
The Flavonoid Cyanidin Shows Immunomodulatory and Broad-Spectrum Antiviral Properties, Including SARS-CoV-2
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cells and Viruses
2.3. Antiviral Activity
2.4. Cytotoxicity Assay
2.5. Adsorption and Penetration Assay
2.6. RSV qRT-PCR Assay
2.7. Immunofluorescence Assay (IF) and Semiquantitative IF Analysis
2.8. Cytokine Determination
2.9. Pulmonary Infection in Mouse Model
2.10. Cytokines qRT-PCR
2.11. Transfections and Reporter Gene Assays
2.12. Statistical Analysis
3. Results
3.1. Broad-Spectrum Antiviral Activity of A18
3.2. Modulation of Cytokine Production and NF-kB Activation by A18 in RSV-Infected Epithelial and Macrophage Cell Lines
3.3. In Vivo Evaluation of A18 Antiviral Effect
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, A.M.; Baric, R.S.; Saphire, E.O.; Ulmer, J.B. Stopping pandemics before they start: Lessons learned from SARS-CoV-2. Science 2022, 375, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Salata, C.; Gribaudo, G. Tackling the Future Pandemics: Broad-Spectrum Antiviral Agents (BSAAs) Based on A-Type Proanthocyanidins. Molecules 2022, 27, 8353. [Google Scholar] [PubMed]
- Mason, S.; Devincenzo, J.P.; Toovey, S.; Wu, J.Z.; Whitley, R.J. Comparison of antiviral resistance across acute and chronic viral infections. Antivir. Res. 2018, 158, 103–112. [Google Scholar] [PubMed]
- Maarifi, G.; Martin, M.F.; Zebboudj, A.; Boulay, A.; Nouaux, P.; Fernandez, J.; Lagisquet, J.; Garcin, D.; Gaudin, R.; Arhel, N.J.; et al. Identifying enhancers of innate immune signaling as broad-spectrum antivirals active against emerging viruses. Cell Chem. Biol. 2022, 29, 1113–1125.e6. [Google Scholar] [CrossRef]
- Pasquero, S.; Gugliesi, F.; Griffante, G.; Dell’Oste, V.; Biolatti, M.; Albano, C.; Bajetto, G.; Delbue, S.; Signorini, L.; Dolci, M.; et al. Novel antiviral activity of PAD inhibitors against human beta-coronaviruses HCoV-OC43 and SARS-CoV-2. Antivir. Res. 2022, 200, 105278. [Google Scholar] [CrossRef]
- Milani, D.; Caruso, L.; Zauli, E.; Al Owaifeer, A.M.; Secchiero, P.; Zauli, G.; Gemmati, D.; Tisato, V. p53/NF-kB Balance in SARS-CoV-2 Infection: From OMICs, Genomics and Pharmacogenomics Insights to Tailored Therapeutic Perspectives (COVIDomics). Front. Pharmacol. 2022, 13, 871583. [Google Scholar]
- Cai, M.; Liao, Z.; Zou, X.; Xu, Z.; Wang, Y.; Li, T.; Li, Y.; Ou, X.; Deng, Y.; Guo, Y.; et al. Herpes Simplex Virus 1 UL2 Inhibits the TNF-α-Mediated NF-κB Activity by Interacting With p65/p50. Front. Immunol. 2020, 11, 549. [Google Scholar] [CrossRef]
- Sallenave, J.M.; Guillot, L. Innate Immune Signaling and Proteolytic Pathways in the Resolution or Exacerbation of SARS-CoV-2 in COVID-19: Key Therapeutic Targets? Front. Immunol. 2020, 11, 1229. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Zhao, Y.; Ivanciuc, T.; Sun, H.; Wakamiya, M.; Garofalo, R.P.; Brasier, A.R. Central role of the NF-κB pathway in the scgb1a1-expressing epithelium in mediating respiratory syncytial virus-induced airway inflammation. J. Virol. 2018, 92, e00441-18. [Google Scholar] [CrossRef]
- Hao, X.; Guan, R.; Huang, H.; Yang, K.; Wang, L.; Wu, Y. Anti-inflammatory activity of cyanidin-3-O-glucoside and cyanidin-3-O-glucoside liposomes in THP-1 macrophages. Food Sci. Nutr. 2021, 9, 6480–6491. [Google Scholar] [CrossRef]
- Yang, M.; Lu, X.; Xu, J.; Liu, X.; Zhang, W.; Guan, R.; Zhong, H. Cellular uptake, transport mechanism and anti-inflammatory effect of cyanidin-3-glucoside nanoliposomes in Caco-2/RAW 264.7 co-culture model. Front. Nutr. 2022, 9, 995391. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, L.; Li, Z.; Liu, Q. Cyanidin-3-O-glucoside, cyanidin, and oxidation products of cyanidin protect neuronal function through alleviating inflammation and oxidative damage. J. Food Sci. 2022, 87, 2159–2172. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Ye, Z.; Zhang, W.; Dai, B.; Chen, G.; Wang, L.; Shao, X.; Xiang, A.; Lu, Z.; Shi, J. Cyanidin Chloride Improves LPS-Induced Depression-Like Behavior in Mice by Ameliorating Hippocampal Inflammation and Excitotoxicity. ACS Chem. Neurosci. 2022, 13, 3023–3033. [Google Scholar] [CrossRef]
- Liu, C.; Zhu, L.; Fukuda, K.; Ouyang, S.; Chen, X.; Wang, C.; Zhang, C.J.; Martin, B.; Gu, C.; Qin, L.; et al. The flavonoid cyanidin blocks binding of the cytokine interleukin-17A to the IL-17RA subunit to alleviate inflammation in vivo. Sci. Signal. 2017, 21, 10. [Google Scholar]
- Wang, B.; Cui, S.; Mao, B.; Zhang, Q.; Tian, F.; Zhao, J.; Tang, X.; Chen, W. Cyanidin Alleviated CCl4-Induced Acute Liver Injury by Regulating the Nrf2 and NF-κB Signaling Pathways. Antioxidants 2022, 11, 2383. [Google Scholar] [CrossRef]
- Thummayot, S.; Tocharus, C.; Jumnongprakhon, P.; Suksamrarn, A.; Tocharus, J. Cyanidin attenuates Aβ 25-35-induced neuroinflammation by suppressing NF-κB activity downstream of TLR4/NOX4 in human neuroblastoma cells. Acta Pharmacol. Sin. 2018, 39, 1439–1452. [Google Scholar]
- Michelini, F.M.; Alché, L.E.; Bueno, C.A. Virucidal, antiviral and immunomodulatory activities of β-escin and Aesculus hippocastanum extract. J. Pharm. Pharmacol. 2018, 70, 1561–1571. [Google Scholar]
- Salinas, F.M.; Vázquez, L.; Gentilini, M.V.; O Donohoe, A.; Regueira, E.; Nabaes Jodar, M.S.; Viegas, M.; Michelini, F.M.; Hermida, G.; Alché, L.E.; et al. Aesculus hippocastanum L. seed extract shows virucidal and antiviral activities against respiratory syncytial virus (RSV) and reduces lung inflammation in vivo. Antiviral. Res. 2019, 164, 1–11. [Google Scholar] [CrossRef]
- Zappia, C.D.; Soto, A.; Granja-Galeano, G.; Fenoy, I.; Fernandez, N.; Davio, C.A.; Shayo, C.; Fitzsimons, C.P.; Goldman, A.; Monczor, F. Azelastine potentiates antiasthmatic dexamethasone effect on a murine asthma model. Pharmacol. Res. Perspect. 2019, 29, e00531. [Google Scholar] [CrossRef]
- Carvajal, J.; Avellaneda, A.; Salazar-Ardiles, C.; Maya, J.; Kalergis, A.; Lay, M. Host Components Contributing to Respiratory Syncytial Virus Pathogenesis. Front. Immunol. 2019, 10, 2152. [Google Scholar]
- Salinas, F.M.; Nebreda, A.D.; Vázquez, L.; Gentilini, M.V.; Marini, V.; Benedetti, M.; Nabaes Jodar, M.S.; Viegas, M.; Shayo, C.; Bueno, C.A. Imiquimod suppresses respiratory syncytial virus (RSV) replication via PKA pathway and reduces RSV induced-inflammation and viral load in mice lungs. Antiviral. Res. 2020, 179, 104817. [Google Scholar] [PubMed]
- Lukacs, N.W.; Moore, M.L.; Rudd, B.D.; Berlin, A.A.; Collins, R.D.; Olson, S.J.; Ho, S.B.; Peebles Jr, R.S. Differential immune responses and pulmonary pathophysiology are induced by two different strains of respiratory syncytial virus. Am. J. Pathol. 2006, 169, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Rudd, P.A.; Chen, W.; Mahalingam, S. Mouse and cotton rat models of human respiratory syncytial virus. Methods Mol. Biol. 2016, 1442, 209–217. [Google Scholar]
- Stokes, K.L.; Chi, M.H.; Sakamoto, K.; Newcomb, D.C.; Currier, M.G.; Huckabee, M.M.; Lee, S.; Goleniewska, K.; Pretto, C.; Williams, J.V.; et al. Differential pathogenesis of respiratory syncytial virus clinical isolates in BALB/c mice. J. Virol. 2011, 85, 5782–5793. [Google Scholar]
- Woolums, A.; Lee, S.; Moore, M. Animal models of respiratory syncytial virus pathogenesis and vaccine development: Opportunities and future directions. In Human Respiratory Syncytial Virus Infection; Resch, B., Ed.; InTech: London, UK, 2011; ISBN 978-953-307-718-5. [Google Scholar]
- Cohen, M.S. Early treatment to prevent progression of SARS-CoV-2 infection. Lancet Respir. Med. 2022, 10, 930–931. [Google Scholar] [CrossRef]
- D’Abramo, A.; Vita, S.; Nicastri, E. The unmet need for COVID-19 treatment in immunocompromised patients. BMC Infect. Dis. 2022, 22, 930. [Google Scholar]
- Therapeutics and COVID-19: Living Guideline (v13.1). World Health Organization (WHO). Available online: https://app.magicapp.org/#/guideline/6914 (accessed on 15 March 2023).
- Li, Y.; Wang, X.; Blau, D.M.; Caballero, M.T.; Feikin, D.R.; Gill, C.J.; Madhi, S.A.; Omer, S.B.; Simões, E.A.F.; Campbell, H.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in children younger than 5 years in 2019: A systematic análisis. Lancet 2022, 399, 2047–2064. [Google Scholar] [CrossRef]
- Nguyen-Van-Tam, J.S.; O’Leary, M.; Martin, E.T.; Heijnen, E.; Callendret, B.; Fleischhackl, R.; Comeaux, C.; Tran, T.M.P.; Weber, K. Burden of respiratory syncytial virus infection in older and high-risk adults: A systematic review and meta-analysis of the evidence from developed countries. Eur. Respir. Rev. 2022, 31, 220105. [Google Scholar] [CrossRef]
- Gatt, D.; Martin, I.; AlFouzan, R.; Moraes, T. Prevention and Treatment Strategies for Respiratory Syncytial Virus (RSV). Pathogens 2023, 12, 154. [Google Scholar] [CrossRef]
- Greenan, E.; Gallagher, S.; Khalil, R.; Murphy, C.C.; Ní Gabhann-Dromgoole, J. Advancing Our Understanding of Corneal Herpes Simplex Virus-1 Immune Evasion Mechanisms and Future Therapeutics. Viruses 2021, 13, 1856. [Google Scholar] [CrossRef]
- Kim, H.; Chung, M.S. Antiviral Activities of Mulberry (Morus alba) Juice and Seed against Influenza Viruses. Evid Based Complement. Altern. Med. 2018, 2018, 2606583. [Google Scholar] [CrossRef] [PubMed]
- Sivarajan, R.; Oberwinkler, H.; Roll, V.; König, E.M.; Steinke, M.; Bodem, J. A defined anthocyanin mixture sourced from bilberry and black currant inhibits Measles virus and various herpesviruses. BMC Complement. Med. Ther. 2022, 22, 181. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Calvo, A.; Jiménez de Oya, N.; Martín-Acebes, M.A.; Garcia-Moruno, E.; Saiz, J.C. Antiviral Properties of the Natural Polyphenols Delphinidin and Epigallocatechin Gallate against the Flaviviruses West Nile Virus, Zika Virus, and Dengue Virus. Front. Microbiol. 2017, 8, 1314. [Google Scholar] [PubMed]
- Bajrai, L.H.; El-Kafrawy, S.A.; Hassan, A.M.; Tolah, A.M.; Alnahas, R.S.; Sohrab, S.S.; Rehan, M.; Azhar, E.I. In vitro screening of anti-viral and virucidal effects against SARS-CoV-2 by Hypericum perforatum and Echinacea. Sci. Rep. 2022, 12, 21723. [Google Scholar] [CrossRef]
- Shrestha, A.; Marahatha, R.; Basnet, S.; Regmi, B.P.; Katuwal, S.; Dahal, S.R.; Sharma, K.R.; Adhikari, A.; Chandra Basnyat, R.; Parajuli, N. Molecular Docking and Dynamics Simulation of Several Flavonoids Predict Cyanidin as an Effective Drug Candidate against SARS-CoV-2 Spike Protein. Adv. Pharmacol. Pharm. Sci. 2022, 2022, 3742318. [Google Scholar] [PubMed]
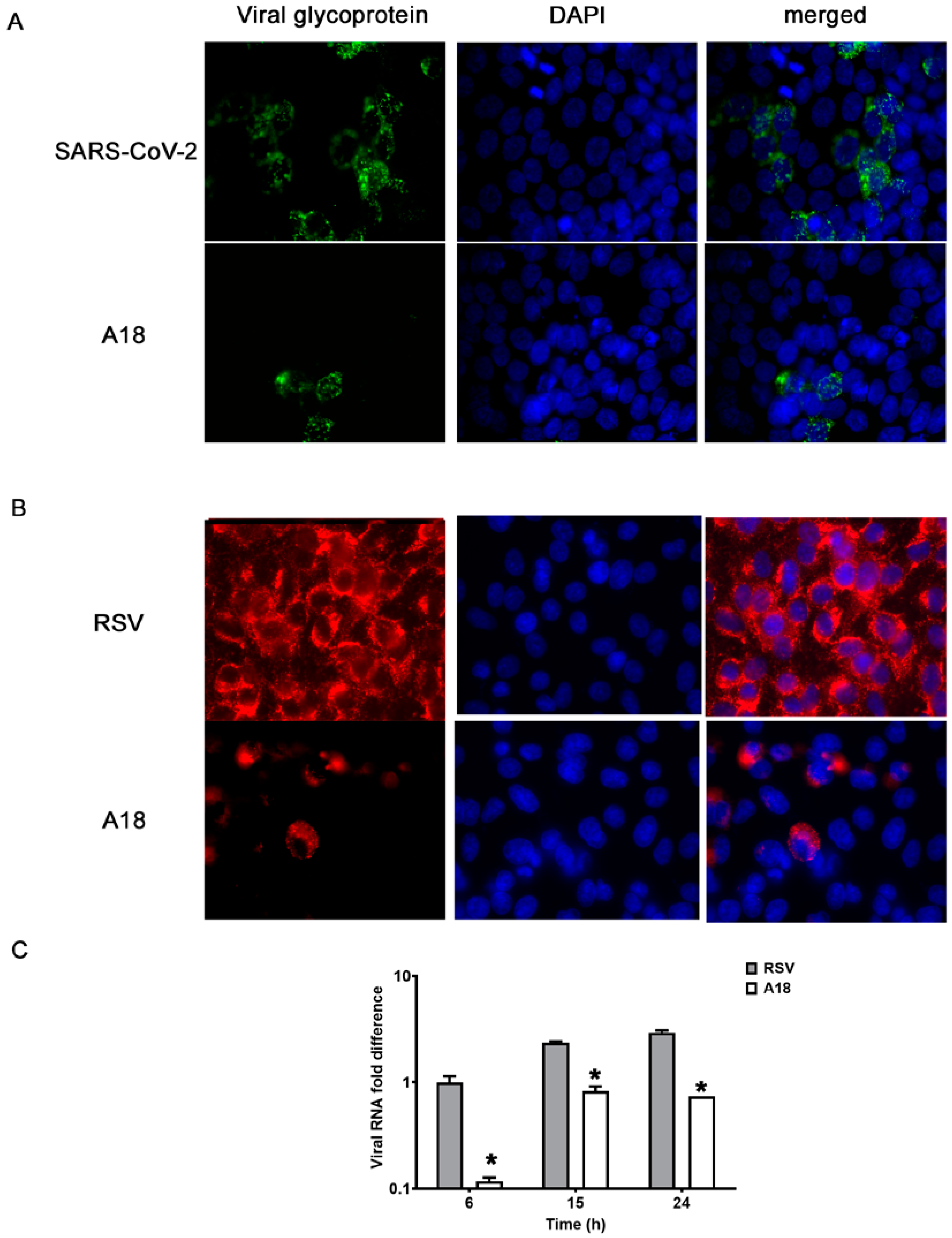


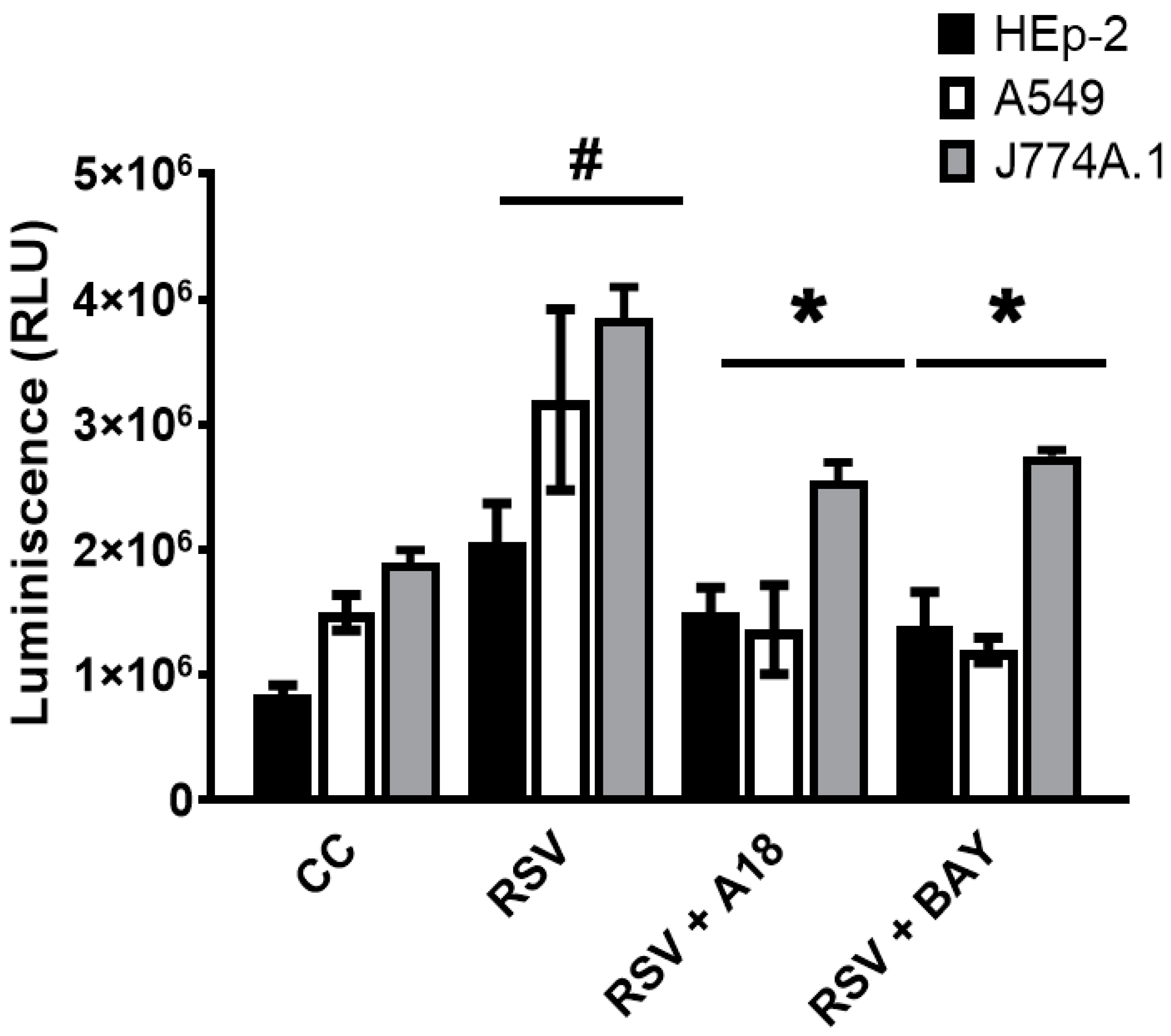
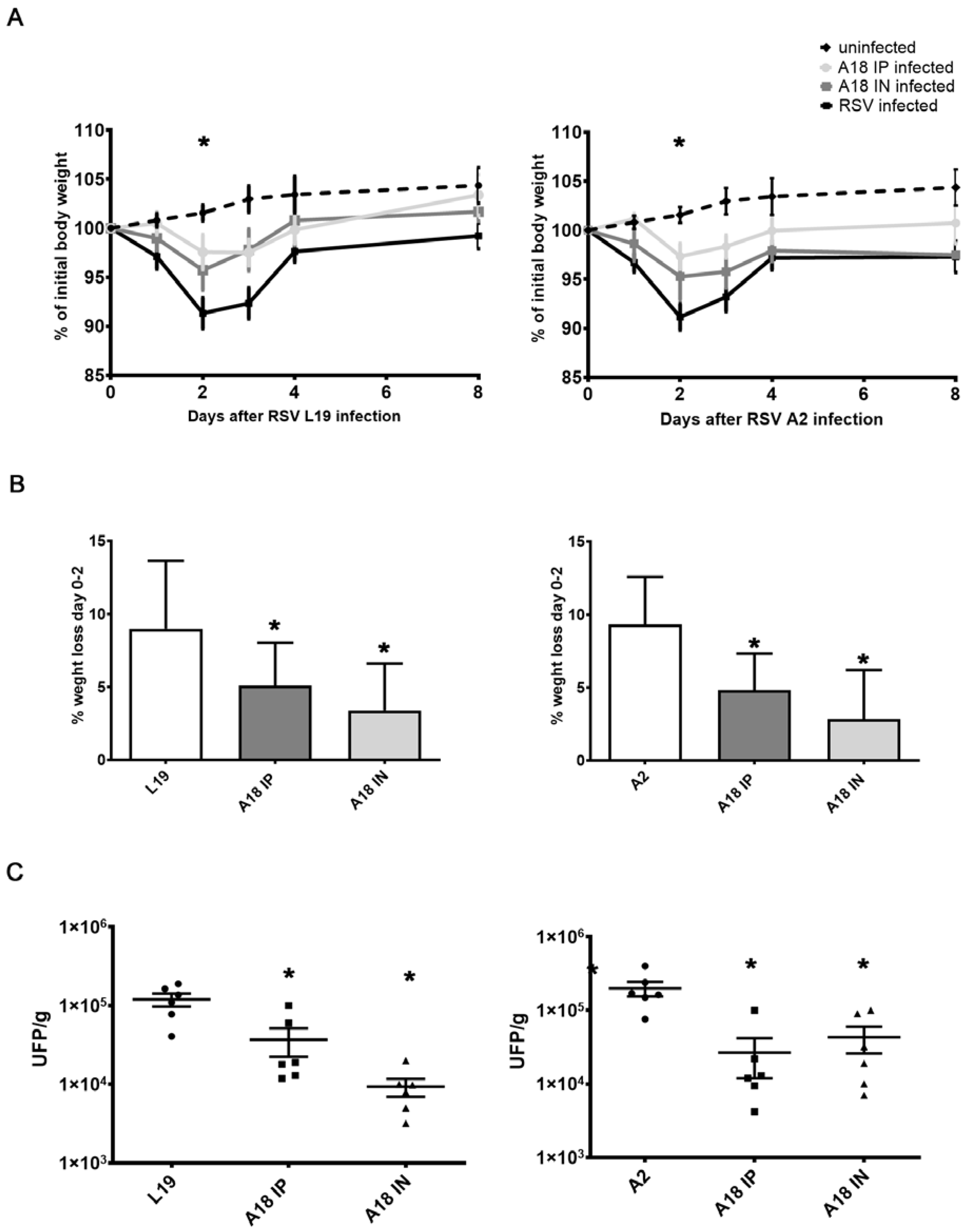



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RSV Line 19 | RSV A2 | HSV-1 KOS | HSV-1 Field | HSV-1 B2006 | SARS-CoV-2 | Canine Coronavirus | |||
|---|---|---|---|---|---|---|---|---|---|
| Cell Line | HEp-2 | A549 | HEp-2 | A549 | HCLE | Calu-3 | CRFK | ||
| EC50 | 8 ± 0.1 | 9.8 ± 0.08 | 3 ± 0.06 | 8.5 ± 0.1 | 5.5± 0.1 | 7.6± 0.05 | 4.6± 0.05 | 8.9± 0.09 | 7.7± 0.1 |
| SI | 25 | 20 | 67 | 24 | 46 | 26 | 43 | 23 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente, J.; Benedetti, M.; Martelliti, P.; Vázquez, L.; Gentilini, M.V.; Peñaranda Figueredo, F.A.; Nabaes Jodar, M.S.; Viegas, M.; Barquero, A.A.; Bueno, C.A. The Flavonoid Cyanidin Shows Immunomodulatory and Broad-Spectrum Antiviral Properties, Including SARS-CoV-2. Viruses 2023, 15, 989. https://doi.org/10.3390/v15040989
Vicente J, Benedetti M, Martelliti P, Vázquez L, Gentilini MV, Peñaranda Figueredo FA, Nabaes Jodar MS, Viegas M, Barquero AA, Bueno CA. The Flavonoid Cyanidin Shows Immunomodulatory and Broad-Spectrum Antiviral Properties, Including SARS-CoV-2. Viruses. 2023; 15(4):989. https://doi.org/10.3390/v15040989
Chicago/Turabian StyleVicente, Josefina, Martina Benedetti, Paula Martelliti, Luciana Vázquez, María Virginia Gentilini, Freddy Armando Peñaranda Figueredo, Mercedes Soledad Nabaes Jodar, Mariana Viegas, Andrea Alejandra Barquero, and Carlos Alberto Bueno. 2023. "The Flavonoid Cyanidin Shows Immunomodulatory and Broad-Spectrum Antiviral Properties, Including SARS-CoV-2" Viruses 15, no. 4: 989. https://doi.org/10.3390/v15040989