lin, C.-Y.; Shiuan, D.; Chen, C.-L.; Lee, Y.-R.; Huang, K.-J. Honokiol, a Lignan Biphenol Derived from the Magnolia Tree, Inhibits Dengue Virus Type 2 Infection. Viruses 2015, 7, 4894–4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.H.; Si, J.; Kang, C.; Chung, B.; Chung, D.; Kim, D. Facile preparation of water soluble curcuminoids extracted from turmeric (Curcuma longa L.) powder by using steviol glucosides. Food Chem. 2017, 214, 366–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.P.; Houston, S.D.; Modhiran, N.; Savchenko, A.I.; Boyle, G.M.; Young, P.R.; Watterson, D.; Williams, C.M. Stachyonic Acid: A Dengue Virus Inhibitor from Basilicum polystachyon. Chem. Eur. J. 2019, 25, 5664–5667. [Google Scholar] [CrossRef]
- Peng, M.; Swarbrick, C.M.D.; Chan, K.W.-K.; Luo, D.; Zhang, W.; Lai, X.; Li, G.; Vasudevan, S.G. Luteolin escape mutants of dengue virus map to prM and NS2B and reveal viral plasticity during maturation. Antivir. Res. 2018, 154, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.M.; Chieng, J.Y.; Pan, Y. Recent insights on anti-dengue virus (DENV) medicinal plants: Review on in vitro, in vivo and in silico discoveries. All Life 2021, 14, 1–33. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanaware, N.; Banerjee, A.; Mullick Bagchi, S.; Bagchi, P.; Mukherjee, A. Dengue Virus Infection: A Tale of Viral Exploitations and Host Responses. Viruses 2021, 13, 1967. https://doi.org/10.3390/v13101967
Nanaware N, Banerjee A, Mullick Bagchi S, Bagchi P, Mukherjee A. Dengue Virus Infection: A Tale of Viral Exploitations and Host Responses. Viruses. 2021; 13(10):1967. https://doi.org/10.3390/v13101967
Chicago/Turabian StyleNanaware, Nikita, Anwesha Banerjee, Satarupa Mullick Bagchi, Parikshit Bagchi, and Anupam Mukherjee. 2021. "Dengue Virus Infection: A Tale of Viral Exploitations and Host Responses" Viruses 13, no. 10: 1967. https://doi.org/10.3390/v13101967