The Human Antibody Response to Dengue Virus Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
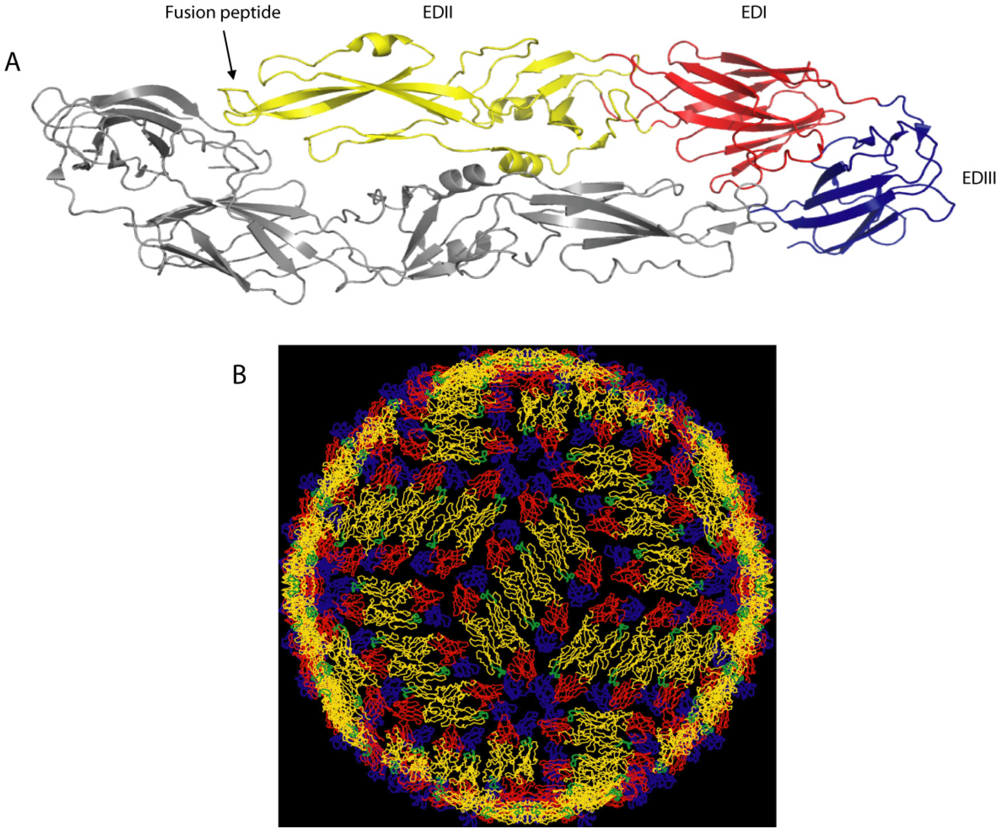
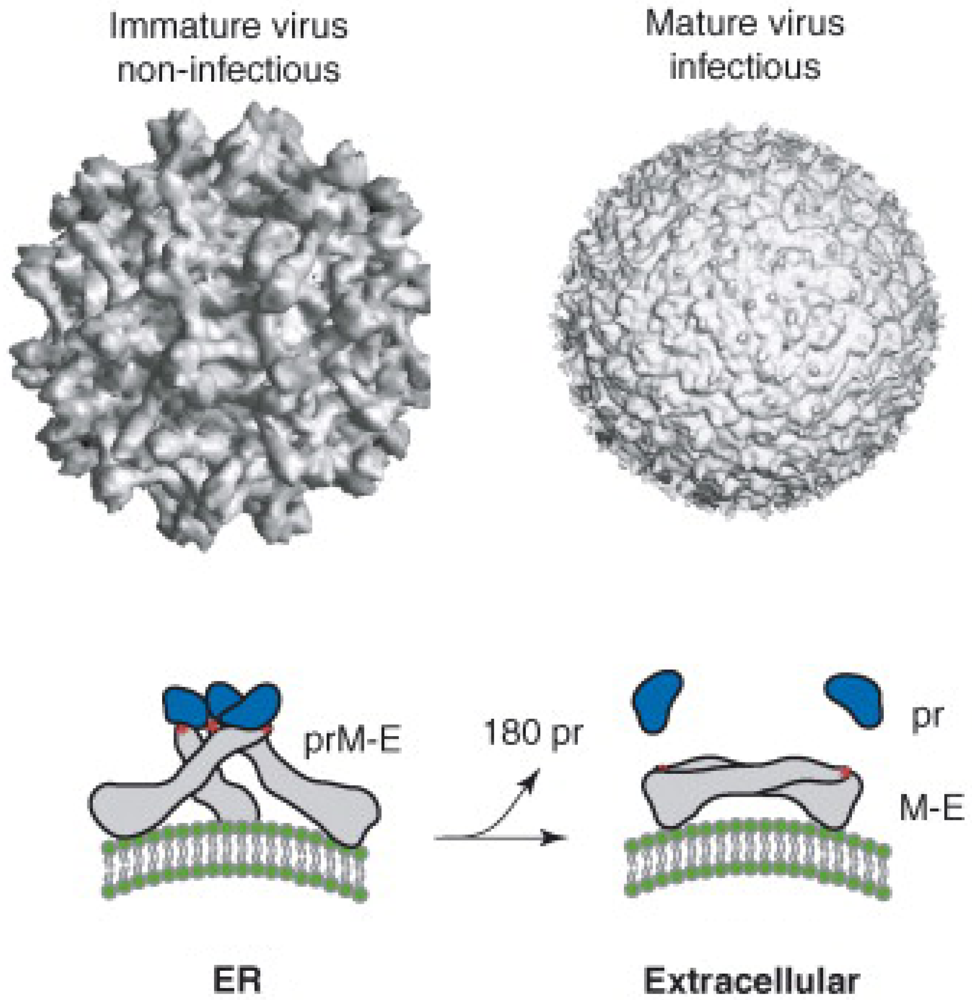
2. Dengue Virion Structure


3. Studies with Mouse MAbs and the Multi-Hit, Threshold Model of Dengue Neutralization

4. Human Antibody Response Following Natural DENV Infection
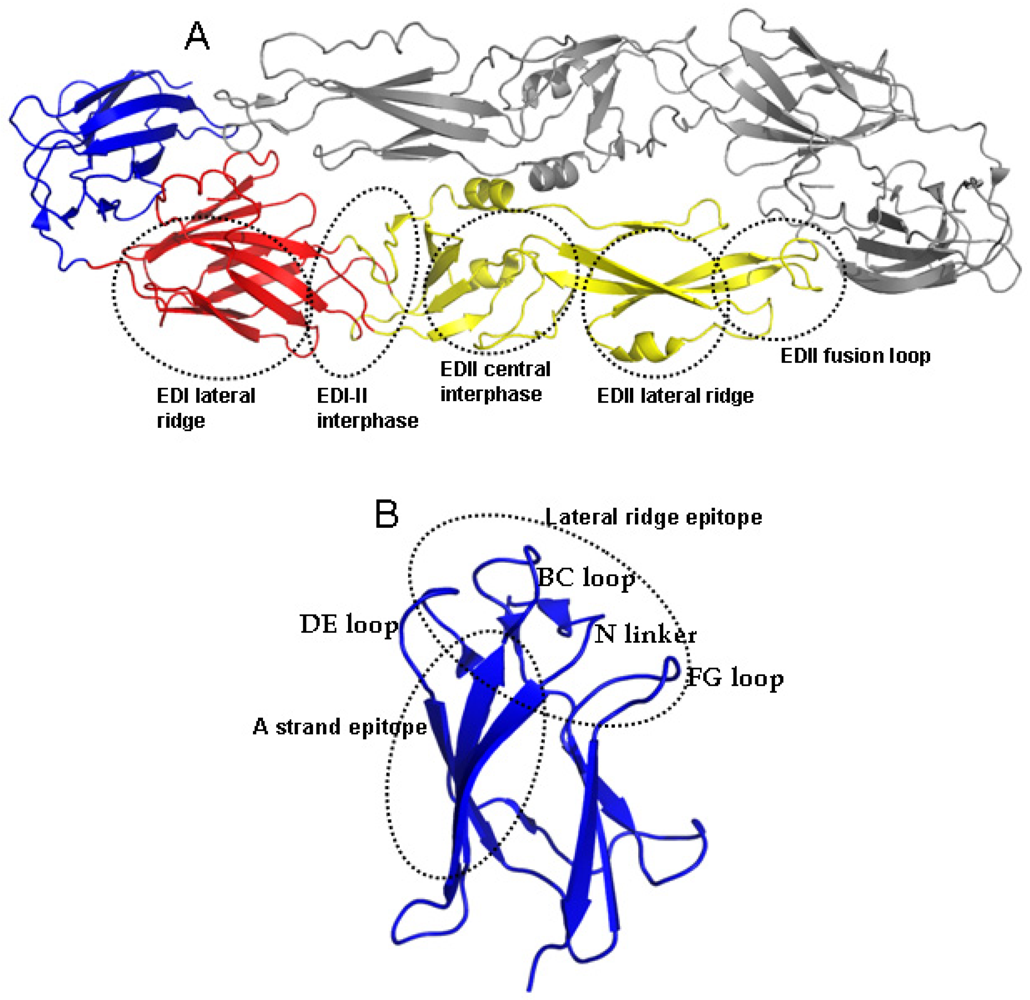
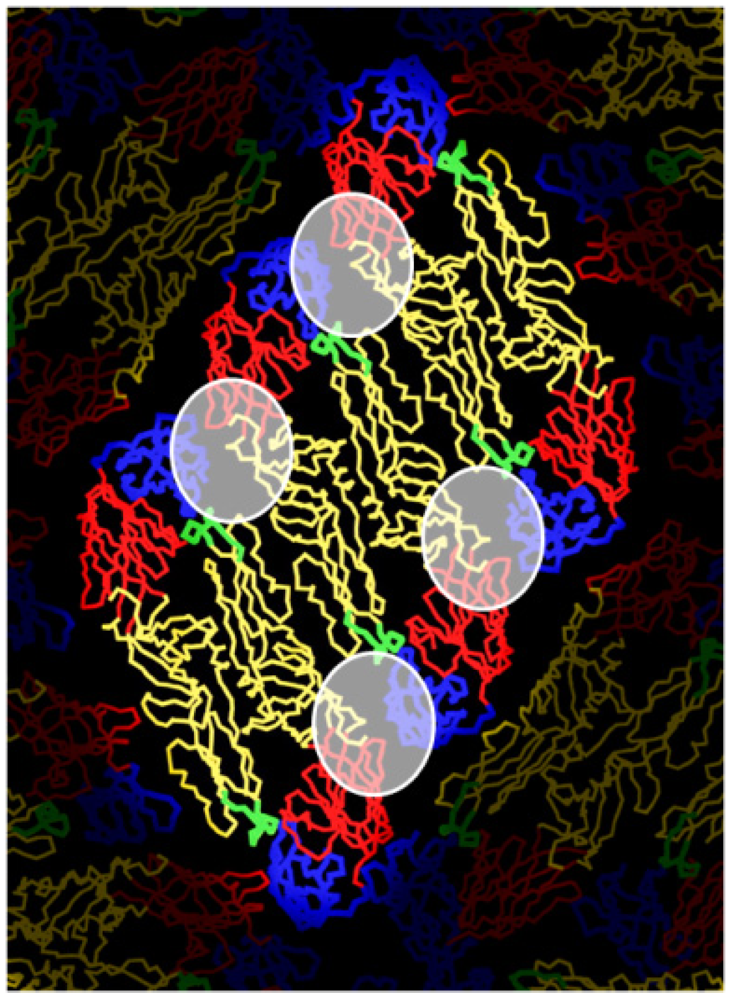
5. DENV Antigens and Epitopes Recognized by Human Antibodies
6. Profiling Antibody Response to Dengue Virus Using Human Monoclonal Antibodies

PrM Protein Is a Major Target of the Human Antibody Response
7. Intra-Serotype Strain Variation and DENV Neutralization
8. In Vitro versus in Vivo Neutralization
9. B-Cell Subsets Involved in the Humoral Response to DENV
10. Role of Antibodies in Enhancing DENV Infection and Disease
11. Properties of DENV Enhancing Antibodies
12. Summary
Notes Added in Proof
Acknowledgements
Conflict of Interest
References
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar]
- Roehrig, J.T. Antigenic structure of flavivirus proteins. Adv. Virus Res. 2003, 59, 141–175. [Google Scholar]
- Rothman, A.L. Immunology and immunopathogenesis of dengue disease. Adv. Virus Res. 2003, 60, 397–419. [Google Scholar]
- Halstead, S.B. Neutralization and antibody-dependent enhancement of dengue viruses. Adv. Virus Res. 2003, 60, 421–467. [Google Scholar]
- Kanai, R.; Kar, K.; Anthony, K.; Gould, L.H.; Ledizet, M.; Fikrig, E.; Marasco, W.A.; Koski, R.A.; Modis, Y. Crystal structure of West Nile virus envelope glycoprotein reveals viral surface epitopes. J. Virol. 2006, 80, 11000–11008. [Google Scholar]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 6986–6991. [Google Scholar]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. Variable surface epitopes in the crystal structure of dengue virus type 3 envelope glycoprotein. J. Virol. 2005, 79, 1223–1231. [Google Scholar]
- Nybakken, G.E.; Nelson, C.A.; Chen, B.R.; Diamond, M.S.; Fremont, D.H. Crystal structure of the West Nile virus envelope glycoprotein. J. Virol. 2006, 80, 11467–11474. [Google Scholar]
- Rey, F.A.; Heinz, F.X.; Mandl, C.; Kunz, C.; Harrison, S.C. The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature 1995, 375, 291–298. [Google Scholar]
- Zhang, Y.; Zhang, W.; Ogata, S.; Clements, D.; Strauss, J.H.; Baker, T.S.; Kuhn, R.J.; Rossmann, M.G. Conformational changes of the flavivirus E glycoprotein. Structure 2004, 12, 1607–1618. [Google Scholar]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of dengue virus: Implications for flavivirus organization, maturation, and fusion. Cell 2002, 108, 717–725. [Google Scholar] [PubMed]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the flavivirus life cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar]
- Kaufmann, B.R.; Nybakken, G.E.; Chipman, P.R.; Zhang, W.; Diamond, M.S.; Fremont, D.H.; Kuhn, R.J.; Rossmann, M.G. West Nile virus in complex with the Fab fragment of a neutralizing monoclonal antibody. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 12400–12404. [Google Scholar]
- Perera, R.; Khaliq, M.; Kuhn, R.J. Closing the door on flaviviruses: Entry as a target for antiviral drug design. Antivir. Res. 2008, 80, 11–22. [Google Scholar]
- Cherrier, M.V.; Kaufmann, B.; Nybakken, G.E.; Lok, S.M.; Warren, J.T.; Chen, B.R.; Nelson, C.A.; Kostyuchenko, V.A.; Holdaway, H.A.; Chipman, P.R.; et al. Structural basis for the preferential recognition of immature flaviviruses by a fusion-loop antibody. EMBO J. 2009, 28, 3269–3276. [Google Scholar] [PubMed]
- Junjhon, J.; Edwards, T.J.; Utaipat, U.; Bowman, V.D.; Holdaway, H.A.; Zhang, W.; Keelapang, P.; Puttikhunt, C.; Perera, R.; Chipman, P.R.; et al. Influence of pr-M cleavage on the heterogeneity of extracellular dengue virus particles. J. Virol. 2010, 84, 8353–8358. [Google Scholar] [PubMed]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science 2010, 328, 745–748. [Google Scholar] [PubMed]
- Rodenhuis-Zybert, I.A.; van der Schaar, H.M.; da Silva Voorham, J.M.; van der Ende-Metselaar, H.; Lei, H.Y.; Wilschut, J.; Smit, J.M. Immature dengue virus: A veiled pathogen? PLoS Pathog. 2010, 6, e1000718. [Google Scholar] [PubMed]
- Perera, R.; Kuhn, R.J. Structural proteomics of dengue virus. Curr. Opin. Microbiol. 2008, 11, 369–377. [Google Scholar]
- Pierson, T.C.; Diamond, M.S. Molecular mechanisms of antibody-mediated neutralisation of flavivirus infection. Expet. Rev. Mol. Med. 2008, 10, e12. [Google Scholar]
- Dowd, K.A.; Pierson, T.C. Antibody-mediated neutralization of flaviviruses: A reductionist view. Virology 2011, 411, 306–315. [Google Scholar]
- Wahala, W.M.; Donaldson, E.F.; de Alwis, R.; Accavitti-Loper, M.A.; Baric, R.S.; de Silva, A.M. Natural strain variation and antibody neutralization of dengue serotype 3 viruses. PLoS Pathog. 2010, 6, e1000821. [Google Scholar]
- Lok, S.-M.; Kostyuchenko, V.; Nybakken, G.E.; Holdaway, H.A.; Battisti, A.J.; Sukupolvi-Petty, S.; Sedlak, D.; Fremont, D.H.; Chipman, P.R.; Roehrig, J.T.; et al. Binding of a neutralizing antibody to dengue virus alters the arrangement of surface glycoproteins. Nat. Struct. Mol. Biol. 2008, 15, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Stiasny, K.; Brandler, S.; Kossl, C.; Heinz, F.X. Probing the flavivirus membrane fusion mechanism by using monoclonal antibodies. J. Virol. 2007, 81, 11526–11531. [Google Scholar]
- Thompson, B.S.; Moesker, B.; Smit, J.M.; Wilschut, J.; Diamond, M.S.; Fremont, D.H. A therapeutic antibody against West Nile virus neutralizes infection by blocking fusion within endosomes. PLoS Pathog. 2009, 5, e1000453. [Google Scholar]
- Chan, K.R.; Zhang, S.L.-X.; Tan, H.C.; Chan, Y.K.; Chow, A.; Lim, A.P.C.; Vasudevan, S.G.; Hanson, B.J.; Ooi, E.E. Ligation of Fc gamma receptor IIB inhibits antibody-dependent enhancement of dengue virus infection. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 12479–12484. [Google Scholar]
- Imrie, A.; Meeks, J.; Gurary, A.; Sukhbaatar, M.; Truong, T.T.; Cropp, C.B.; Effler, P. Antibody to dengue 1 detected more than 60 years after infection. Viral Immunol. 2007, 20, 672–675. [Google Scholar]
- Halstead, S.B.; Rojanasuphot, S.; Sangkawibha, N. Original antigenic sin in dengue. Am. J. Trop. Med. Hyg. 1983, 32, 154–156. [Google Scholar]
- Midgley, C.M.; Bajwa-Joseph, M.; Vasanawathana, S.; Limpitikul, W.; Wills, B.; Flanagan, A.; Waiyaiya, E.; Tran, H.B.; Cowper, A.E.; Chotiyarnwon, P.; et al. An in-depth analysis of original antigenic sin in dengue virus infection. J. Virol. 2011, 85, 410–421. [Google Scholar] [PubMed]
- Fish, S.; Zenowich, E.; Fleming, M.; Manser, T. Molecular analysis of original antigenic sin. I. Clonal selection, somatic mutation, and isotype switching during a memory B cell response. J. Exp. Med. 1989, 170, 1191–1209. [Google Scholar] [PubMed]
- Virelizier, J.L.; Allison, A.C.; Schild, G.C. Antibody responses to antigenic determinants of influenza virus hemagglutinin. II. Original antigenic sin: A bone marrow-derived lymphocyte memory phenomenon modulated by thymus-derived lymphocytes. J. Exp. Med. 1974, 140, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Kosasih, H.; Yusuf, H.; Sudjana, P.; Alisjahbana, B.; Wuryadic, S.; Akterono; Tan, R.I.; Yuwonoc, D.; Beckett, C.; Porter, K.R.; et al. Report of four volunteers with primary, secondary and tertiary dengue infections during a prospective cohort study. Dengue Bulletin 2006, 30, 87–92. [Google Scholar]
- Myers, R.M.; Varkey, M.J. A note on sequential dengue infection, presumptive and proved, with report of an instance of a third proved attack in one individual. Indian J. Med. Res. 1971, 59, 1231–1236. [Google Scholar] [PubMed]
- Koraka, P.; Suharti, C.; Setiati, T.E.; Mairuhu, A.T.; Van Gorp, E.; Hack, C.E.; Juffrie, M.; Sutaryo, J.; Van Der Meer, G.M.; Groen, J.; et al. Kinetics of dengue virus-specific serum immunoglobulin classes and subclasses correlate with clinical outcome of infection. J. Clin. Microbiol. 2001, 39, 4332–4338. [Google Scholar] [PubMed]
- Koraka, P.; Murgue, B.; Deparis, X.; Setiati, T.E.; Suharti, C.; van Gorp, E.C.; Hack, C.E.; Osterhaus, A.D.; Groen, J. Elevated levels of total and dengue virus-specific immunoglobulin E in patients with varying disease severity. J. Med. Virol. 2003, 70, 91–98. [Google Scholar]
- Thein, S.; Aaskov, J.; Myint, T.T.; Shwe, T.N.; Saw, T.T.; Zaw, A. Changes in levels of anti-dengue virus IgG subclasses in patients with disease of varying severity. J. Med. Virol. 1993, 40, 102–106. [Google Scholar]
- Watanaveeradej, V.; Endy, T.P.; Samakoses, R.; Kerdpanich, A.; Simasathien, S.; Polprasert, N.; Aree, C.; Vaughn, D.W.; Ho, C.; Nisalak, A. Transplacentally transferred maternal-infant antibodies to dengue virus. Am. J. Trop. Med. Hyg. 2003, 69, 123–128. [Google Scholar]
- Hofmeister, Y.; Planitzer, C.B.; Farcet, M.R.; Teschner, W.; Butterweck, H.A.; Weber, A.; Holzer, G.W.; Kreil, T.R. Human IgG Subclasses: In vitro neutralization of and in vivo protection against West Nile virus. J. Virol. 2011, 85, 1896–1899. [Google Scholar] [PubMed]
- Valdes, K.; Alvarez, M.; Pupo, M.; Vazquez, S.; Rodriguez, R.; Guzman, M.G. Human dengue antibodies against structural and nonstructural proteins. Clin. Diagn. Lab. Immunol. 2000, 7, 856–857. [Google Scholar]
- Churdboonchart, V.; Bhamarapravati, N.; Peampramprecha, S.; Sirinavin, S. Antibodies against dengue viral proteins in primary and secondary dengue hemorrhagic fever. Am. J. Trop. Med. Hyg. 1991, 44, 481–493. [Google Scholar]
- Lai, C.Y.; Tsai, W.Y.; Lin, S.R.; Kao, C.L.; Hu, H.P.; King, C.C.; Wu, H.C.; Chang, G.J.; Wang, W.K. Antibodies to envelope glycoprotein of dengue virus during the natural course of infection are predominantly cross-reactive and recognize epitopes containing highly conserved residues at the fusion loop of domain II. J. Virol. 2008, 82, 6631–6643. [Google Scholar]
- AbuBakar, S.; Azmi, A.; Mohamed-Saad, N.; Shafee, N.; Chee, H.Y. Antibody responses of dengue fever patients to dengue 2 (New Guinea C strain) viral proteins. Malays. J. Pathol. 1997, 19, 41–51. [Google Scholar]
- Abubakar, S.; Azila, A.; Suzana, M.; Chang, L.Y. Antigenic cell associated dengue 2 virus proteins detected in vitro using dengue fever patients sera. Malays. J. Pathol. 2002, 24, 29–36. [Google Scholar] [PubMed]
- Lazaro-Olan, L.; Mellado-Sanchez, G.; Garcia-Cordero, J.; Escobar-Gutierrez, A.; Santos-Argumedo, L.; Gutierrez-Castaneda, B.; Cedillo-Barron, L. Analysis of antibody response in human dengue patients from the Mexican coast using recombinant antigens. Vector Borne Zoonotic Dis. 2008, 8, 69–79. [Google Scholar]
- Falgout, B.; Bray, M.; Schlesinger, J.J.; Lai, C.J. Immunization of mice with recombinant vaccinia virus expressing authentic dengue virus nonstructural protein NS1 protects against lethal dengue virus encephalitis. J. Virol. 1990, 64, 4356–4363. [Google Scholar]
- Lin, C.-F.; Wan, S.-W.; Cheng, H.-J.; Lei, H.-Y.; Lin, Y.-S. Autoimmune pathogenesis in dengue virus infection. Viral Immunol. 2006, 19, 127–132. [Google Scholar]
- dos Santos, F.B.; Miagostovich, M.P.; Nogueira, R.M.; Schatzmayr, H.G.; Riley, L.W.; Harris, E. Analysis of recombinant dengue virus polypeptides for dengue diagnosis and evaluation of the humoral immune response. Am. J. Trop. Med. Hyg. 2004, 71, 144–152. [Google Scholar]
- Crill, W.D.; Hughes, H.R.; Delorey, M.J.; Chang, G.J. Humoral immune responses of dengue fever patients using epitope-specific serotype-2 virus-like particle antigens. PLoS One 2009, 4, e4991. [Google Scholar]
- Wahala, W.M.; Kraus, A.A.; Haymore, L.B.; Accavitti-Loper, M.A.; de Silva, A.M. Dengue virus neutralization by human immune sera: Role of envelope protein domain III-reactive antibody. Virology 2009, 392, 103–113. [Google Scholar]
- Oliphant, T.; Nybakken, G.E.; Austin, S.K.; Xu, Q.; Bramson, J.; Loeb, M.; Throsby, M.; Fremont, D.H.; Pierson, T.C.; Diamond, M.S. Induction of epitope-specific neutralizing antibodies against West Nile virus. J. Virol. 2007, 81, 11828–11839. [Google Scholar]
- Sánchez, M.D.; Pierson, T.C.; DeGrace, M.M.; Mattei, L.M.; Hanna, S.L.; Del Piero, F.; Doms, R.W. The neutralizing antibody response against West Nile virus in naturally infected horses. Virology 2007, 359, 336–348. [Google Scholar]
- de Alwis, R.; Beltramello, M.; Messer, W.B.; Sukupolvi-Petty, S.; Wahala, W.M.; Kraus, A.; Olivarez, N.P.; Pham, Q.; Brian, J.; Tsai, W.Y.; et al. In-depth analysis of the antibody response of individuals exposed to primary dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e1188. [Google Scholar] [PubMed]
- Beltramello, M.; Williams, K.L.; Simmons, C.P.; Macagno, A.; Simonelli, L.; Quyen, N.T.; Sukupolvi-Petty, S.; Navarro-Sanchez, E.; Young, P.R.; de Silva, A.M.; et al. The human immune response to Dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe 2010, 8, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Schieffelin, J.; Costin, J.; Nicholson, C.; Orgeron, N.; Fontaine, K.; Isern, S.; Michael, S.; Robinson, J. Neutralizing and non-neutralizing monoclonal antibodies against dengue virus E protein derived from a naturally infected patient. Virol. J. 2010, 7, 28. [Google Scholar]
- Traggiai, E.; Becker, S.; Subbarao, K.; Kolesnikova, L.; Uematsu, Y.; Gismondo, M.R.; Murphy, B.R.; Rappuoli, R.; Lanzavecchia, A. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nat. Med. 2004, 10, 871–875. [Google Scholar]
- Lai, C.J.; Goncalvez, A.P.; Men, R.; Wernly, C.; Donau, O.; Engle, R.E.; Purcell, R.H. Epitope determinants of a chimpanzee dengue virus type 4 (DENV-4)-neutralizing antibody and protection against DENV-4 challenge in mice and rhesus monkeys by passively transferred humanized antibody. J. Virol. 2007, 81, 12766–12774. [Google Scholar]
- Kaufmann, B.R.; Vogt, M.R.; Goudsmit, J.; Holdaway, H.A.; Aksyuk, A.A.; Chipman, P.R.; Kuhn, R.J.; Diamond, M.S.; Rossmann, M.G. Neutralization of West Nile virus by cross-linking of its surface proteins with Fab fragments of the human monoclonal antibody CR4354. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 18950–18955. [Google Scholar]
- Vogt, M.R.; Moesker, B.; Goudsmit, J.; Jongeneelen, M.; Austin, S.K.; Oliphant, T.; Nelson, S.; Pierson, T.C.; Wilschut, J.; Throsby, M.; et al. Human monoclonal antibodies against West Nile virus induced by natural infection neutralize at a postattachment step. J. Virol. 2009, 83, 6494–6507. [Google Scholar] [PubMed]
- Purdue News. Scientists solve structure of dengue virus. Available online: http://www.purdue.edu/uns/images/kuhn.dengue1.jpeg (accessed on 1 September 2011).
- Rico-Hesse, R. Microevolution and virulence of dengue viruses. Adv. Virus Res. 2003, 59, 315–341. [Google Scholar]
- Holmes, E.C. RNA virus genomics: A world of possibilities. J. Clin. Invest. 2009, 119, 2488–2495. [Google Scholar]
- Blaney, J.E., Jr.; Matro, J.M.; Murphy, B.R.; Whitehead, S.S. Recombinant, live-attenuated tetravalent dengue virus vaccine formulations induce a balanced, broad, and protective neutralizing antibody response against each of the four serotypes in rhesus monkeys. J. Virol. 2005, 79, 5516–5528. [Google Scholar] [PubMed]
- Endy, T.P.; Nisalak, A.; Chunsuttitwat, S.; Vaughn, D.W.; Green, S.; Ennis, F.A.; Rothman, A.L.; Libraty, D.H. Relationship of preexisting dengue virus (DV) neutralizing antibody levels to viremia and severity of disease in a prospective cohort study of DV infection in Thailand. J. Infect. Dis. 2004, 189, 990–1000. [Google Scholar]
- Zulueta, A.; Martín, J.; Hermida, L.; Alvarez, M.; Valdés, I.; Prado, I.; Chinea, G.; Rosario, D.; Guillén, G.; Guzmán, M.G. Amino acid changes in the recombinant Dengue 3 Envelope domain III determine its antigenicity and immunogenicity in mice. Virus Res. 2006, 121, 65–73. [Google Scholar]
- Li, L.; Barrett, A.D.T.; Beasley, D.W.C. Differential expression of domain III neutralizing epitopes on the envelope proteins of West Nile virus strains. Virology 2005, 335, 99–105. [Google Scholar]
- Sánchez, M.D.; Pierson, T.C.; McAllister, D.; Hanna, S.L.; Puffer, B.A.; Valentine, L.E.; Murtadha, M.M.; Hoxie, J.A.; Doms, R.W. Characterization of neutralizing antibodies to West Nile virus. Virology 2005, 336, 70–82. [Google Scholar]
- Brien, J.D.; Austin, S.K.; Sukupolvi-Petty, S.; O'Brien, K.M.; Johnson, S.; Fremont, D.H.; Diamond, M.S. Genotype-specific neutralization and protection by antibodies against dengue virus type 3. J. Virol. 2010, 84, 10630–10643. [Google Scholar]
- Shrestha, B.; Brien, J.D.; Sukupolvi-Petty, S.; Austin, S.K.; Edeling, M.A.; Kim, T.; O'Brien, K.M.; Nelson, C.A.; Johnson, S.; Fremont, D.H.; et al. The development of therapeutic antibodies that neutralize homologous and heterologous genotypes of dengue virus type 1. PLoS Pathog. 2010, 6, e1000823. [Google Scholar] [PubMed]
- Sukupolvi-Petty, S.; Austin, S.K.; Engle, M.; Brien, J.D.; Dowd, K.A.; Williams, K.L.; Johnson, S.; Rico-Hesse, R.; Harris, E.; Pierson, T.C.; et al. Structure and function analysis of therapeutic monoclonal antibodies against dengue virus type 2. J. Virol. 2010, 84, 9227–9239. [Google Scholar] [PubMed]
- Messer, W.B.; Yount, B.; Hacker, K.E.; Donaldson, E.F.; Huynh, J.P.; de Silva, A.M.; Baric, R.S. Development and characterization of a reverse genetic system for studying dengue virus serotype 3 strain variation and neutralization. PLoS Negl. Trop. Dis. 2011. submitted for publication. [Google Scholar]
- Dunn, M.; Rossi, S.; Carter, D.; Vogt, M.; Mehlhop, E.; Diamond, M.; Ross, T. Enhancement of anti-DIII antibodies by the C3d derivative P28 results in lower viral titers and augments protection in mice. Virol. J. 2010, 7, 95. [Google Scholar]
- Mehlhop, E.; Nelson, S.; Jost, C.A.; Gorlatov, S.; Johnson, S.; Fremont, D.H.; Diamond, M.S.; Pierson, T.C. Complement protein C1q reduces the stoichiometric threshold for antibody-mediated neutralization of West Nile virus. Cell Host Microbe 2009, 6, 381–391. [Google Scholar]
- Mehlhop, E.; Ansarah-Sobrinho, C.; Johnson, S.; Engle, M.; Fremont, D.H.; Pierson, T.C.; Diamond, M.S. Complement protein C1q inhibits antibody-dependent enhancement of flavivirus infection in an IgG subclass-specific manner. Cell Host Microbe 2007, 2, 417–426. [Google Scholar]
- Mehlhop, E.; Fuchs, A.; Engle, M.; Diamond, M.S. Complement modulates pathogenesis and antibody-dependent neutralization of West Nile virus infection through a C5-independent mechanism. Virology 2009, 393, 11–15. [Google Scholar]
- Avirutnan, P.; Mehlhop, E.; Diamond, M.S. Complement and its role in protection and pathogenesis of flavivirus infections. Vaccine 2008, 26, I100–I107. [Google Scholar]
- Halstead, S.B.; Mahalingam, S.; Marovich, M.A.; Ubol, S.; Mosser, D.M. Intrinsic antibody-dependent enhancement of microbial infection in macrophages: disease regulation by immune complexes. Lancet Infect. Dis. 2010, 10, 712–722. [Google Scholar]
- Laoprasopwattana, K.; Libraty, D.H.; Endy, T.P.; Nisalak, A.; Chunsuttiwat, S.; Ennis, F.A.; Rothman, A.L.; Green, S. Antibody-dependent cellular cytotoxity mediated by plasma obtained before secondary dengue virus infections: Potential involvement in early control of viral replication. J. Infect. Dis. 2007, 195, 1108–1116. [Google Scholar]
- Kurane, I.; Hebblewaite, D.; Brandt, W.E.; Ennis, F.A. Lysis of dengue virus-infected cells by natural cell-mediated cytotoxicity and antibody-dependent cell-mediated cytotoxicity. J. Virol. 1984, 52, 223–230. [Google Scholar]
- Kyle, J.L.; Balsitis, S.J.; Zhang, L.; Beatty, P.R.; Harris, E. Antibodies play a greater role than immune cells in heterologous protection against secondary dengue virus infection in a mouse model. Virology 2008, 380, 296–303. [Google Scholar]
- Libraty, D.H.; Acosta, L.P.; Tallo, V.; Segubre-Mercado, E.; Bautista, A.; Potts, J.A.; Jarman, R.G.; Yoon, I.K.; Gibbons, R.V.; Brion, J.D.; et al. A prospective nested case-control study of Dengue in infants: Rethinking and refining the antibody-dependent enhancement dengue hemorrhagic fever model. PLoS Med. 2009, 6, e1000171. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, B.M.; Summers, P.L.; Dubois, D.R.; Cohen, W.H.; Gentry, M.K.; Timchak, R.L.; Burke, D.S.; Eckels, K.H. Monoclonal antibodies for dengue virus prM glycoprotein protect mice against lethal dengue infection. Am. J. Trop. Med. Hyg. 1989, 41, 576–580. [Google Scholar]
- Vogt, M.R.; Dowd, K.A.; Engle, M.; Tesh, R.B.; Johnson, S.; Pierson, T.C.; Diamond, M.S. Poorly neutralizing cross-reactive antibodies against the fusion loop of West Nile virus envelope protein protect in vivo via Fc-{gamma} receptor and complement-dependent effector mechanisms. J. Virol. 2011.
- Baumgarth, N.C.Y.; Rothaeusler, K.; Yang, Y.; Herzenberg, L.A. B cell lineage contributions to antiviral host responses. Curr. Top. Microbiol. Immunol. 2008, 319, 41–61. [Google Scholar]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar]
- Kliks, S.C.; Nimmanitya, S.; Nisalak, A.; Burke, D.S. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am. J. Trop. Med. Hyg. 1988, 38, 411–419. [Google Scholar]
- Chau, T.N.; Hieu, N.T.; Anders, K.L.; Wolbers, M.; Lien le, B.; Hieu, L.T.; Hien, T.T.; Hung, N.T.; Farrar, J.; Whitehead, S.; et al. Dengue virus infections and maternal antibody decay in a prospective birth cohort study of Vietnamese infants. J. Infect. Dis. 2009, 200, 1893–1900. [Google Scholar] [PubMed]
- Chau, T.N.; Quyen, N.T.; Thuy, T.T.; Tuan, N.M.; Hoang, D.M.; Dung, N.T.; Lien le, B.; Quy, N.T.; Hieu, N.T.; Hieu, L.T.; et al. Dengue in Vietnamese infants—Results of infection-enhancement assays correlate with age-related disease epidemiology, and cellular immune responses correlate with disease severity. J. Infect. Dis. 2008, 198, 516–524. [Google Scholar] [PubMed]
- Kliks, S.C.; Nisalak, A.; Brandt, W.E.; Wahl, L.; Burke, D.S. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am. J. Trop. Med. Hyg. 1989, 40, 444–451. [Google Scholar]
- Laoprasopwattana, K.; Libraty, D.H.; Endy, T.P.; Nisalak, A.; Chunsuttiwat, S.; Vaughn, D.W.; Reed, G.; Ennis, F.A.; Rothman, A.L.; Green, S. Dengue virus (DV) enhancing antibody activity in preillness plasma does not predict subsequent disease severity or viremia in secondary dv infection. J. Infect. Dis. 2005, 192, 510–519. [Google Scholar]
- Halstead, S.B.; Shotwell, H.; Casals, J. Studies on the pathogenesis of Dengue infection in monkeys. II. Clinical laboratory responses to heterologous infection. J. Infect. Dis. 1973, 128, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Goncalvez, A.P.; Engle, R.E.; St.Claire, M.; Purcell, R.H.; Lai, C.-J. Monoclonal antibody-mediated enhancement of dengue virus infection in vitro and in vivo and strategies for prevention. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 9422–9427. [Google Scholar] [PubMed]
- Yauch, L.E.; Shresta, S. Mouse models of dengue virus infection and disease. Antivir. Res. 2008, 80, 87–93. [Google Scholar]
- Zellweger, R.M.; Prestwood, T.R.; Shresta, S. Enhanced infection of liver sinusoidal endothelial cells in a mouse model of antibody-induced severe dengue disease. Cell Host Microbe 2010, 7, 128–139. [Google Scholar]
- Balsitis, S.J.; Williams, K.L.; Lachica, R.; Flores, D.; Kyle, J.L.; Mehlhop, E.; Johnson, S.; Diamond, M.S.; Beatty, P.R.; Harris, E. Lethal antibody enhancement of dengue disease in mice is prevented by Fc modification. PLoS Pathog. 2010, 6, e1000790. [Google Scholar]
- Ubol, S.; Halstead, S.B. How innate immune mechanisms contribute to antibody-enhanced viral infections. Clin. Vaccine Immunol. 2010, 17, 1829–1835. [Google Scholar]
- Chareonsirisuthigul, T.; Kalayanarooj, S.; Ubol, S. Dengue virus (DENV) antibody-dependent enhancement of infection upregulates the production of anti-inflammatory cytokines, but suppresses anti-DENV free radical and pro-inflammatory cytokine production, in THP-1 cells. J. Gen. Virol. 2007, 88, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Ubol, S.; Phuklia, W.; Kalayanarooj, S.; Modhiran, N. Mechanisms of immune evasion induced by a complex of dengue virus and preexisting enhancing antibodies. J. Infect. Dis. 2010, 201, 923–935. [Google Scholar]
- Boonnak, K.; Dambach, K.M.; Donofrio, G.C.; Tassaneetrithep, B.; Marovich, M.A. Cell type specificity and host genetic polymorphisms influence antibody-dependent enhancement of dengue virus infection. J. Virol. 2011, 85, 1671–1683. [Google Scholar]
- Kou, Z.; Lim, J.Y.H.; Beltramello, M.; Quinn, M.; Chen, H.; Liu, S.-n.; Martnez-Sobrido, L.; Diamond, M.S.; Schlesinger, J.J.; de Silva, A.; et al. Human antibodies against dengue enhance dengue viral infectivity without suppressing type I interferon secretion in primary human monocytes. Virology 2011, 410, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Rolph, M.S.; Zaid, A.; Rulli, N.E.; Mahalingam, S. Downregulation of interferon-β in antibody-dependent enhancement of dengue viral infections of human macrophages is dependent on interleukin-6. J. Infect. Dis. 2011, 204, 489–491. [Google Scholar] [PubMed]
- Colpitts, T.M.; Rodenhuis-Zybert, I.; Moesker, B.; Wang, P.; Fikrig, E.; Smit, J. prM-antibody renders immature West Nile virus infectious in vivo. J. Gen. Virol. 2011, 92, 2281–2285. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; West, K.; Kalayanarooj, S.; Gibbons, R.V.; Srikiatkhachorn, A.; Green, S.; Libraty, D.; Jaiswal, S.; Rothman, A.L. B-cell responses during primary and secondary dengue virus infections in humans. J. Infect. Dis. 2011, 204, 1514–1522. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wahala, W.M.P.B.; De Silva, A.M. The Human Antibody Response to Dengue Virus Infection. Viruses 2011, 3, 2374-2395. https://doi.org/10.3390/v3122374
Wahala WMPB, De Silva AM. The Human Antibody Response to Dengue Virus Infection. Viruses. 2011; 3(12):2374-2395. https://doi.org/10.3390/v3122374
Chicago/Turabian StyleWahala, Wahala M. P. B., and Aravinda M. De Silva. 2011. "The Human Antibody Response to Dengue Virus Infection" Viruses 3, no. 12: 2374-2395. https://doi.org/10.3390/v3122374