
Co-Circulation of West Nile, Usutu, and Tick-Borne Encephalitis Viruses in the Same Area: A Great Challenge for Diagnostic and Blood and Organ Safety
 , , ,
, , ,
Abstract
:1. The Changing World of Flaviviruses and Their Vectors
2. Superior Criteria for Public Health Departments, Transplantation, and Blood Establishments in the European Union
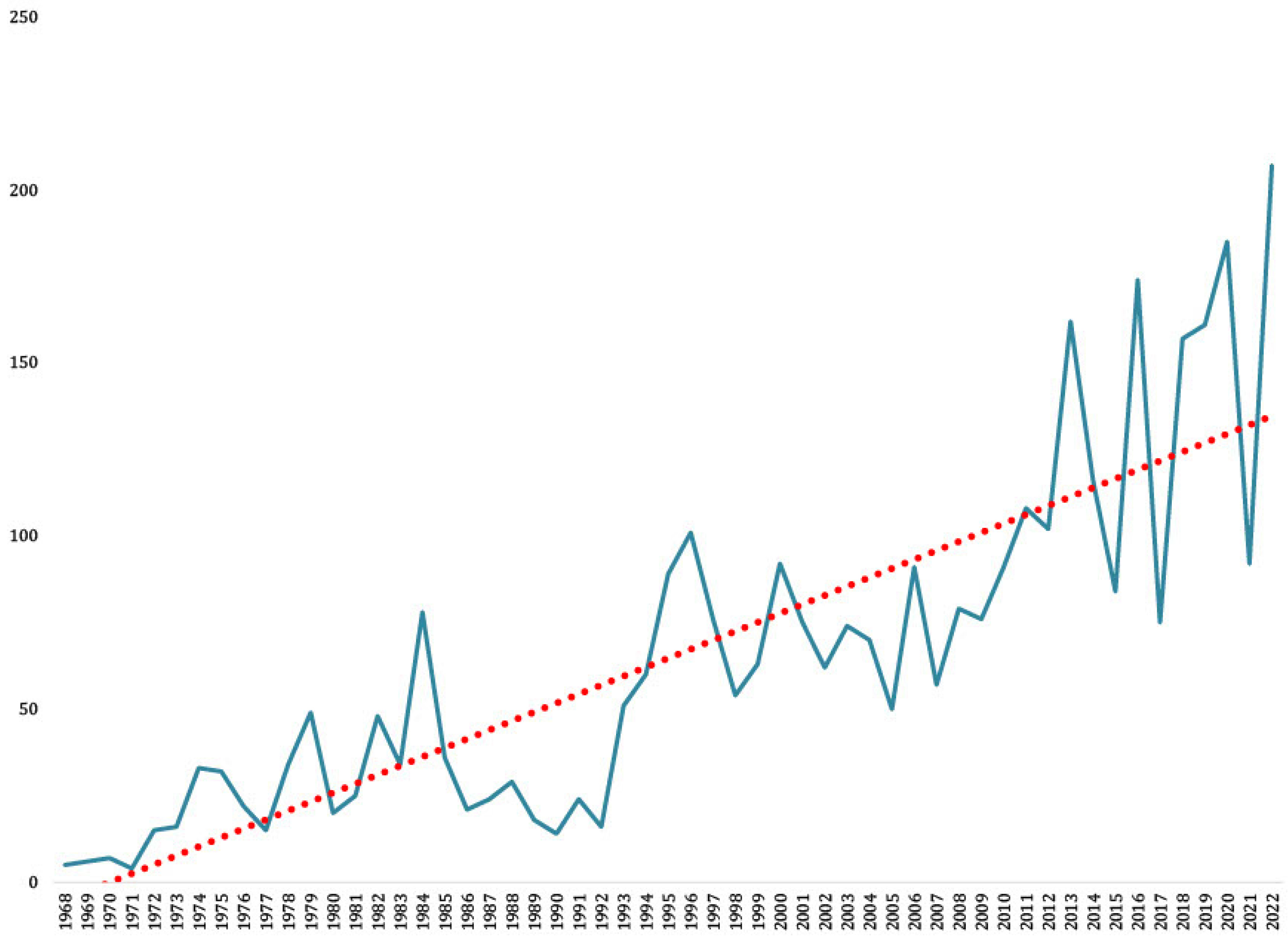
3. Overlapping Area of Co-Circulating WNV, USUV, and TBEV
4. Nascent Challenges for Clinical and Laboratory Diagnostics
5. Are Blood Components and Transplanted Organs Still Safe?
6. The Call for the Novelty to Protect Human Health
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Wondim, M.A.; Czupryna, P.; Pancewicz, S.; Kruszewska, E.; Groth, M.; Moniuszko-Malinowska, A. Epidemiological Trends of Trans-Boundary Tick-Borne Encephalitis in Europe, 2000–2019. Pathogens 2022, 11, 704. [Google Scholar] [CrossRef]
- Cunze, S.; Glock, G.; Kochmann, J.; Klimpel, S. Ticks on the move—Climate change-induced range shifts of three tick species in Europe: Current and future habitat suitability for Ixodes ricinus in comparison with Dermacentor reticulatus and Dermacentor marginatus. Parasitol. Res. 2021, 121, 2241–2252. [Google Scholar] [CrossRef]
- Bødker, R.; Klitgård, K.; Byriel, D.B.; Kristensen, B. Establishment of the West Nile virus vector, Culex modestus, in a residential area in Denmark. J. Vector Ecol. 2014, 39, 1–3. [Google Scholar] [CrossRef]
- Lindström, A.; Lilja, T. First finding of the West Nile virus vector Culex modestus Ficalbi 1889 (Diptera; Culicidae) in Sweden. J. Eur. Mosq. Control Assoc. 2018, 36, 1–2. [Google Scholar]
- Haba, Y.; McBride, L. Origin and status of Culex pipiens mosquito ecotypes. Curr. Biol. 2022, 32, R237–R246. [Google Scholar] [CrossRef]
- Čabanová, V.; Boršová, K.; Svitok, M.; Oboňa, J.; Svitková, I.; Barbušinová, E.; Derka, T.; Sláviková, M.; Klempa, B. An unwanted companion reaches the country: The first record of the alien mosquito Aedes japonicus japonicus (Theobald, 1901) in Slovakia. Parasites Vectors 2021, 14, 572. [Google Scholar] [CrossRef]
- Schaffner, F.; Medlock, J.M.; Van Bortel, W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. 2013, 19, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Lindh, E.; Argentini, C.; Remoli, M.E.; Fortuna, C.; Faggioni, G.; Benedetti, E.; Amendola, A.; Marsili, G.; Lista, F.; Rezza, G.; et al. The Italian 2017 Outbreak Chikungunya Virus Belongs to an Emerging Aedes albopictus-Adapted Virus Cluster Introduced from the Indian Subcontinent. Open Forum Infect. Dis. 2018, 12, ofy321. [Google Scholar] [CrossRef]
- Roelandt, S.; D´hondt, B. Alien Species in Belgium: A Fact Sheet for Rhipicephalus sanguineus. Available online: https://ias.biodiversity.be/documents/AA%20-%20Rhipicephalus%20(AA).pdf (accessed on 4 November 2022).
- Hornok, S.; Sándor, A.D.; Tomanović, S.; Beck, R.; D´Amico, G.; Kontschán, J.; Takács, N.; Görföl, T.; Bendjeddou, M.L.; Földvári, G.; et al. East and west separation of Rhipicephalus sanguineus mitochondrial lineages in the Mediterranean Basin. Parasites Vectors 2017, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Pascoe, E.L.; Nava, S.; Labruna, M.B.; Paddock, C.D.; Levin, M.L.; Marcantonio, M.; Foley, J.E. Predicting the northward expansion of tropical lineage Rhipicephalus sanguineus sensu lato ticks in the United States and its implications for medical and veterinary health. PLoS ONE 2022, 17, e0271683. [Google Scholar] [CrossRef]
- Perveen, N.; Khan, G. Crimean-Congo hemorrhagic fever in the Arab world: A systematic review. Front. Vet. Sci. 2022, 9, 938601. [Google Scholar] [CrossRef]
- Lindgren, E.; Gustafson, R. Tick-borne encephalitis in Sweden and climate change. Lancet 2001, 358, 16–18. [Google Scholar] [CrossRef]
- Biernat, B.; Karbowiak, G.; Werszko, J.; Stańczak, J. Prevalence of tick-borne encephalitis virus (TBEV) RNA in Dermacentor reticulatus ticks from natural and urban environment, Poland. Exp. Appl. Acarol. 2014, 64, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Commission Implementing Decision (EU) 2018/945 of 22 June 2018 on the Communicable Diseases and Related Special Health Issues to be Covered by Epidemiological Surveillance as Well as Relevant Case Definitions. Available online: https://eur-lex.europa.eu/legalcontent/EN/TXT/PDF/?uri=CELEX:32018D0945 (accessed on 12 December 2022).
- Directive 2002/98/EC of the European Parliament and of the Council of 27 January 2003 Setting Standards of Quality and Safety for the Collection, Testing, Processing, Storage and Distribution of Human Blood and Blood Components and Amending Directive 2001/83/EC. Available online: https://www.ema.europa.eu/en/documents/regulatory-procedural-guideline/directive-2002/98/ec-european-parliament-council-27-january-2003-setting-standards-quality-safety-collection-testing_en.pdf (accessed on 12 December 2022).
- Commission Directive 2004/33/EC of 22 March 2004 Implementing Directive 2002/98/EC of the European Parliament and of the Council as Regards Certain Technical Requirements for Blood and Blood Components. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2004:091:0025:0039:EN:PDF (accessed on 12 December 2022).
- Commission Directive 2014/110/EU of 17 December 2014 Amending Directive 2004/33/EC as Regards Temporary Deferral Criteria for Donors of Allogeneic Blood Donations. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32014L0110 (accessed on 12 December 2022).
- Directive 2010/45/EU of the European Parliament and of the Council of 7 July 2010 on Standards of Quality and Safety of Human Organs Intended for Transplantation. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32010L0053 (accessed on 12 December 2022).
- Čabanová, V.; Tichá, E.; Bradbury, R.S.; Zubriková, D.; Valentová, D.; Chovancová, G.; Grešáková, Ľ.; Víchová, B.; Šikutová, S.; Csank, T.; et al. Mosquito surveillance of West Nile and Usutu viruses in four territorial units of Slovakia and description of a confirmed autochthonous human case of West Nile fever, 2018 to 2019. Euro Surveill. 2021, 26, 2000063. [Google Scholar] [CrossRef]
- Ličková, M.; Vichová, B.; Derdáková, M.; Sláviková, M.; Fumačová Havlíková, S.; Zubríková, D.; Seleymová, D.; Chvostáč, M.; Blaňarová, L.; Kerlik, J.; et al. An integrated approach involving seroprevalence in farm animals and virus detection in collected ticks as an effective tool for tick-borne encephalitis virus surveillance in Slovakia. In Proceedings of the 16th International Conference on Lyme Borreliosis and Other Tick-Borne Diseases, Amsterdam, Netherlands, 4–7 September 2022. [Google Scholar]
- The Epidemic Intelligence Information System (EPIS). Available online: https://www.ecdc.europa.eu/en/publications-data/epidemic-intelligence-information-system-epis (accessed on 5 December 2022).
- Šumilo, D.; Bormane, A.; Asokliene, L.; Vasilenko, V.; Golovljova, I.; Avsic-Zupanc, T.; Hubalek, Z.; Randolph, S.E. Socio-economic factors in the differential upsurge of tick-borne encephalitis in Central and Eastern Europe. Rev. Med. Virol. 2008, 18, 81–95. [Google Scholar] [CrossRef]
- Růžek, D.; Županc, T.A.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A. Tick-Borne Encephalitis in Europe and Russia: Review of Pathogenesis, Clinical Features, Therapy, and Vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef]
- Kerlik, J.; Avdičová, M.; Štefkovičová, M.; Tarkovská, V.; Valachová, M.P.; Molčányi, T.; Mezencev, R. Slovakia Reports Highest Occurrence of Alimentary Tick-Borne Encephalitis in Europe: Analysis of Tick-Borne Encephalitis Outbreaks in Slovakia during 2007–2016. Travel Med. Infect. Dis. 2018, 26, 37–42. [Google Scholar] [CrossRef]
- Ličková, M.; Fumačová Havlíková, S.; Sláviková, M.; Klempa, B. Alimentary Infections by Tick-Borne Encephalitis Virus. Viruses 2022, 14, 56. [Google Scholar] [CrossRef]
- Korytár, Ľ.; Peňazziová, K.; Pistl, J.; Tichá, E.; Čabanová, V.; Csank, T. Retrospective review and current knowledge on the occurrence of West Nile virus in mosquito vectors, reservoirs and hosts in Slovakia (Central Europe). Acta Virol. 2020, 64, 187–200. [Google Scholar] [CrossRef]
- Kennedy, P.G.E.; Quan, P.L.; Lipkin, W.I. Viral Encephalitis of Unknown Cause: Current Perspective and Recent Advances. Viruses 2017, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Kemmerly, S.A. Diagnosis and treatment of West Nile infections. Ochsner J. 2003, 3, 16–17. [Google Scholar]
- Gdoura, M.; Fares, W.; Bougatef, S.; Inoubli, A.; Touzi, H.; Hogga, N.; Ben Dhifallah, I.; Hannachi, N.; Argoubi, A.; Kacem, S.; et al. The value of West Nile virus RNA detection by real-time RT-PCR in urine samples from patients with neuroinvasive forms. Arch. Microbiol. 2022, 204, 238. [Google Scholar] [CrossRef]
- Veje, M.; Studahl, M.; Norberg, P.; Roth, A.; Möbius, U.; Brink, M.; Bergström, T. Detection of Tick-borne encephalitis virus RNA in urine. J. Clin. Microbiol. 2014, 52, 4111–4112. [Google Scholar] [CrossRef] [Green Version]
- Barzon, L.; Pacenti, M.; Franchin, E.; Squarzon, L.; Sinigaglia, A.; Ulbert, S.; Cusinato, R.; Palù, G. Isolation of West Nile virus from urine samples of patients with acute infection. J. Clin. Microbiol. 2014, 52, 3411–3413. [Google Scholar] [CrossRef] [Green Version]
- Lustig, Y.; Sofer, D.; Bucris, E.D.; Mendelson, E. Surveillance and diagnosis of West Nile virus in the face of flavivirus cross-reactivity. Front. Microbiol. 2018, 9, 2421. [Google Scholar] [CrossRef]
- Pan American Health Organization/World Health Organization. Laboratory Diagnosis of West Nile Virus Encephalitis Virus Infection. Available online: https://www.paho.org/hq/dmdocuments/2016/WNV-Detection-and-Laboratory-Diagnosis-Dec-2016.pdf (accessed on 5 December 2022).
- Santos, P.D.; Michel, F.; Wylezich, C.; Höper, D.; Keller, M.; Holicki, C.M.; Szentiks, C.A.; Eiden, M.; Mulluneh, A.; Neubauer-Juric, A.; et al. Co-infections: Simultaneous detections of West Nile virus and Usutu virus in birds from Germany. Transbound. Emerg. Dis. 2021, 69, 776–792. [Google Scholar] [CrossRef]
- Hurtado-Monzón, A.M.; Cordero-Rivera, C.D.; Farfan-Morales, C.N.; Osuna-Ramos, J.F.; De Jesús-González, L.A.; Reyes-Ruiz, J.M.; Del Ángel, R.M. The role of anti-flavivirus humoral immune response in protection and pathogenesis. Rev. Med. Virol. 2020, 30, e2100. [Google Scholar] [CrossRef]
- Rey, F.A.; Stiasny, K.; Vaney, M.C.; Dellarole, M.; Heinz, F.X. The bright and the dark side of human antibody responses to flaviviruses: Lessons for vaccine design. EMBO Rep. 2018, 19, 206–224. [Google Scholar] [CrossRef]
- Salgado, R.; Hawks, S.A.; Frere, F.; Vázquez, A.; Huang, C.Y.H.; Duggal, N.K. West Nile virus vaccination protects against Usutu virus disease in mice. Viruses 2021, 13, 2352. [Google Scholar] [CrossRef]
- Phillpotts, R.J.; Stephenson, J.R.; Porterfield, J.S. Antiobody-dependend enhancement of tick-borne encephalitis virus infectivity. J. Gen. Virol. 1985, 66, 1831–1837. [Google Scholar] [CrossRef]
- Kubinski, M.; Beicht, J.; Gerlach, T.; Volz, A.; Sutter, G.; Rimmelzwaan, G.F. Tick-borne encephalitis virus: A quest for better vaccines against a virus on the rise. Vaccines 2020, 8, 451. [Google Scholar] [CrossRef]
- Carpio, K.L.; Barrett, A.D.T. Flavivirus NS1 and its potential in vaccine development. Vaccines 2021, 9, 622. [Google Scholar] [CrossRef]
- World Health Organization. Laboratory Testing for Zika Virus and Dengue Virus Infections. Available online: https://apps.who.int/iris/bitstream/handle/10665/359857/WHO-ZIKV-DENV-LAB-2022.1-eng.pdf (accessed on 5 December 2022).
- Frank, C.; Schmidt-Chanasit, J.; Ziegler, U.; Lachmann, R.; Preußel, K.; Offergeld, R. West Nile Virus in Germany: An Emerging Infection and Its Relevance for Transfusion Safety. Transfus. Med. Hemotherapy 2022, 49, 192–204. [Google Scholar] [CrossRef]
- Lourenço, J.; Barros, S.C.; Zé-Zé, L.; Damineli, D.S.C.; Giovanetti, M.; Osório, H.C.; Amaro, F.; Henriques, A.M.; Ramos, F.; Luís, T.; et al. West Nile virus transmission potential in Portugal. Commun. Biol. 2022, 5, 6. [Google Scholar] [CrossRef]
- Napp, S.; Petric´, D.; Busquets, N. West Nile virus and other mosquito-borne viruses present in Eastern Europe. Pathog. Glob. Health 2018, 112, 233–248. [Google Scholar] [CrossRef] [Green Version]
- Wahlberg, P.; Saikku, P.; Brummer-Korvenkontio, M. Tick-borne Viral Encephalitis in Finland. The Clinical Features of Kumlinge Disease during 1959–1987. J. Intern. Med. 1989, 225, 173–177. [Google Scholar] [CrossRef]
- Lipowski, D.; Popiel, M.; Perlejewski, K.; Nakamura, S.; Bukowska-O´sko, I.; Rzadkiewicz, E.; Dzieciatkowski, T.; Milecka, A.; Wenski, W.; Ciszek, M. A cluster of fatal Tick-borne encephalitis virus infection in organ transplant setting. J. Infect. Dis. 2017, 215, 896–901. [Google Scholar] [CrossRef] [Green Version]
- Gaibani, P.; Cavrini, F.; Gould, E.A.; Rossini, G.; Pierro, A.; Landini, M.P.; Sambri, V. Comparative genomic and phylogenetic analysis of the first Usutu virus isolate from a human patient presenting with neurological symptoms. PLoS ONE 2013, 8, e64761. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Jungbauer, C.; Aberle, S.W.; Kolodziejek, J.; Dimmel, K.; Stiasny, K.; Allerberger, F.; Nowotny, N. Usutu virus infections among blood donors, Austria, July and August 2017–Raising awereness for diagnostic challenges. Euro Surveill. 2017, 41, 17-00644. [Google Scholar]
- World Health Organization. Seventy-Fifth World Health Assembly: Human Organ and Tissue Transplantation. Available online: https://apps.who.int/gb/ebwha/pdf_files/WHA75/A75_41-en.pdf (accessed on 3 December 2022).
- Kolodziejek, J.; Seidel, B.; Jungbauer, C.; Dimmel, K.; Kolodziejek, M.; Rudolf, I.; Hubálek, Z.; Allerberger, F.; Nowotny, N. West Nile Virus positive blood donation and subsequent entomological investigation, Austria 2014. PLoS ONE 2015, 5, e0126381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, A.; Peeling, R.W.; Chu, M.C.; Gubler, D.J.; de Silva, A.M.; Harris, E.; Murtagh, M.; Chua, A.; Rodriquez, W.; Kelly, C.; et al. Innovative and new approaches to laboratory diagnosis of Zika and Dengue: A meeting report. J. Infect. Dis. 2017, 217, 1060–1068. [Google Scholar] [CrossRef]
- Cao, Z.; Sugimura, N.; Burgermeister, E.; Ebert, M.P.; Zuo, T.; Lan, P. The gut virome: A new microbiome component in health and disease. eBioMedicine 2022, 81, 104113. [Google Scholar] [CrossRef] [PubMed]
- Young, K.T.; Lahmers, K.K.; Sellers, H.S.; Stallknecht, D.E.; Poulson, R.L.; Saliki, S.T.; Tompkins, M.; Padykula, I.; Siepker, C.; Howerth, E.W.; et al. Randomly primed, strand-switching MinION-based sequencing for the detection and characterization of cultured RNA viruses. J. Vet. Diagn. Investig. 2021, 33, 202–215. [Google Scholar] [CrossRef]
- Brejová, B.; Boršová, K.; Hodorová, V.; Čabanová, V.; Reizigová, L.; Paul, E.D.; Čekan, P.; Klempa, B.; Nosek, J.; Vinář, T. A SARS-CoV-2 mutant from B.1.258 lineage with ∆H69/∆V70 deletion in the Spike protein circulating in Central Europe in the fall 2020. Virus Genes 2021, 57, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Kohler, S.; Zhao, S.; Mai, R.; Liu, Z.; Lu, H.; Xing, C. High-throughput, low-cost and rapid DNA sequencing using surface-coating techniques. BioRxiv 2020. [Google Scholar] [CrossRef]
- Amraoui, F.; Failloux, A.B. Chikungunya: An unexpected emergence in Europe. Curr. Opin. Virol. 2016, 21, 146–150. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Laboratory Test | Isolation of Virus from Clinical Specimen | Detection of Virus’ Nucleic Acid in Clinical Specimen | Specific Antibody Response (IgM) in CSF | IgM High Titre + Detection of IgG + Confirmation by Neutralisation | Seroconversion or Four-Fold Increase of Specific Antibodies in Paired Serum Samples |
|---|---|---|---|---|---|
| WNV | ✔ | ✔ | ✔ | ✔ | ✕ |
| TBEV | ✔ | ✔ | ✔ | ✔ * | ✔ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čabanová, V.; Kerlik, J.; Kirschner, P.; Rosochová, J.; Klempa, B.; Sláviková, M.; Ličková, M. Co-Circulation of West Nile, Usutu, and Tick-Borne Encephalitis Viruses in the Same Area: A Great Challenge for Diagnostic and Blood and Organ Safety. Viruses 2023, 15, 366. https://doi.org/10.3390/v15020366
Čabanová V, Kerlik J, Kirschner P, Rosochová J, Klempa B, Sláviková M, Ličková M. Co-Circulation of West Nile, Usutu, and Tick-Borne Encephalitis Viruses in the Same Area: A Great Challenge for Diagnostic and Blood and Organ Safety. Viruses. 2023; 15(2):366. https://doi.org/10.3390/v15020366
Chicago/Turabian StyleČabanová, Viktória, Jana Kerlik, Peter Kirschner, Jana Rosochová, Boris Klempa, Monika Sláviková, and Martina Ličková. 2023. "Co-Circulation of West Nile, Usutu, and Tick-Borne Encephalitis Viruses in the Same Area: A Great Challenge for Diagnostic and Blood and Organ Safety" Viruses 15, no. 2: 366. https://doi.org/10.3390/v15020366