
Dynamics of SARS-CoV-2 Variants of Concern in Vaccination Model City in the State of Sao Paulo, Brazil
 , , , , , , , ,
, , , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement and Study Location
2.2. Molecular Confirmation of SARS-CoV-2 Infection
2.3. SARS-CoV-2 Sequencing
2.4. Bioinformatics Pipeline
2.5. Phylogenetics Analysis
2.6. Statistical Analysis
3. Results
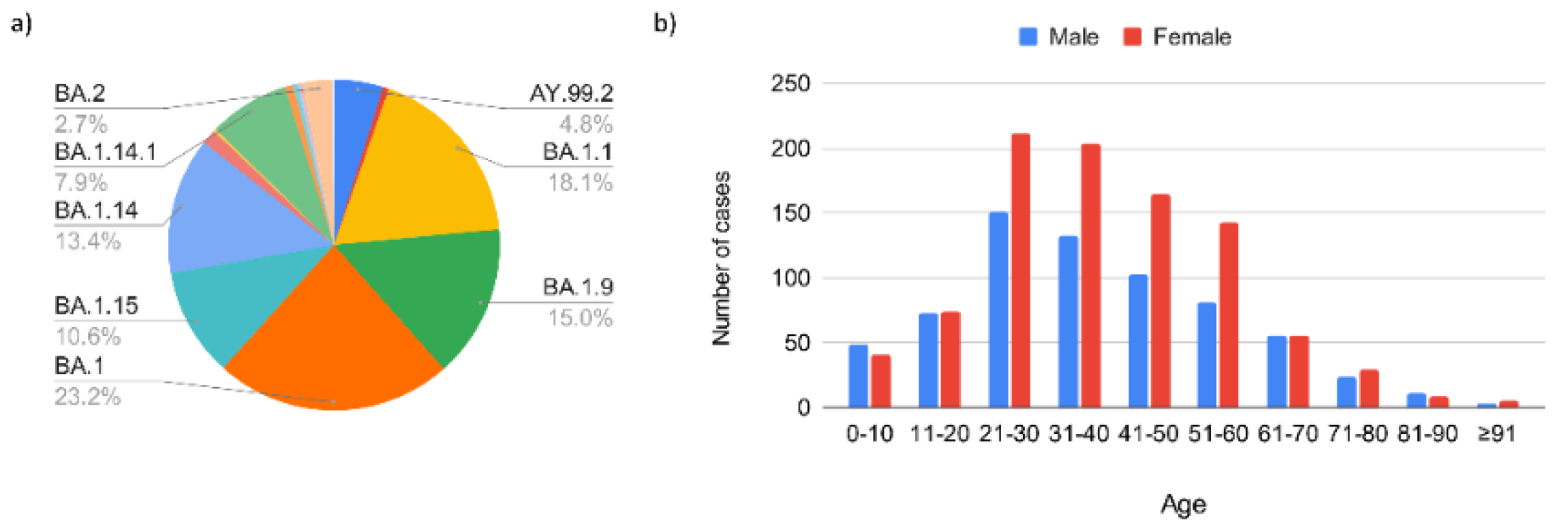
3.1. Aspects of the Tested Population and Performed Sequencing
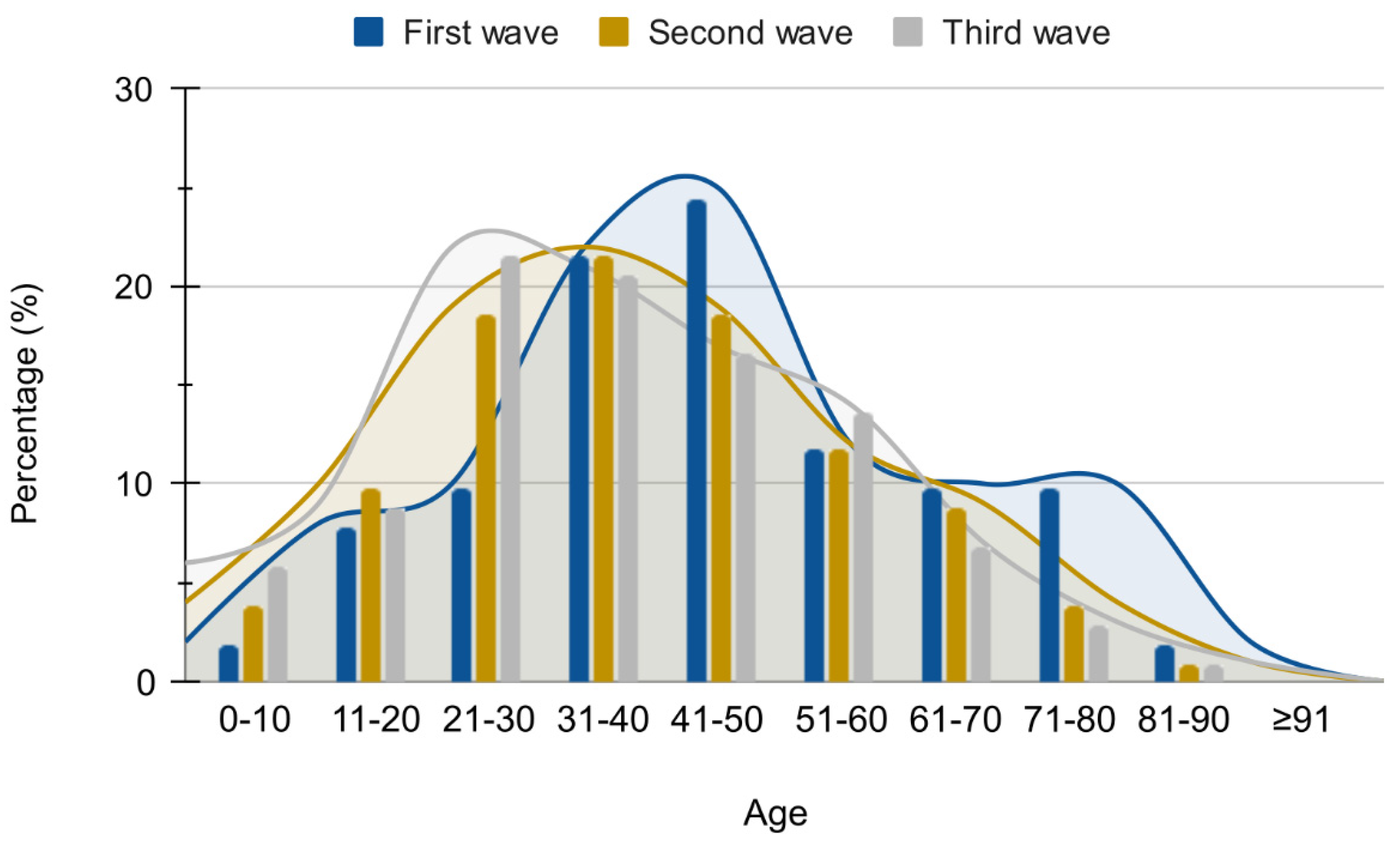
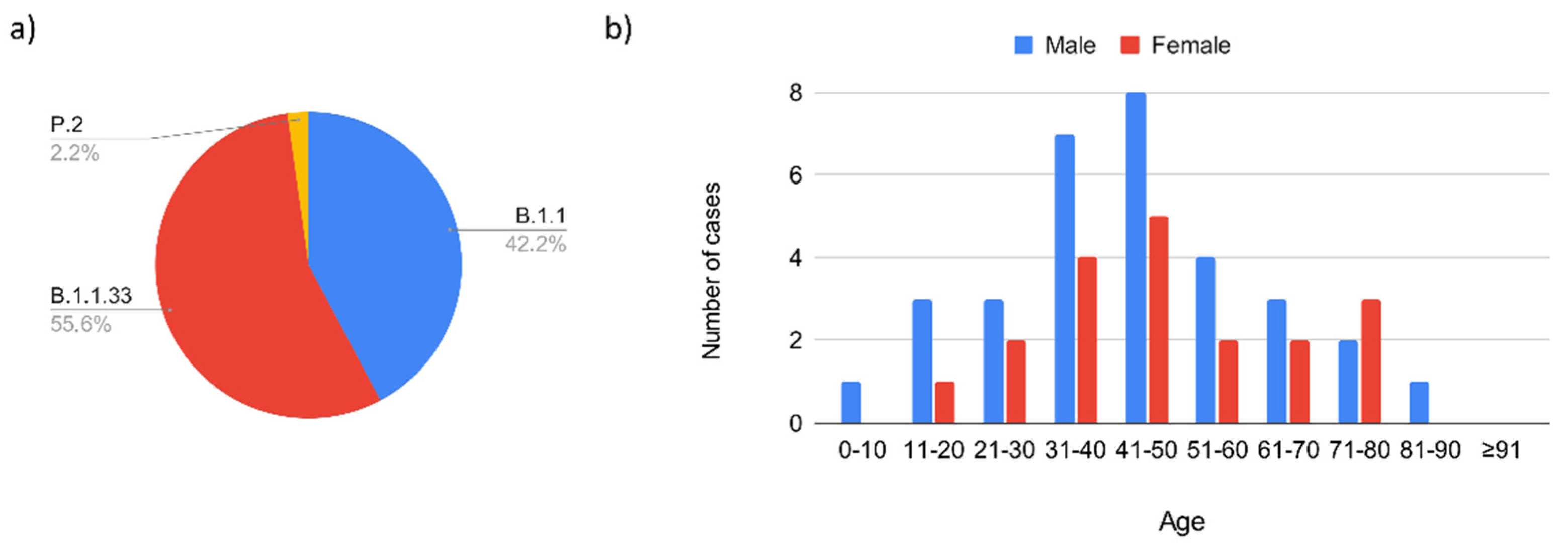
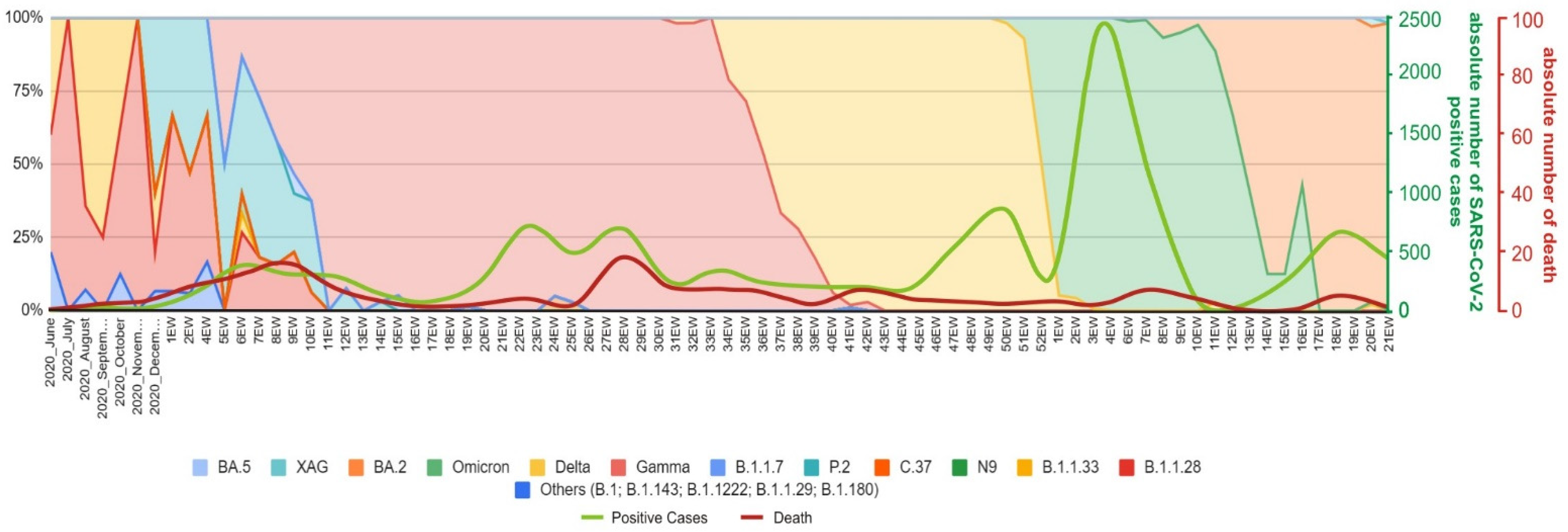
3.1.1. Introduction of SARS-CoV-2 in Serrana and First Epidemic Wave (February 2020–November 2020)
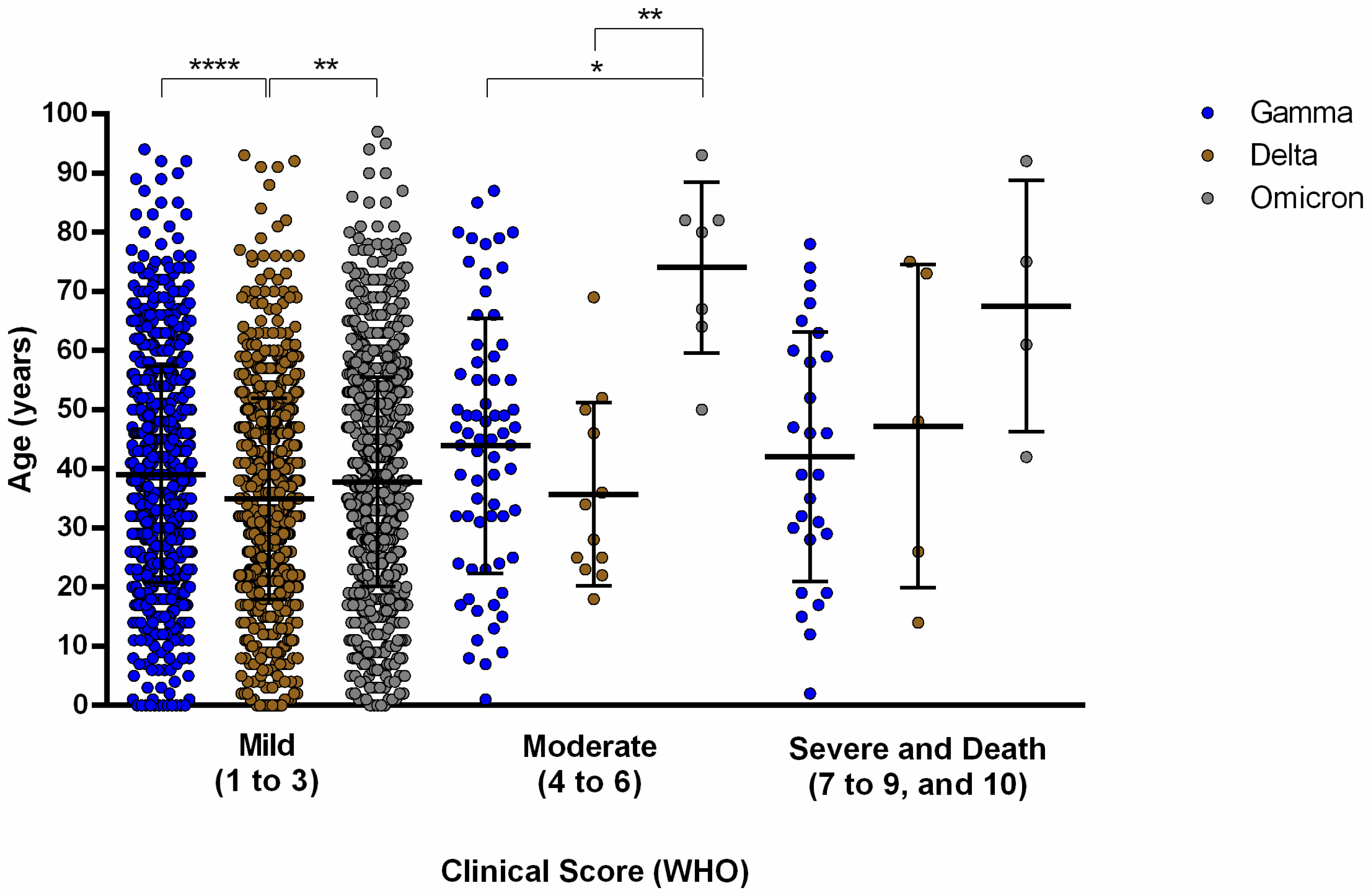
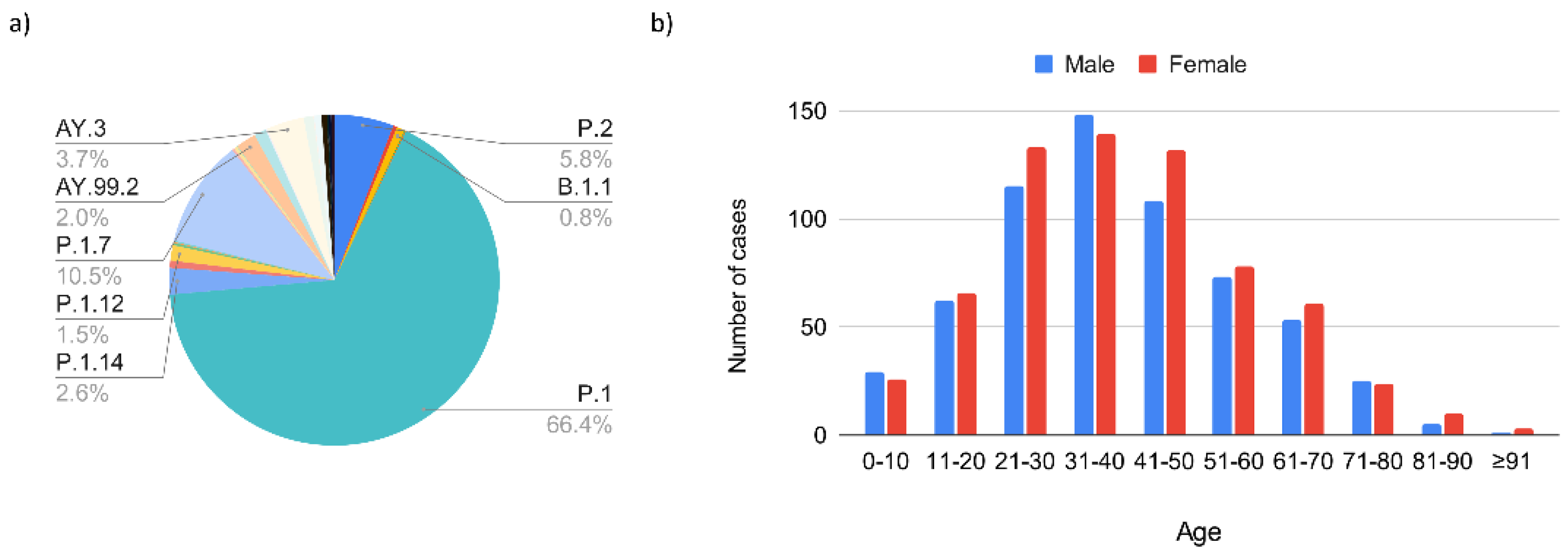
3.1.2. The Second Epidemic Wave (November 2020–October 2021) Related to the Gamma and Delta Variants of Concern
3.1.3. Third Epidemic Wave (December 2021–April 2022)
3.2. Global SARS-CoV-2 Phylogenetic Analysis in the Town of Serrana
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coronavirus Brasil. Available online: https://covid.saude.gov.br/ (accessed on 29 July 2022).
- Naveca, F.G.; Nascimento, V. COVID-19 in Amazonas, Brazil, was driven by the persistence of endemic lineages and P.1 emergence. Nat. Med. 2021, 7, 1230–1238. [Google Scholar] [CrossRef]
- Carvalho, M.S.; Bastos, L.S. Incidence of SARS-CoV-2 over four epidemic waves in a low-resource community in Rio de Janeiro, Brazil: A prospective cohort study. Lancet Reg. Health Am. 2022, 12, 100283. [Google Scholar] [CrossRef] [PubMed]
- Moutinho, S. Brazilian Town Experiment Shows Mass Vaccination Can Wipe Out COVID-19. Available online: https://www.science.org/content/article/brazilian-town-experiment-shows-mass-vaccination-can-wipe-out-covid-19 (accessed on 1 June 2021).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 29 July 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Narasimhan, V.; Danecek, P.; Scally, A.; Xue, Y.; Tyler-Smith, C.; Durbin, R. BCFtools/RoH: A Hidden Markov Model Approach for Detecting Autozygosity from next-Generation Sequencing Data. Bioinformatics 2016, 32, 1749–1751. [Google Scholar] [CrossRef]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, 0163962. [Google Scholar] [CrossRef]
- Aksamentov, I.; Roemer, C. Nextclade: Clade assignment, mutation calling and quality control for viral genomes. JOSS 2021, 67, 3373–3379. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. MEE 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Boletim Epidemiológico de Serrana 17. 2020. Available online: http://www.serrana.sp.gov.br/media/uploads/be-12-4-22.pdf (accessed on 29 July 2022).
- Jesus, J.G.; Sacchi, C. Importation and early local transmission of COVID-19 in Brazil, 2020. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Jernigan, D.B.; 2019-nCoV CDC Response Team. Initial Public Health Response and Interim Clinical Guidance for the 2019 Novel Coronavirus Outbreak—United States, 31 December 2019–4 February 2020. MMWR Morb. Mortal Wkly Rep. 2020, 69, 140–146, Erratum in MMWR Morb. Mortal Wkly Rep. 2020, 69, 173. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.N.; Garibaldi, P.M.M.; Moraes, G.R.; Moura, J.C.; Klein, T.M.; Machado, L.E.; Scofoni, L.F.B.; Haddad, S.K.; Calado, R.T.; Covas, D.T.; et al. The impact of an enhanced health surveillance system for COVID-19 management in Serrana, Brazil. Public Health Pr. 2022, 4, 100301. [Google Scholar] [CrossRef]
- Slavov, S.N.; Giovanetti, M. Molecular surveillance of the on-going SARS-COV-2 epidemic in Ribeirao Preto City, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, 104976. [Google Scholar] [CrossRef]
- Giovanetti, M.; Slavov, S.N. Genomic epidemiology of the SARS-CoV-2 epidemic in Brazil. Nat. Microbiol. 2022, 7, 1490–1500. [Google Scholar] [CrossRef]
- Alcantara, L.C.J.; Nogueira, E. SARS-CoV-2 epidemic in Brazil: How the displacement of variants has driven distinct epidemic waves. Virus Res. 2022, 315, 198785. [Google Scholar] [CrossRef]
- Kashima, S.; Slavov, S.N. Introduction of SARS-CoV-2 C.37 (WHO VOI lambda) in the Sao Paulo State, Southeast Brazil. J. Med. Virol. 2021, 94, 1206–1211. [Google Scholar] [CrossRef]
- Arantes, I.G.; Salvato, R.S. Multiple Introductions of SARS-CoV-2 C.37 Lambda lineage in the Southern Brazilian region. J. Travel Med. 2021, 28, taab153. [Google Scholar] [CrossRef] [PubMed]
- Banho, C.A.; Sacchetto, L. Impact of SARS-CoV-2 Gamma lineage introduction and COVID-19 vaccination on the epidemiological landscape of a Brazilian city. Commun. Med. 2022, 41, 41. [Google Scholar] [CrossRef] [PubMed]
- Basso, P.; Negro, C.; Cegolon, L.; Larese Filon, F. Risk of Vaccine Breakthrough SARS-CoV-2 Infection and Associated Factors in Healthcare Workers of Trieste Teaching Hospitals (North-Eastern Italy). Viruses 2022, 14, 336. [Google Scholar] [CrossRef]
- Ai, J.; Wang, X. Antibody evasion of SARS-CoV-2 Omicron BA.1, BA.1.1, BA.2, and BA.3 sub-lineages. Cell Host Microb. 2022, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Mohkhedkar, M. Mutations in spike protein T cell epitopes of SARS-CoV-2 variants: Plausible influence on vaccine efficacy. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 9, 166432. [Google Scholar] [CrossRef]
- Yan, L.N.; Zhao, Z.X. Neutralizing antibodies and cellular immune response after two doses of inactivated SARS-CoV-2 vaccine in China. Expert Rev. Vaccines 2022, 29, 1–9. [Google Scholar] [CrossRef]
- Cegolon, L.; Ronchese, F.; Ricci, F.; Negro, C.; Larese-Filon, F. SARS-CoV-2 Infection in Health Care Workers of Trieste (North-Eastern Italy), 1 October 2020–7 February 2022: Occupational Risk and the Impact of the Omicron Variant. Viruses 2022, 14, 1663. [Google Scholar] [CrossRef]
- Madhi, S.A.; Kwatra, G. Population Immunity and COVID-19 Severity with Omicron Variant in South Africa. N. Engl. J. Med. 2022, 386, 1314–1326. [Google Scholar] [CrossRef]
- Lewnard, J.A.; Hong, V.X. Clinical outcomes associated with SARS-CoV-2 Omicron (B.1.1.529) variant and BA.1/BA.1.1 or BA.2 subvariant infection in southern California. Nat. Med. 2022. accepted. [Google Scholar] [CrossRef]
- Elliott, P.; Eales, O.; Bodinier, B.; Tang, D.; Wang, H.; Jonnerby, J.; Haw, D.; Elliott, J.; Whitaker, M.; Walters, C.E.; et al. Dynamics of a national Omicron SARS-CoV-2 epidemic during January 2022 in England. Nat. Commun. 2022, 13, 4500. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slavov, S.N.; de La-Roque, D.G.L.; da Costa, P.N.M.; Rodrigues, E.S.; Santos, E.V.; Borges, J.S.; Evaristo, M.; de Matos Maçonetto, J.; Marques, A.A.; Milhomens, J.; et al. Dynamics of SARS-CoV-2 Variants of Concern in Vaccination Model City in the State of Sao Paulo, Brazil. Viruses 2022, 14, 2148. https://doi.org/10.3390/v14102148
Slavov SN, de La-Roque DGL, da Costa PNM, Rodrigues ES, Santos EV, Borges JS, Evaristo M, de Matos Maçonetto J, Marques AA, Milhomens J, et al. Dynamics of SARS-CoV-2 Variants of Concern in Vaccination Model City in the State of Sao Paulo, Brazil. Viruses. 2022; 14(10):2148. https://doi.org/10.3390/v14102148
Chicago/Turabian StyleSlavov, Svetoslav Nanev, Debora Glenda Lima de La-Roque, Pericles Natan Mendes da Costa, Evandra Strazza Rodrigues, Elaine Vieira Santos, Josiane Serrano Borges, Mariane Evaristo, Juliana de Matos Maçonetto, Adriana Aparecida Marques, Jonathan Milhomens, and et al. 2022. "Dynamics of SARS-CoV-2 Variants of Concern in Vaccination Model City in the State of Sao Paulo, Brazil" Viruses 14, no. 10: 2148. https://doi.org/10.3390/v14102148