
Identification of New Microfoci and Genetic Characterization of Tick-Borne Encephalitis Virus Isolates from Eastern Germany and Western Poland

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Collection
2.2. Molecular Detection
2.3. Statistical and Phylogenetic Analyses
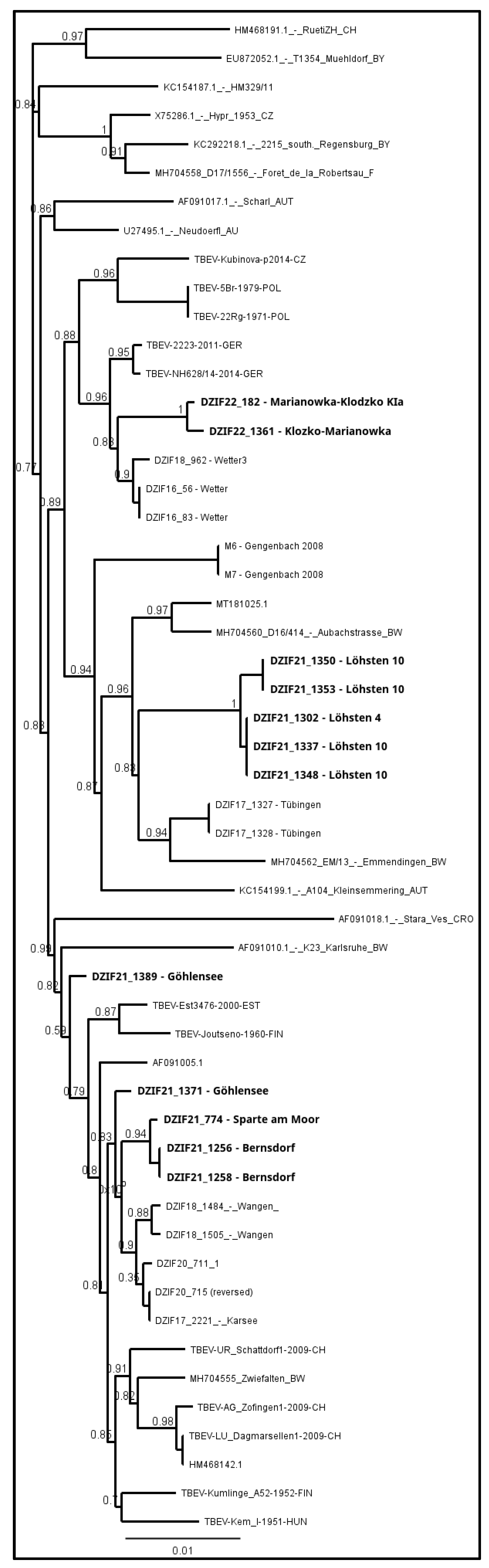
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Worku, D.A. Tick-borne encephalitis (TBE): From tick to pathology. J. Clin. Med. 2023, 12, 6859. [Google Scholar] [CrossRef] [PubMed]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Dobler, G.; Kaier, K.; Hehn, P.; Böhmer, M.M.; Kreusch, T.M.; Borde, J.P. Tick-borne encephalitis virus vaccination breakthrough infections in Germany: A retrospective analysis from 2001 to 2018. Clin. Microbiol. Infect. 2020, 26, 1090.e7–1090.e13. [Google Scholar] [CrossRef]
- Borde, J.P.; Glaser, R.; Braun, K.; Riach, N.; Hologa, R.; Kaier, K.; Chitimia-Dobler, L.; Dobler, G. Decoding the Geography of Natural TBEV Microfoci in Germany: A Geostatistical Approach Based on Land-Use Patterns and Climatological Conditions. Int. J. Environ. Res. Public Health 2022, 19, 11830. [Google Scholar] [CrossRef] [PubMed]
- Zeman, P. Cyclic patterns in the central European tick-borne encephalitis incidence series. Epidemiol. Infect. 2017, 145, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Zeman, P.; Beneš, C. A tick-borne encephalitis ceiling in Central Europe has moved upwards during the last 30 years: Possible impact of global warming? Int. J. Med. Microbiol. 2004, 293, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Michelitsch, A.; Wernike, K.; Klaus, C.; Dobler, G.; Beer, M. Exploring the Reservoir Hosts of Tick-Borne Encephalitis Virus. Viruses 2019, 11, 669. [Google Scholar] [CrossRef] [PubMed]
- Paradowska-Stankiewicz, I.; Zbrzeźniak, J. (Eds.) Kleszczowe Zapalenie Mózgu w Polsce i na Świecie: Ocena Sytuacji Epidemiologicznej KZM w Polsce w Latach 2015–2019 w Oparciu o Dane Pochodzące z Nadzoru Epidemiologicznego; Narodowy Instytut Zdrowia Publicznego—Państwowy Zakład Higieny: Warsaw, Poland, 2021; ISBN 978-83-65870-39-1. (In Polish) [Google Scholar]
- Holzmann, H.; Aberle, S.W.; Stiasny, K.; Werner, P.; Mischak, A.; Zainer, B.; Netzer, M.; Koppi, S.; Bechter, E.; Heinz, F.X. Tick-borne encephalitis from eating goat cheese in a mountain region of Austria. Emerg. Infect. Dis. 2009, 15, 1671–1673. [Google Scholar] [CrossRef]
- Brockmann, S.O.; Oehme, R.; Buckenmaier, T.; Beer, M.; Jeffery-Smith, A.; Spannenkrebs, M.; Haag-Milz, S.; Wagner-Wiening, C.; Schlegel, C.; Fritz, J.; et al. A cluster of two human cases of tick-borne encephalitis (TBE) transmitted by unpasteurised goat milk and cheese in Germany, May 2016. Eurosurveillance 2018, 23, 8–14. [Google Scholar] [CrossRef]
- Paulsen, K.M.; Stuen, S.; das Neves, C.G.; Suhel, F.; Gurung, D.; Soleng, A.; Stiasny, K.; Vikse, R.; Andreassen, Å.K.; Granquist, E.G. Tick-borne encephalitis virus in cows and unpasteurized cow milk from Norway. Zoonoses Public Health 2019, 66, 216–222. [Google Scholar] [CrossRef]
- Caini, S.; Szomor, K.; Ferenczi, E.; Székelyné Gáspár, Á.; Csohán, Á.; Krisztalovics, K.; Molnár, Z.; Horváth, J.K. Tick-borne encephalitis transmitted by unpasteurised cow milk in western Hungary, September to October 2011. Eurosurveillance 2012, 17, 20128. [Google Scholar] [CrossRef]
- Geeraedts, F.; Wertenbroek, A.; de Klerk, J.; Prick, J.J.; Reichman, L.J.A.; Hess, D.; Bosma, F.; Reimerink, J.; Skidmore, B.; Laverman, G.D. Defining a risk area for tick-borne encephalitis (TBE) in a country where TBE is emerging, the Netherlands, July 2016-October 2020. Ticks Tick Borne Dis. 2022, 13, 101898. [Google Scholar] [CrossRef]
- Hellenbrand, W.; Kreusch, T.; Böhmer, M.M.; Wagner-Wiening, C.; Dobler, G.; Wichmann, O.; Altmann, D. Epidemiology of Tick-Borne Encephalitis (TBE) in Germany, 2001–2018. Pathogens 2019, 8, 42. [Google Scholar] [CrossRef]
- Topp, A.-K.; Springer, A.; Dobler, G.; Bestehorn-Willmann, M.; Monazahian, M.; Strube, C. New and Confirmed Foci of Tick-Borne Encephalitis Virus (TBEV) in Northern Germany Determined by TBEV Detection in Ticks. Pathogens 2022, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Sulik, M.; Toczyłowski, K.; Grygorczuk, S. Epidemiology of tick-borne encephalitis in Poland (2010–2019) and the impact of the COVID-19 pandemic on the notified incidence of the disease. Przegl Epidemiol. 2021, 75, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Zanotto, P.M.; Gao, G.F.; Gritsun, T.; Marin, M.S.; Jiang, W.R.; Venugopal, K.; Reid, H.W.; Gould, E.A. An arbovirus cline across the northern hemisphere. Virology 1995, 210, 152–159. [Google Scholar] [CrossRef]
- Heinze, D.M.; Gould, E.A.; Forrester, N.L. Revisiting the clinal concept of evolution and dispersal for the tick-borne flaviviruses by using phylogenetic and biogeographic analyses. J. Virol. 2012, 86, 8663–8671. [Google Scholar] [CrossRef]
- Klaus, C.; Hoffmann, B.; Hering, U.; Mielke, B.; Sachse, K.; Beer, M.; Süss, J. Tick-borne encephalitis (TBE) virus prevalence and virus genome characterization in field-collected ticks (Ixodes ricinus) from risk, non-risk and former risk areas of TBE, and in ticks removed from humans in Germany. Clin. Microbiol. Infect. 2010, 16, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Chitimia-Dobler, L.; Lemhöfer, G.; Król, N.; Bestehorn, M.; Dobler, G.; Pfeffer, M. Repeated isolation of tick-borne encephalitis virus from adult Dermacentor reticulatus ticks in an endemic area in Germany. Parasit. Vectors 2019, 12, 90. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany; Cham, Switzerland, 2017; ISBN 9783319637600. [Google Scholar]
- Siuda, K. Ticks of Poland II (Acari: Ixodida): Systematics and Distribution; Polskie Towarzystwo Parazytologiczne: Warsaw, Poland, 1993; ISBN 839013490X. (In Polish) [Google Scholar]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar]
- Pettersson, J.H.-O.; Golovljova, I.; Vene, S.; Jaenson, T.G.T. Prevalence of tick-borne encephalitis virus in Ixodes ricinus ticks in northern Europe with particular reference to Southern Sweden. Parasit. Vectors 2014, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Kjær, L.J.; Klitgaard, K.; Soleng, A.; Edgar, K.S.; Lindstedt, H.E.H.; Paulsen, K.M.; Andreassen, Å.K.; Korslund, L.; Kjelland, V.; Slettan, A.; et al. Spatial patterns of pathogen prevalence in questing Ixodes ricinus nymphs in southern Scandinavia, 2016. Sci. Rep. 2020, 10, 19376. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Chitimia-Dobler, L.; Dautel, H.; Meyer-Kayser, E.; Kahl, O. Atlas of ticks (Acari: Argasidae, Ixodidae) in Germany. Exp. Appl. Acarol. 2021, 84, 183–214. [Google Scholar] [CrossRef] [PubMed]
- Kiewra, D.; Czułowska, A.; Dyczko, D.; Zieliński, R.; Plewa-Tutaj, K. First record of Haemaphysalis concinna (Acari: Ixodidae) in Lower Silesia, SW Poland. Exp. Appl. Acarol. 2019, 77, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Dwużnik-Szarek, D.; Mierzejewska, E.J.; Rodo, A.; Goździk, K.; Behnke-Borowczyk, J.; Kiewra, D.; Kartawik, N.; Bajer, A. Monitoring the expansion of Dermacentor reticulatus and occurrence of canine babesiosis in Poland in 2016–2018. Parasit Vectors 2021, 14, 267. [Google Scholar] [CrossRef]
- Drehmann, M.; Chitimia-Dobler, L.; Lindau, A.; Frank, A.; Mai, S.; Fachet, K.; Hauck, D.; Knoll, S.; Strube, C.; Lühken, R.; et al. Ixodes frontalis: A neglected but ubiquitous tick species in Germany. Exp. Appl. Acarol. 2019, 78, 79–91. [Google Scholar] [CrossRef]
- Ličková, M.; Fumačová Havlíková, S.; Sláviková, M.; Slovák, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne encephalitis virus. Ticks Tick Borne Dis. 2020, 11, 101414. [Google Scholar] [CrossRef] [PubMed]
- Süss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia-an overview. Ticks Tick Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef]
- Biernat, B.; Karbowiak, G.; Werszko, J.; Stańczak, J. Prevalence of tick-borne encephalitis virus (TBEV) RNA in Dermacentor reticulatus ticks from natural and urban environment, Poland. Exp. Appl. Acarol. 2014, 64, 543–551. [Google Scholar] [CrossRef]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.M.; Pedersen, B.N.; Soleng, A.; Okbaldet, Y.B.; Pettersson, J.H.-O.; Dudman, S.G.; Ottesen, P.; Vik, I.S.S.; Vainio, K.; Andreassen, Å. Prevalence of tick-borne encephalitis virus in Ixodes ricinus ticks from three islands in north-western Norway. Apmis 2015, 123, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Wójcik-Fatla, A.; Cisak, E.; Zając, V.; Zwoliński, J.; Dutkiewicz, J. Prevalence of tick-borne encephalitis virus in Ixodes ricinus and Dermacentor reticulatus ticks collected from the Lublin region (eastern Poland). Ticks Tick. Borne Dis. 2011, 2, 16–19. [Google Scholar] [CrossRef]
- Borde, J.P.; Kaier, K.; Hehn, P.; Matzarakis, A.; Frey, S.; Bestehorn, M.; Dobler, G.; Chitimia-Dobler, L. The complex interplay of climate, TBEV vector dynamics and TBEV infection rates in ticks-Monitoring a natural TBEV focus in Germany, 2009–2018. PLoS ONE 2021, 16, e0244668. [Google Scholar] [CrossRef]
- Ott, D.; Ulrich, K.; Ginsbach, P.; Öhme, R.; Bock-Hensley, O.; Falk, U.; Teinert, M.; Lenhard, T. Tick-borne encephalitis virus (TBEV) prevalence in field-collected ticks (Ixodes ricinus) and phylogenetic, structural and virulence analysis in a TBE high-risk endemic area in southwestern Germany. Parasit. Vectors 2020, 13, 303. [Google Scholar] [CrossRef]
- Biernat, B.; Cieniuch, S.; Stańczak, J. Detection of TBEV RNA in Ixodes ricinus ticks in north-eastern Poland. Ann. Agric. Environ. Med. 2014, 21, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Chitimia-Dobler, L.; Bestehorn-Willmann, M.; Lindau, A.; Drehmann, M.; Stroppel, G.; Hengge, H.; Mackenstedt, U.; Kaier, K.; Dobler, G.; et al. The Emergence and Dynamics of Tick-Borne Encephalitis Virus in a New Endemic Region in Southern Germany. Microorganisms 2022, 10, 2125. [Google Scholar] [CrossRef] [PubMed]
- Borde, J.P.; Zajkowska, J. Chapter 5: TBE in adults. In The TBE Book, 6th ed.; Dobler, G., Erber, W., Bröker, M., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2023; pp. 79–93. [Google Scholar]
- Jaenson, T.G.T.; Petersson, E.H.; Jaenson, D.G.E.; Kindberg, J.; Pettersson, J.H.-O.; Hjertqvist, M.; Medlock, J.M.; Bengtsson, H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: Incidence of human TBE correlates with abundance of deer and hares. Parasit. Vectors 2018, 11, 477. [Google Scholar] [CrossRef]
- Kriz, B.; Daniel, M.; Benes, C.; Maly, M. The role of game (wild boar and roe deer) in the spread of tick-borne encephalitis in the Czech Republic. Vector Borne Zoonotic Dis. 2014, 14, 801–807. [Google Scholar] [CrossRef]
- Balling, A.; Plessow, U.; Beer, M.; Pfeffer, M. Prevalence of antibodies against tick-borne encephalitis virus in wild game from Saxony, Germany. Ticks Tick. Borne Dis. 2014, 5, 805–809. [Google Scholar] [CrossRef]
- Burke, G.; Wikel, S.K.; Spielman, A.; Telford, S.R.; McKay, K.; Krause, P.J. Hypersensitivity to ticks and Lyme disease risk. Emerg. Infect. Dis. 2005, 11, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Hofhuis, A.; Herremans, T.; Notermans, D.W.; Sprong, H.; Fonville, M.; van der Giessen, J.W.B.; van Pelt, W. A prospective study among patients presenting at the general practitioner with a tick bite or erythema migrans in The Netherlands. PLoS ONE 2013, 8, e64361. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | State/Voivodship | Name of Site | GPS Coordinates | Short Description |
|---|---|---|---|---|
| Germany | Brandenburg | Cottbus | 51.72912, 14.34782 | the forest between a settlement and the lake |
| Frauendorf | 51.69781, 14.38748 | woodlands near the residential area | ||
| Geierwalder See | 51.49863, 14.13382 | green areas between lakes | ||
| Deulowitzer See | 51.92312, 14.64813 | the forest around the lake | ||
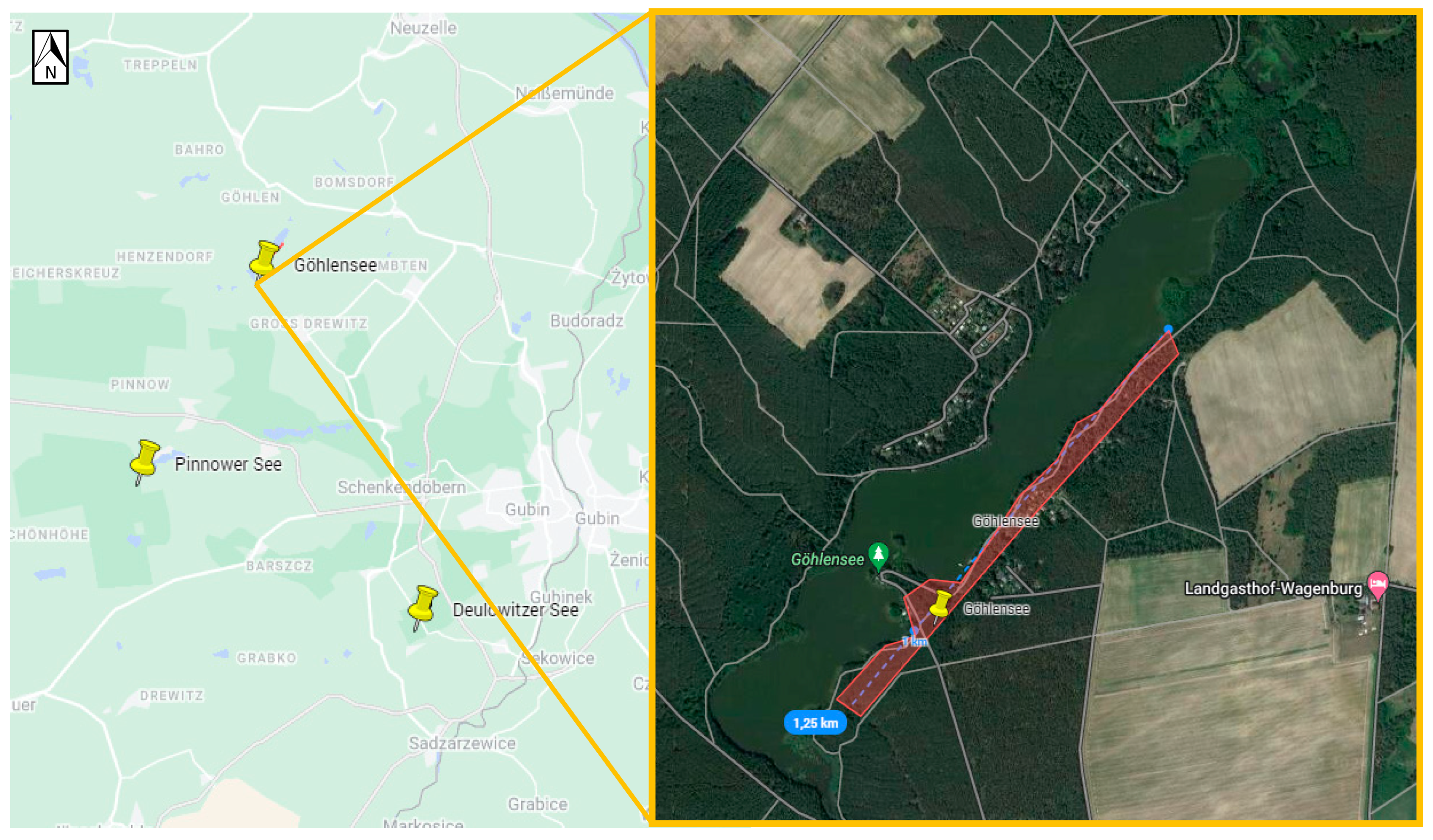
| Göhlensee | 52.02116, 14.57116 | the walking path on the eastern side of the lake | ||
| Heidesee | 52.25275, 13.77984 | the forested area between lakes | ||
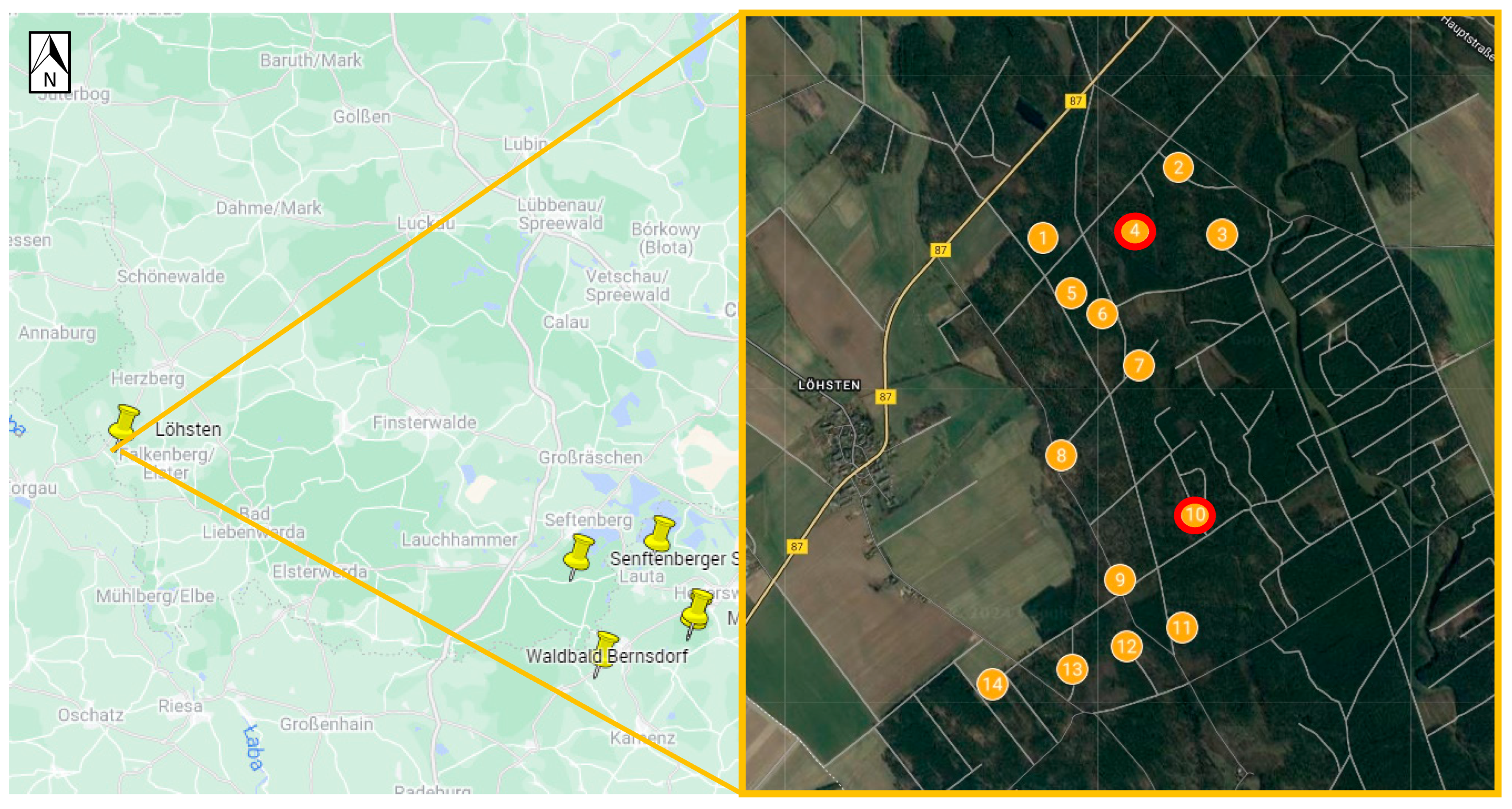
| Löhsten | 51.62287, 13.16589 | 14 sub-spots in the forest | ||
| Pinnower See | 51.96456, 14.51513 | woodlands between the lake and the settlement | ||
| Spremberg See | 51.64993, 14.41396 | green areas around the lake | ||
| Mecklenburg-West Pomerania | Hiddensee Island | 54.53988, 13.09666 | the walking area on the island | |
| Saxony | Michalken | 51.41198, 14.20098 | the forest near residential areas | |
| Senftenberger See | 51.47759, 13.98911 | the walking path along the southern part of the lake | ||
| Sparte am Moor | 51.41669, 14.20562 | the walking path in a forest | ||
| Waldbad Bernsdorf | 51.36904, 14.03123 | walking paths in the forest around the small lake | ||
| Poland | Lubush | Jezioro Głębokie | 51.74357, 14.8375 | the forest around the lake |
| Jeziory Wysokie | 51.78434, 14.76023 | walking paths in the forest | ||
| Lower Silesia | Kamienna | 50.28478, 16.75311 | green areas around the settlement | |
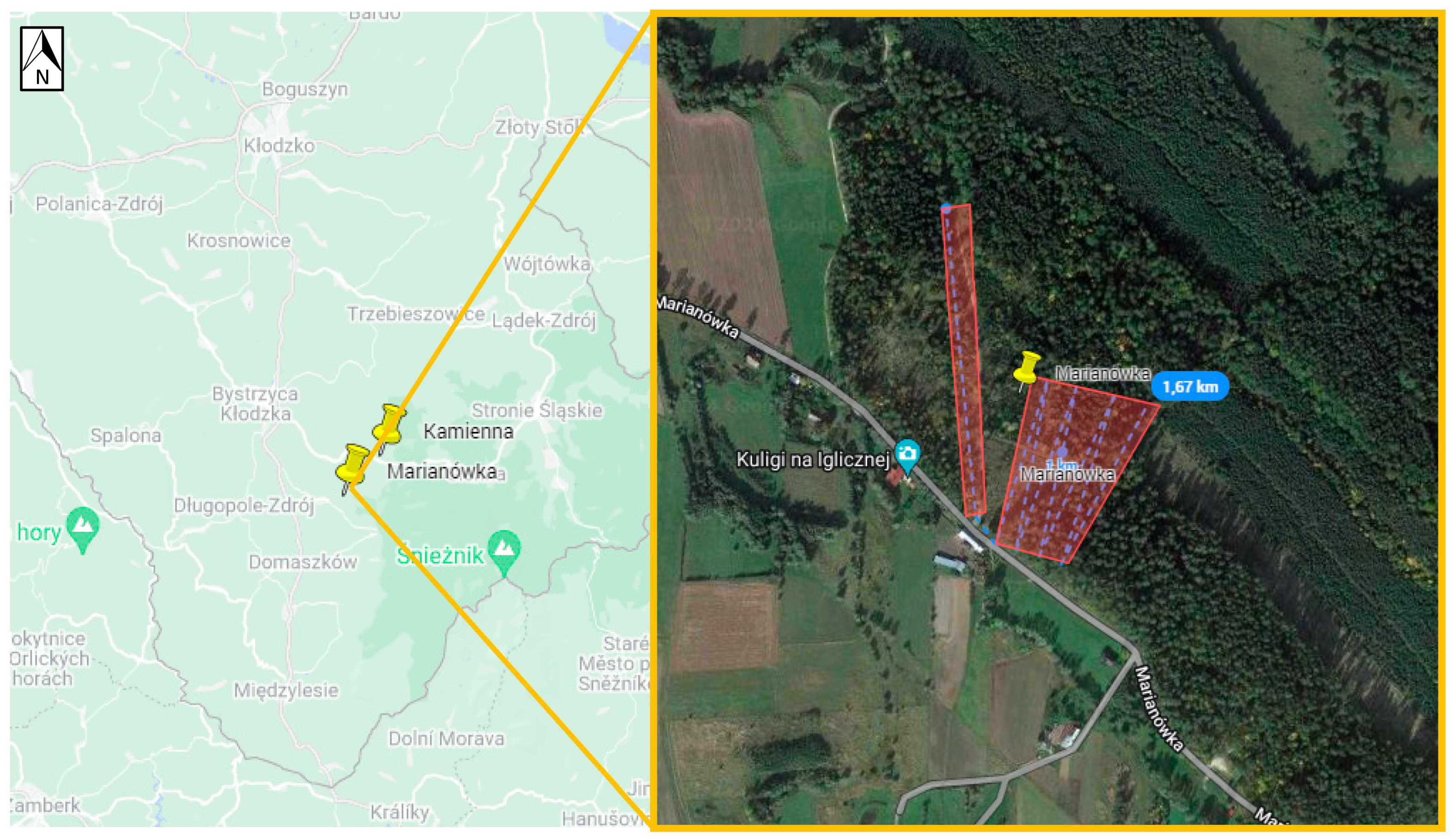
| Marianówka | 50.26388, 16.72314 | the forest close to the settlement | ||
| Dziewiętlin | 51.44850, 17.31076 | walking paths in the forest |
| Country | State/Voivodship | Name of Site | Number of Collected Ticks | Total | |||
|---|---|---|---|---|---|---|---|
| D. reticulatus | H. concinna | I. frontalis | I. ricinus | ||||
| Germany | Brandenburg | Cottbus | 0 | 0 | 0 | 76 | 76 |
| * Carl-Thiem-Klinikum Cottbus (from a patient) | 0 | 0 | 0 | 1 * | 1 * | ||
| Frauendorf | 3 | 0 | 0 | 223 | 226 | ||
| Geierwalder See | 18 | 0 | 0 | 34 | 52 | ||
| Deulowitzer See | 2 | 0 | 0 | 21 | 23 | ||
| Göhlensee | 10 | 2 | 0 | 620 | 632 | ||
| Heidesee | 0 | 0 | 0 | 62 | 62 | ||
| Löhsten | 646 | 7 | 0 | 16 | 669 | ||
| Pinnower See | 124 | 0 | 0 | 0 | 124 | ||
| Spremberg See | 6 | 0 | 0 | 33 | 39 | ||
| Mecklenburg-West Pomerania | Hiddensee Island | 0 | 0 | 0 | 205 | 205 | |
| Saxony | Michalken | 163 | 60 | 1 | 207 | 431 | |
| Senftenberger See | 77 | 6 | 1 | 195 | 279 | ||
| Sparte am Moor | 114 | 26 | 0 | 604 | 744 | ||
| Waldbad Bernsdorf | 278 | 4 | 0 | 344 | 626 | ||
| * Waldbad Bernsdorf (from a dog) | 41 * | 0 | 0 | 0 | 41 * | ||
| Subtotal | 1482 | 105 | 2 | 2641 | 4230 | ||
| Poland | Lubush | Jezioro Głębokie | 1928 | 2 | 0 | 67 | 1997 |
| Jeziory Wysokie | 58 | 1 | 0 | 76 | 135 | ||
| Lower Silesia | Kamienna | 0 | 0 | 0 | 123 | 123 | |
| Marianówka | 0 | 0 | 0 | 880 | 880 | ||
| Dziewiętlin | 38 | 0 | 0 | 997 | 1035 | ||
| Subtotal | 2024 | 3 | 0 | 2143 | 4170 | ||
| Total | 3506 | 108 | 2 | 4784 | 8400 | ||
| Country | State/Voivodship | Name of Site | Number of Positive Pools (MIR) | |||||
|---|---|---|---|---|---|---|---|---|
| D. reticulatus | I. ricinus | Total | ||||||
| Females | Males | Females | Males | Nymphs | ||||
| Germany | Brandenburg | Göhlensee | 0 | 0 | 5 (2.99%) | 1 (0.79%) | 0 | 6 (0.95%) |
| Löhsten | 2 (0.66%) | 4 (1.17%) | 0 | 1 (33.33%) | 0 | 7 (1.05%) | ||
| Saxony | Senftenberger See | 0 | 0 | 0 | 0 | 1 (0.62%) | 1 (0.36%) | |
| Sparte am Moor | 0 | 0 | 0 | 0 | 1 (0.18%) | 1 (0.13%) | ||
| Waldbad Bernsdorf | 0 | 0 | 1 (1.2%) | 0 | 1 (0.55%) | 2 (0.32%) | ||
| Subtotal | 2 (0.36%) | 4 (0.71%) | 6 (2.03%) | 2 (0.75%) | 3 (0.24%) | 17 (0.58%) | ||
| Poland | Lower Silesia | Marianówka | 0 | 0 | 1 (0.9%) | 0 | 1 (0.19%) | 2 (0.23%) |
| Total | 2 (0.36%) | 4 (0.71%) | 7 (1.76%) | 2 (0.54%) | 4 (0.23%) | 19 (0.5%) | ||
| Site | Number of Collected Ticks (Number of TBEV-Positive Pools) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D. reticulatus | H. concinna | I. ricinus | Total | ||||||||
| F | M | Subtotal | N | L | Subtotal | F | M | N | Subtotal | ||
| Löhsten 1 | 39 | 39 | 76 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 77 |
| Löhsten 2 | 22 | 22 | 44 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 45 |
| Löhsten 3 | 6 | 8 | 14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 |
| Löhsten 4 | 36 (1) | 53 | 89 (1) | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 90 (1) |
| Löhsten 5 | 6 | 6 | 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 |
| Löhsten 6 | 17 | 25 | 42 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 43 |
| Löhsten 7 | 18 | 28 | 46 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 47 |
| Löhsten 8 | 24 | 28 | 52 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 53 |
| Löhsten 9 | 12 | 5 | 17 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 18 |
| Löhsten 10 | 81 (1) | 78 (4) | 159 (5) | 0 | 0 | 0 | 0 | 1 (1) | 0 | 1 (1) | 160 (6) |
| Löhsten 11 | 0 | 3 | 3 | 3 | 0 | 3 | 0 | 0 | 4 | 4 | 10 |
| Löhsten 12 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 2 |
| Löhsten 13 | 33 | 40 | 73 | 2 | 0 | 2 | 0 | 0 | 5 | 5 | 80 |
| Löhsten 14 | 10 | 8 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 18 |
| Total | 303 (2) | 343 (4) | 646 (6) | 6 | 1 | 7 | 1 | 3 (1) | 12 | 16 (1) | 669 (7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Król, N.; Chitimia-Dobler, L.; Dobler, G.; Kiewra, D.; Czułowska, A.; Obiegala, A.; Zajkowska, J.; Juretzek, T.; Pfeffer, M. Identification of New Microfoci and Genetic Characterization of Tick-Borne Encephalitis Virus Isolates from Eastern Germany and Western Poland. Viruses 2024, 16, 637. https://doi.org/10.3390/v16040637
Król N, Chitimia-Dobler L, Dobler G, Kiewra D, Czułowska A, Obiegala A, Zajkowska J, Juretzek T, Pfeffer M. Identification of New Microfoci and Genetic Characterization of Tick-Borne Encephalitis Virus Isolates from Eastern Germany and Western Poland. Viruses. 2024; 16(4):637. https://doi.org/10.3390/v16040637
Chicago/Turabian StyleKról, Nina, Lidia Chitimia-Dobler, Gerhard Dobler, Dorota Kiewra, Aleksandra Czułowska, Anna Obiegala, Joanna Zajkowska, Thomas Juretzek, and Martin Pfeffer. 2024. "Identification of New Microfoci and Genetic Characterization of Tick-Borne Encephalitis Virus Isolates from Eastern Germany and Western Poland" Viruses 16, no. 4: 637. https://doi.org/10.3390/v16040637