Mechanisms and Factors that Influence High Frequency Retroviral Recombination
by
Krista Delviks-Frankenberry
1, 
Andrea Galli
2,3,
Olga Nikolaitchik
2,
Helene Mens
4,
Vinay K. Pathak
1 and
Wei-Shau Hu
2,* 1
Viral Mutation Section, HIV Drug Resistance Program, National Cancer Institute at Frederick, Frederick, MD 21702, USA
2
Viral Recombination Section, HIV Drug Resistance Program, National Cancer Institute at Frederick, Frederick, MD 21702, USA
3
Copenhagen Hepatitis C Program, Department of Infectious Diseases, Copenhagen University Hospital, Hvidovre 2650, Denmark
4
Department of Epidemic Diseases, Rigshospitalet, København 2100, Denmark
*
Author to whom correspondence should be addressed.
Viruses 2011, 3(9), 1650-1680; https://doi.org/10.3390/v3091650
Submission received: 21 June 2011
/
Revised: 18 August 2011
/
Accepted: 25 August 2011
/
Published: 9 September 2011
(This article belongs to the Special Issue Recombination in Viruses)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:With constantly changing environmental selection pressures, retroviruses rely upon recombination to reassort polymorphisms in their genomes and increase genetic diversity, which improves the chances for the survival of their population. Recombination occurs during DNA synthesis, whereby reverse transcriptase undergoes template switching events between the two copackaged RNAs, resulting in a viral recombinant with portions of the genetic information from each parental RNA. This review summarizes our current understanding of the factors and mechanisms influencing retroviral recombination, fidelity of the recombination process, and evaluates the subsequent viral diversity and fitness of the progeny recombinant. Specifically, the high mutation rates and high recombination frequencies of HIV-1 will be analyzed for their roles in influencing HIV-1 global diversity, as well as HIV-1 diagnosis, drug treatment, and vaccine development.
1. Introduction
Retroviridae is a family of RNA viruses that include an integrated DNA phase in their replication cycle. Several members of this family are viral human pathogens such as human immunodeficiency virus type 1 and type 2 (HIV-1 and HIV-2). One of the hallmarks of retroviral replication is the high frequency of recombination that can assort mutations in the viral genome to increase viral population diversity. High variation increases the probability of the population to survive, as it is likely that some proportion of the viral population will be able to escape the changing environmental selection pressures.
Most retroviruses exhibit very high population diversity; one of the best examples is the variation observed with HIV-1. HIV-1 can be divided into four groups, M, O, N, and P; however, 95% of the HIV-1 infections worldwide are caused by group M. Although group M virus was first transmitted into the human population by a single zoonotic event from SIVcpz, a simian immunodeficiency virus that infects chimpanzees [1], group M has diverged tremendously in the human population and therefore can be further divided based on sequence identity into several subtypes (A, B, C, D, F, G, H, J, and K) [2] and intersubtype recombinants. Different group M subtypes vary in their prevalence and distribution; for example, subtype C variants are the most widespread, accounting for 48% of all HIV-1 infections globally, whereas most HIV-1 infections in the Americas and Western Europe are caused by subtype B variants [3]. Generally, variants within the same subtype have more than 90% sequence identity whereas variants from different subtypes have between 80–90% sequence identity in their genomes. One of the major mechanisms generating this high diversity is frequent recombination, which can quickly distribute existing sequences and generate variants with different assortments of polymorphisms in the viral population. Recombination can occur within the same subtype, known as intrasubtype recombination, or between different subtypes, known as intersubtype recombination. Recombination can also occur between different HIV groups, albeit rarely. This review focuses on the mechanisms and factors that affect retroviral recombination, and how recombination influences HIV-1 diagnosis, treatment, and vaccine development.
1.1. Retroviral Replication Cycle
The detailed mechanisms of retrovirus replication are complex and involve intricate interactions with the host cell machinery at multiple steps [4]. Retroviruses package two copies of full-length viral RNAs into each particle, and each mature particle contains three virally encoded enzymes: protease, reverse transcriptase (RT) and integrase. RT uses the viral RNA as a template to synthesize a double-stranded DNA copy of the viral genome. This process, known as reverse transcription, occurs in the target cells for all retroviruses except spumaviruses [5]. The newly synthesized viral DNA integrates into the host cell genome, a step that is mediated by the viral enzyme integrase, to form a provirus. Host cell machinery transcribes the provirus to generate viral RNAs; some of the viral RNAs are spliced whereas others remain full-length. Viral RNAs are then exported out of the nucleus and used as templates for the translation of viral proteins. Newly synthesized viral proteins, together with two copies of the full-length viral RNA, assemble into viral particles to be released from the host cell.
1.2. Reverse Transcription of the Viral RNA into DNA
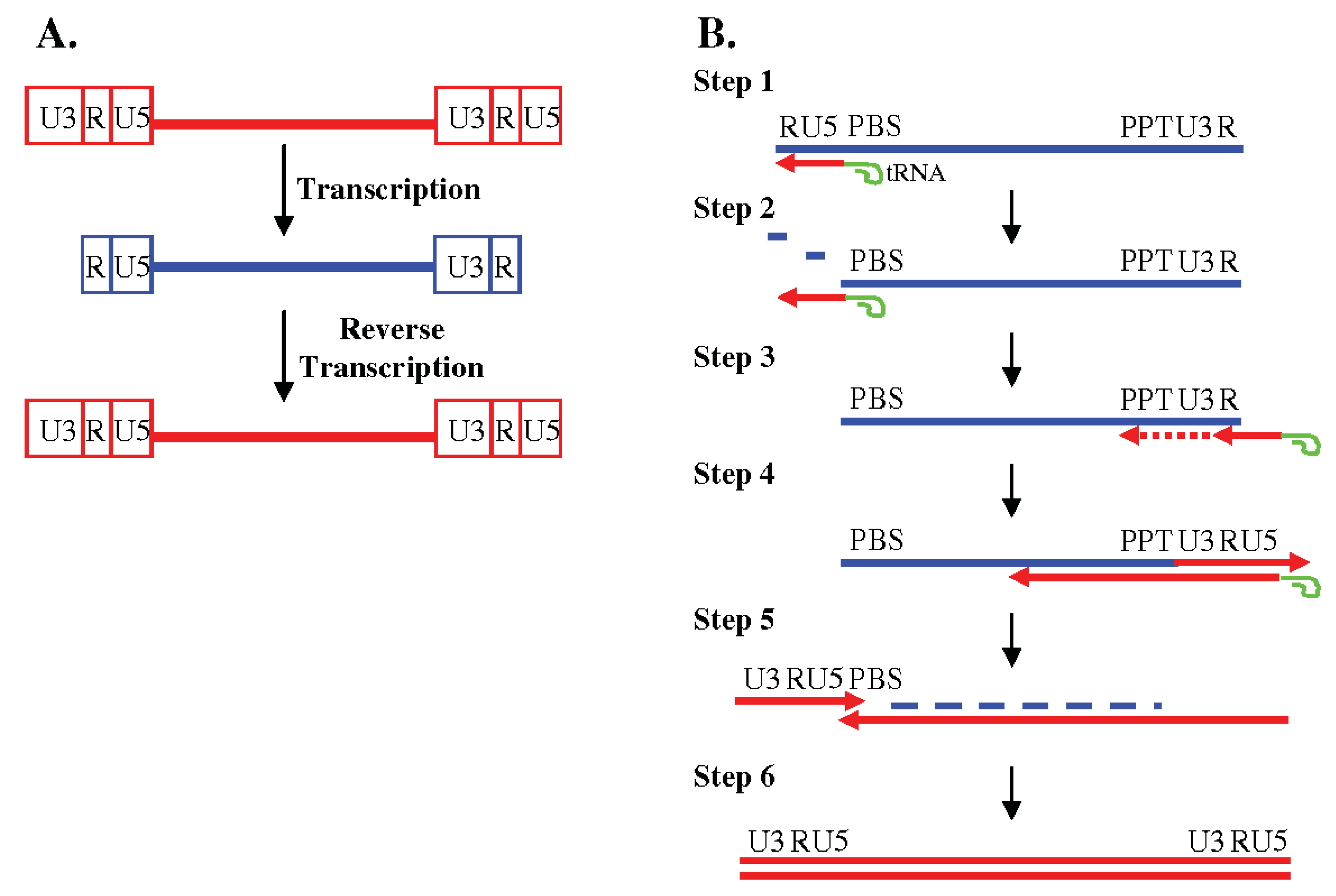
Although RT is the only enzyme required for synthesizing DNA from an RNA template in vitro, reverse transcription of the viral genome in vivo is connected to many other steps in early viral infection such as uncoating [6]. Figure 1 shows the complicated, but elegant process of reverse transcription [4]. A provirus generally has two identical blocks of sequences at each end known as long terminal repeats (LTRs) (Figure 1A). LTRs can be further divided into three regions, unique 3' region (U3), repeat (R), and unique 5' region (U5). As shown in Figure 1A, the sequence of the viral RNA is shorter than the proviral DNA, lacking both the 5' U3 region and the 3' U5 region. Since the viral promoter is located in the 5' U3 region, the expression of the viral DNA is dependent upon the process of reverse transcription to reconstitute full-length LTRs.
Reverse transcription involves at least two obligatory template switching events, namely, minus-strand and plus-strand DNA transfer (Figure 1B). Reverse transcription initiates near the 5' end of the viral RNA using a host cell tRNA primer, which is partially annealed to a region of viral RNA based on complementarity, termed the primer binding site (PBS) (Step 1). Since the viral RNA packaged into the virion is plus sense, the first strand of DNA synthesized is referred to as the minus strand. RT, which is also a DNA polymerase, copies a short stretch of the viral RNA including the 5' U5 and 5' R region. In addition to the DNA polymerase activity, RT also has RNase H activity, which specifically degrades RNA in an RNA:DNA hybrid. Degradation of the RNA that was used as template to synthesize the minus-strand DNA, exposes the newly synthesized 5' R region (Step 2), which can form base-pairing with the R region at the 3' end of the viral genome, completing the first template-switching event known as minus-strand DNA transfer. It has been shown that during this step, the minus-strand DNA can transfer either to the same RNA molecule (intramolecular transfer) or to the copackaged RNA (intermolecular transfer) [7,8,9]. RT then continues copying the U3 located at the 3' end of the viral RNA (Step 3). Once minus-strand DNA synthesis continues past a region called the polypurine tract (PPT), which is resistant to RNase H cleavage, RT makes a specific cleavage between the PPT and 3' U3. RT uses the PPT as a primer to initiate plus-strand DNA synthesis and copies the 3' LTR (U3, R and U5) along with a portion of the tRNA primer (Step 4). The plus-strand DNA synthesis in many retroviruses is discontinuous and can initiate from more than one position; some viruses, such as HIV-1, contains a second PPT located in the middle of the genome termed central PPT. Using the short complementarity between the plus-strand DNA and the PBS region on the minus-strand DNA, RT makes a second obligatory switch, termed plus-strand DNA transfer (Step 5); although shown as a linear model, it is likely that the two ends of the nucleic acid complex are in close proximity during plus-strand DNA transfer. Both plus-strand and minus-strand DNA synthesis then continue to complete reverse transcription (Step 6).
1.3. Retroviral Recombination Models
In order to complete reverse transcription, RT must undergo two obligatory template switches. Based on the natural ability of RT to switch templates, a model for retroviral recombination was proposed known as the forced copy choice model [10]. This model proposes that due to the presence of breaks found in the packaged viral RNA, RT will switch to the copackaged RNA when it encounters a break in the RNA, and use it as a template for DNA synthesis. As a result, a recombinant DNA molecule will be generated that contains portions of the genetic information from each copackaged RNA. This model was later modified to be known as the dynamic copy choice model [11], which proposes that template switching is influenced by a balance between the polymerase and RNase H activity of RT; during minus-strand DNA synthesis, polymerase activity synthesizes the DNA whereas RNase H activity degrades the RNA in the RNA:DNA hybrid. This process is dynamic, and when the newly synthesized DNA in the RNA:DNA hybrid becomes exposed, it is more likely to switch to the other copackaged RNA and use it as a template for DNA synthesis. This model is described in more detail in Section 2.
The forced copy choice and dynamic copy choice models describe recombination that occurs during minus-strand DNA synthesis; the strand displacement assimilation model has also been proposed to depict recombination during plus-strand DNA synthesis [12]. This model was based upon the observation that in avian retroviruses, such as avian sarcoma leukosis virus, plus-strand DNA synthesis is discontinuous and synthesized in many pieces. This model posits that some of the short pieces of plus-strand DNA can dissociate and anneal to the minus-strand DNA synthesized using the copackaged RNA as a template. After DNA repair that preserves the genetic information on the plus-strand DNA, a recombinant can be generated.
1.4. Requirements for the Generation of Observable Recombination Events
The high frequency of retroviral recombination occurs during reverse transcription of the copackaged viral RNA, and not during coinfection between two different viruses [13]. A dually-infected cell that contains two proviruses can generate viral particles that contain two copies of RNA from parent 1 or two copies of RNAs from parent 2 (homozygous particles), or one copy of RNA from parent 1 and one copy of RNA from parent 2 (heterozygous particles). Although recombination can occur in both homozygous and heterozygous particles, only the progeny of heterozygous particles can generate a recombinant that is genotypically different than the two parents [13]. Therefore, multiple requirements must be met to generate a recombination event that can be identified: (1) a cell must be infected by more than one virus; (2) both proviruses must express their genomes; (3) RNAs from the coinfected proviruses must be copackaged; (4) RT must switch templates between the copackaged RNAs to generate a recombinant DNA copy; and (5) the recombinant DNA integrates and expresses its genome. Although multiple steps are required to generate a recombinant, retroviruses still have one of the highest recombination rates. The impact of this high recombination potential can be demonstrated in the current HIV-1 epidemic as a large proportion of the currently circulating strains of HIV-1 are known recombinants.
2. Template Switching During Reverse Transcription
Reverse transcription of the retroviral genome requires RT to undergo two obligatory template switching (strand transfer) events to complete viral DNA synthesis. This requirement leads to the proposal that RT evolved to select for low template affinity and low processivity in order to complete reverse transcription. RT is estimated to dissociate from the template during reverse transcription at least 8 times for a 10-kb viral genome [14]. Specifically for HIV-1, it is estimated that 3–12 template switches occur per genome per replication cycle [15,16,17]. Since retroviruses package two copies of their genome into the viral particle, template switches that occur can either be intermolecular, resulting in homologous or nonhomologous recombination [18,19,20], or intramolecular, resulting in deletions, deletions with insertions, and insertions and duplications [14,21,22]. However, if RT reassociates back onto the same complex at the same position, then no recombination occurs. In this section, we will discuss factors that influence template switching, as measured through in vivo direct repeat deletions and in vitro biochemical studies, in simple retroviruses such as spleen necrosis virus (SNV) and murine leukemia virus (MLV) as well as complex retroviruses such as HIV-1.
2.1. In Vivo Direct Repeat Deletion Reveals that Template Switching Occurs Frequently During Reverse Transcription and is Dependent Upon the Size and Distance Between the Direct Repeats
Since the early 1980s it has been reported that directly repeated sequences in both viral genomes and viral vectors are unstable [23,24,25,26]. Specifically, direct repeats in the enhancer sequences of the viral LTR (50–75 bp in length) have been shown to vary in copy number and subsequently alter viral expression [27,28]. An SNV retroviral vector containing a 110-bp direct repeat was found to delete at a frequency of 41% in a single cycle assay [29], and it was hypothesized that the RT and the nascent strand dissociate from the template during reverse transcription, reassociating with a homologous section of template, resulting in deletion of the directly repeated sequence and any intervening sequences [21].
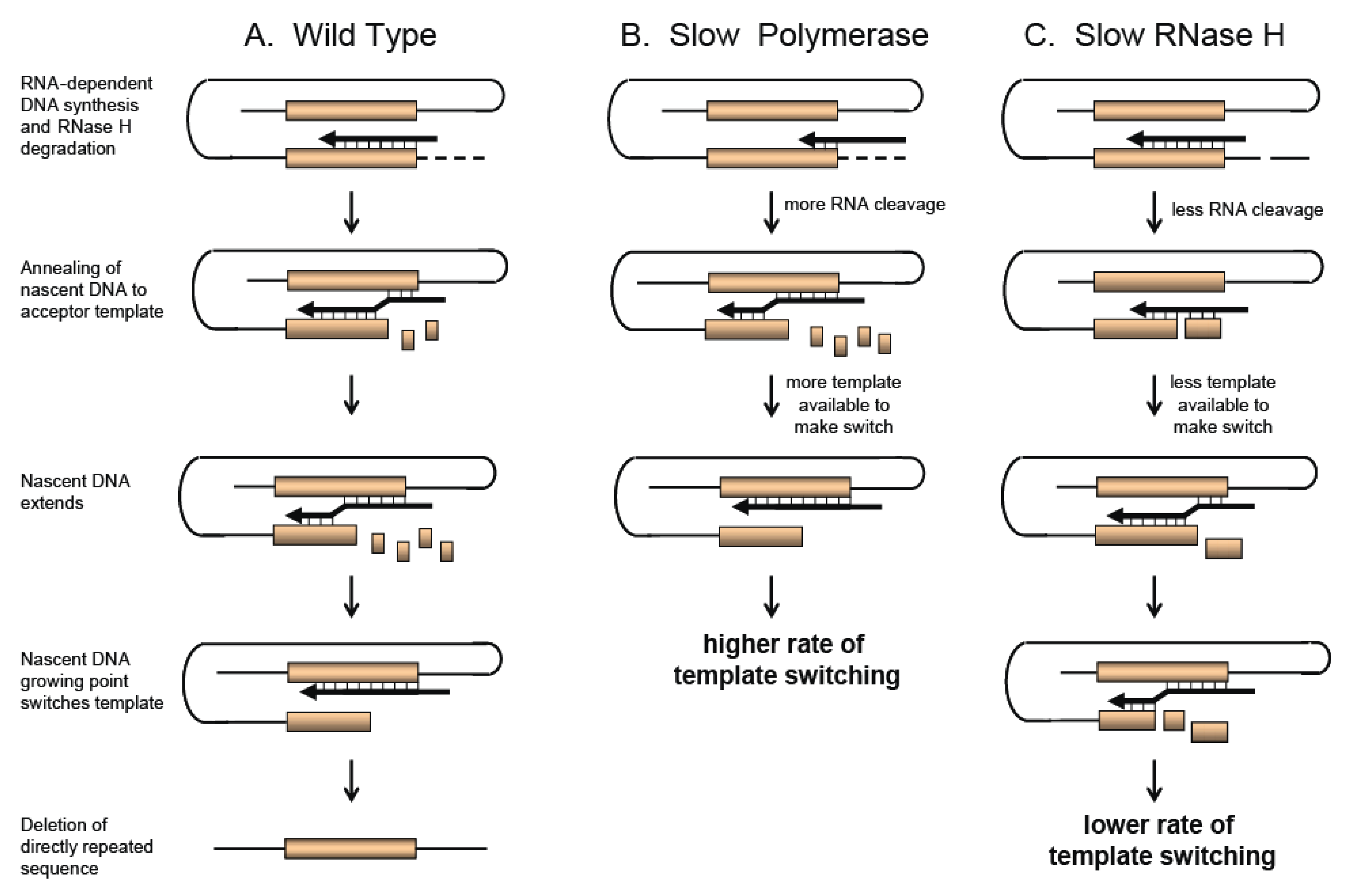
A model for direct repeat deletion and recombination [21,30,31] is shown in Figure 2A. The model shows that during RNA-dependent DNA synthesis, dissociation of RT and the growing nascent strand occurs due to RTs inherent low affinity to the template and the resulting minimal base pairing of the growing strand with the RNA template from RNase H degradation. RT and the growing strand can either reassociate with the upstream directly repeated sequence on the same RNA strand (intramolecular; shown in Figure 2) or the copackaged RNA strand (intermolecular), resulting in deletion of one of the direct repeats. Although shown with direct repeats, this model also describes recombination during DNA synthesis when the two homology blocks shown in Figure 2 are from two different RNAs.
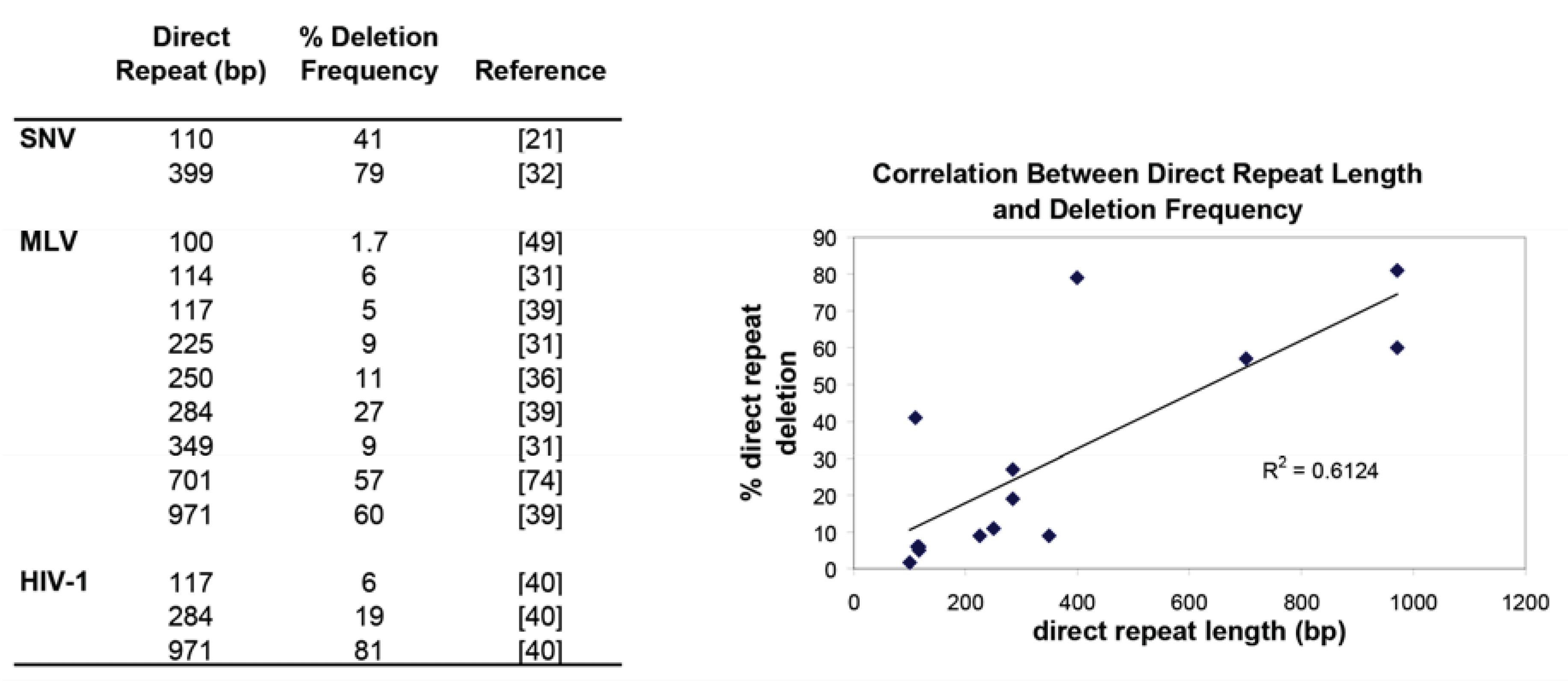
Direct repeat deletions occur mostly through intramolecular template switches [18] and the rates of template switching are similar during RNA- and DNA-dependent DNA synthesis as measured by direct repeat deletion of a 400-bp direct repeat in an SNV viral vector system [32]. The frequency of direct repeat deletions has been shown to be dependent upon the size of the direct repeats for both simple and complex retroviruses. Figure 3 summarizes the reported direct repeat deletion frequencies and shows in general that the larger the direct repeat, the higher the frequency of direct repeat deletion. Furthermore, the deletion frequencies are similar amongst the tested retroviruses suggesting that the MLV, SNV and HIV-1 RTs share some properties, such as template affinity and processivity, which determine the frequencies of RT template switching.
Overall, these results suggest that longer lengths of homology are one of the driving forces that influence a successful template switch. This was first suggested by Zhang and Temin that the efficiency of recombination is dependent upon the length of template identity [33]. Confirmation of this idea through an experiment using direct repeats was shown by keeping the direct repeat length constant, but varying the percent sequence similarity between the two direct repeats. Decreasing the percent sequence similarity to 95% dropped the deletion frequency ~2-fold, to 90% dropped the deletion frequency 4-fold, and anything lower than 82% homology dropped the deletion frequency to less than 1% [34]. Essentially, the experiment was also measuring non-homologous recombination and confirmed that nonhomologous recombination is inefficient, which was previously measured to be at least 1000-fold lower than the rate of homologous recombination [13,35].
Interestingly, increasing the distance between two direct repeats actually increases the frequency of deletion. Direct repeat deletion in an MLV vector in which a 701-bp direct repeat was separated by distances from 100 bp to 3.5 kb, resulted in high deletion frequencies (>90%) for distances >1500 bp [31]. This result provides insight into the organization of the RNA during reverse transcription indicating that large stretches of template are available for template switching upstream of the growing reverse transcription complex.
2.2. Frequency of Direct Repeat Deletion in Vivo Is Influenced by Nucleotide Pools and Mutations in RT
The frequency of direct repeat deletion is not only dependent upon the template itself, but can also be influenced by factors that affect RT activity. Drugs or mutations that affect the balance between polymerization and/or RNase H activity of RT can influence template switching. The dynamic copy choice model shown in Figure 2 demonstrates that RTs with a slow polymerase will have a higher rate of template switching, while RTs with lower RNase H activity will have lower rate of template switching [11,36,37]. By slowing down polymerization, more efficient degradation of the template strand occurs behind the growing nascent DNA strand providing more time for hydrogen bonding to occur between the acceptor template and the nascent DNA. This results in an increase in template switching (Figure 2B). In the case where RNase H activity is reduced, less nascent DNA is available to hydrogen bond with the template strand to facilitate a template switch, resulting in a reduced frequency of template switching (Figure 2C).
Support for this model has been validated by a number of studies. To induce RT to slow down polymerization, cells can either be treated with hyrdoxyurea (HU) to upset cellular nucleotide pools or treated with the nucleoside analog AZT to severely slow down polymerization. As expected, HU treatment in general showed a 2-fold increase in direct repeat deletion for both simple and complex retroviruses: SNV, 2.1-fold [38,39], MLV, 1.7–2.7-fold [11,36,39], and HIV-1, 1.8- 2.3-fold [37,40]. AZT treatment was also shown to increase direct repeat deletion in a dose dependent manner. The deletion frequency of a 250-bp direct repeat was shown to increase from 16% to 40% with increasing concentrations of AZT [41]. It was hypothesized that incorporation and excision of AZT-monophosphate slows down DNA synthesis, and this in turn increases template switching. This was also confirmed by analysis of a 117-bp direct repeat from an HIV-1 vector in which the presence of AZT caused a 1.6-fold increase in the deletion frequency [40]. RNA structure can also create pause sites and slow down polymerization resulting in an increase in template switching [42,43,44,45,46,47,48]. In MLV, a stem-loop structure placed between a 110-bp direct repeat increased deletion frequencies up to 5-fold [49]. Furthermore, studies using a 61-bp direct repeat separated by stem loops, showed deletion frequencies increased from 5% up to 47%, as the stability of the stem loop increased [50]. Lastly, even nucleotide sequence, for example, homopolymeric runs, can increase template switching [21,29,51,52].
Mutations in RT that also directly affect the polymerase activity and slow down polymerization, as expected, increase template switching. Mutations in the MLV RT dNTP binding site, polymerase active site, and thumb domain, which affect RT processivity, were also shown to increase direct repeat deletion in vivo [36]. This was also proven to be true for HIV-1, as mutations near the polymerase active site or dNTP binding site, such as K65R, Q151N, and E89G, increased direct repeat deletion 3-, 6- and 3-fold, respectively [37]. Interestingly, these mutations are selected in response to drug therapy in patients, and these results show that mutations conferring drug resistance can lead to increases in template switching, thus influencing not only recombination, but also viral evolution. Lastly, viral proteins, such as nucleocapsid (NC), can also influence template switching. In MLV, NC mutants which decreased the overall RT processivity (i.e., slowed down polymerization), also increased direct repeat deletion. Whereas, NC mutants which caused a decrease in template annealing, decreased direct repeat deletion, as expected [49].
The dynamic copy choice model also predicts that decreasing RNase H activity will lead to a decrease in template switching. Mutations in the MLV RNase H (S526A, Y598V, R657S) and HIV RNase H (H539N, D549N) known to affect RNase H activity were shown to decrease template switching, resulting in a 2-fold decrease in direct repeat deletion [36,37]. Experiments in which MLV virions were phenotypically mixed to have lower levels of RNase H, demonstrated lower levels of direct repeat deletion [53]. For HIV, mutational analysis showed that mutations in the RNase H primer grip that affected template binding, and subsequently decreased RNase H activity, also decreased direct repeat deletion [54]. Interestingly, RT mutations in the connection and RNase H domain acquired by HIV-1 treatment-experienced patients, which reduce RNase H cleavage, also show a decrease in direct repeat deletion [55,56]. Eight novel mutations in HIV-1, E312Q, G335C/D, N348I, A360I/V, V365I, and A376S were identified that contribute to AZT resistance and also decrease template switching [55,56].
2.3. In Vitro Studies Dissecting the Biochemical Nature of Template Switching
Template switching events have also been extensively studied in vitro using donor and acceptor oligonucleotide templates, and have mostly modeled recombination events occurring during minus-strand synthesis. The “Dock and Lock” in vitro model [57,58,59] for strand transfer complements the in vivo dynamic copy choice model. During RNA-dependent DNA synthesis, RNase H cleavage occurs behind the polymerizing RT exposing regions of single-stranded cDNA that are accessible to anneal (“dock”) to an acceptor RNA template. RT and the nascent cDNA can then switch templates (“lock”) and continue synthesizing on the acceptor RNA template. This model for invasion-driven transfer as expected is influenced by RT polymerization activity, RNase H activity, and RNA structure.
Detailed studies on the sites of strand invasion, show that the invasion sites onto the acceptor template normally occur at the base of a strong hairpin structure or a site where RT pausing was significant [42,58,59,60,61]. Pausing slows down polymerization and allows more RNase H cleavages to occur, enhancing the probability for the acceptor RNA and nascent DNA to interact and undergo a template switch [48,61,62,63]. Destefano et al. showed that strand transfer was 25% lower in the absence of a pause site, and decreasing dNTP concentrations in the reactions also lead to an increase in strand transfer [62]. Mutations in RT polymerase that affect dNTP binding, such as Q151N and V148I in HIV-1, also show enhanced strand transfer that is attributed to stalled DNA synthesis and thus slower polymerization [64]. RNA hairpin structures such as the dimerization initiation signal (DIS) have also been shown to promote strand transfer in vitro [65,66] and will be discussed in further detail in subsequent sections. Studies on DIS suggest that dimerization increases the local concentration of the nascent DNA and acceptor RNA template increasing the rate of strand transfer.
The “Dock and Lock” model is dependent upon RNase H cleavage and the removal of the short RNA oligos to provide access for strand invasion. In vitro assays have shown that MLV or HIV-1 RTs that are deficient in RNase H activity cannot undergo template switching [62,63] and fail to undergo minus strand transfer [67,68]. Strand invasion and subsequent strand transfer is thus dependent upon both polymerase-dependent and polymerase-independent RNase H cleavage. Pausing increases polymerase-dependent primary and secondary RNase H cleavages [69] helping to create an invasion site, while the excess of polymerase-independent RTs packaged per virion (50–100) degrade the remaining hybridized RNA fragments, which also helps to contribute to strand transfer [70,71,72,73]. Overall, the in vitro and in vivo models for strand transfer are consistent in showing that the two activities of RT, polymerase and RNase H, play a major role in influencing recombination events.
2.4. The Use of Directly Repeated Sequences for the Development of Viral Vectors for Gene Therapy and for Probing In Vivo Reverse Transcription
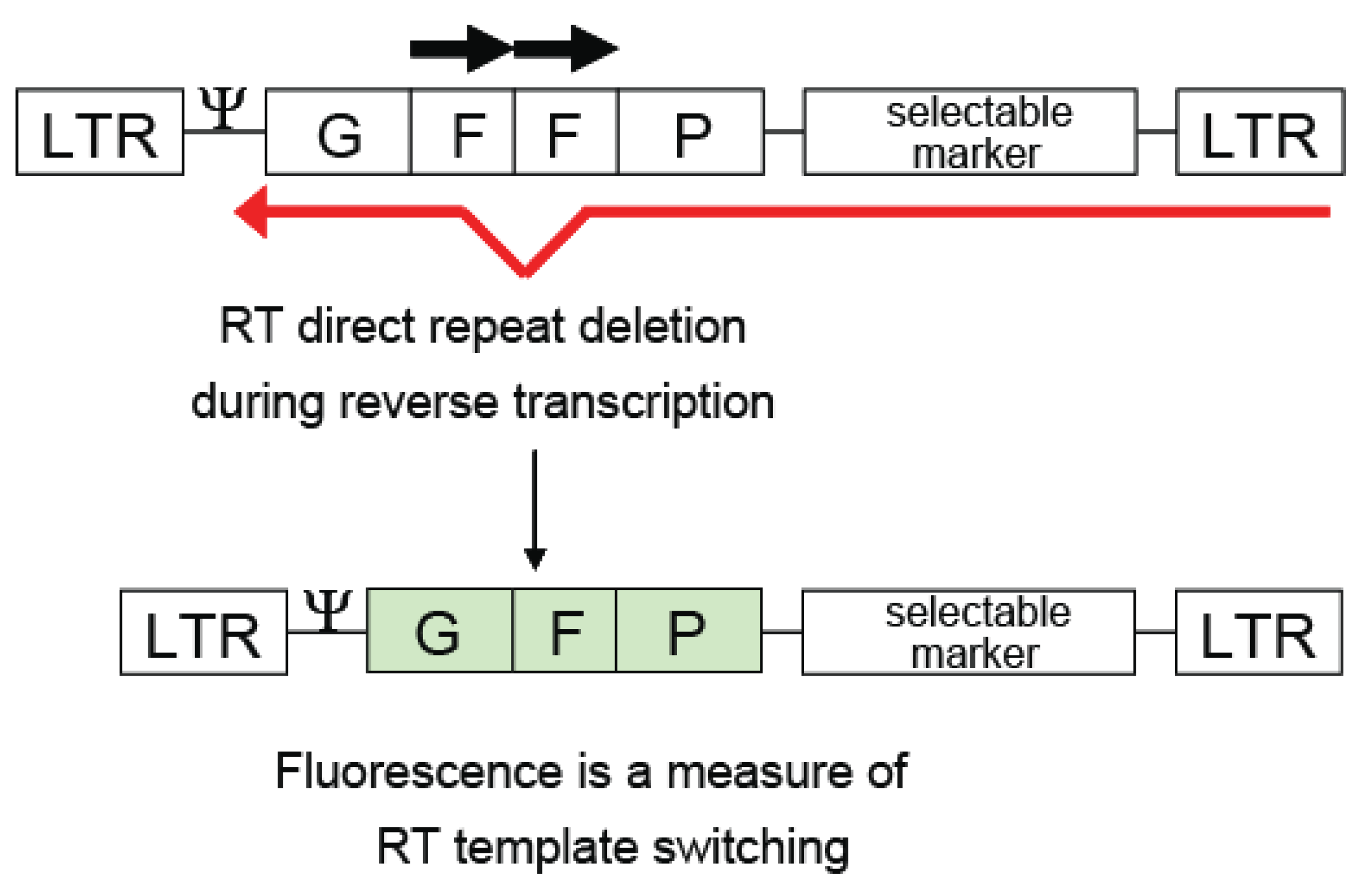
Direct repeat deletion is highly precise and accurately deletes repetitive sequences. Because of this phenomenon, direct repeats can be used to make informative vectors for laboratory use to study recombination by analyzing intra- and intermolecular template switching. Gene sequences from selectable drug markers such as neomycin, herpes simplex thymidine kinase gene, and lacZ can be duplicated to create direct repeats, and used to determine if viral infection has taken place. Even more informative is the use of fluorescent gene sequences, such as the green fluorescent protein (GFP), to detect infection of MLV [36] or HIV-1 [37] in cell lines (Figure 4).
The power of these vectors can be extended further to also delete unwanted sequences after infection. For example, placing a selectable marker between the direct repeats will result in a virus lacking the selectable marker after infection [74,75]. Likewise, placing the packaging signal between the direct repeats can result in >99% efficient deletion [30,74,75], which would be useful for gene therapy and delivery of genes to target cells without further mobilization of the viral vector.
Overall, template switching is a dynamic process. A recent paper demonstrates that RT is able to slide, flip orientations, and dissociate from the template during reverse transcription [76]. These properties allow RT to complete the necessary “jumps” required during minus-strand strong-stop and plus-strand strong-stop transfer to complete reverse transcription, and undergo strand transfer events to reassort mutations and generate recombinants to ensure the survival of progeny virus.
3. Recombination: Rates, RNA Copackaging, and Fidelity
3.1. Recombination Rates in a Single Replication Cycle
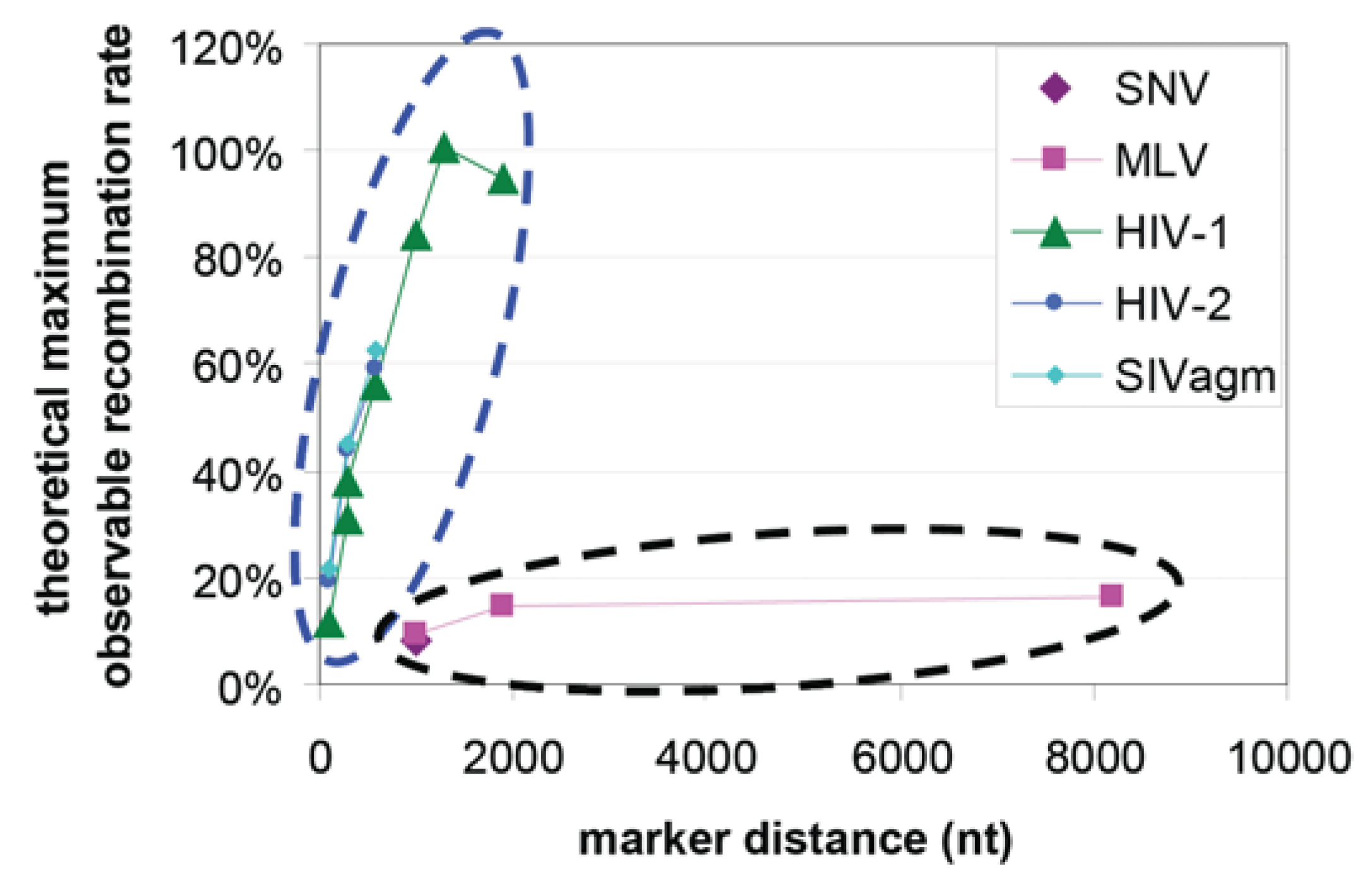
Retrovirus recombination rates were first measured in SNV and it was found that genetic markers separated by 1 kb recombined at 4% per replication cycle [13]. Similar measurements were made with MLV and it was found that markers separated by 1 kb, 1.9 kb, and 7.1 kb recombined at 4.7%, 7.4%, and 8.2%, respectively, per replication cycle [77]. Recombination rates were also measured in HIV-1 using the same template sequences as the MLV study and it was found that the recombination rates per replication for markers separated by 1 kb, 1.3 kb, and 1.9 kb were 42.4%, 50.4%, and 47.4%, respectively [78]. Using a different set of genetic markers, it was concluded that SIVagm and HIV-2 also recombine at high rates similar to HIV-1 [79,80]. Since the various assays used above generate rates that may not be directly comparable, we have converted the measured rates into percent of theoretical maximum measurable rates [79] as shown in Figure 5. These results point to the conclusions that lentiviruses (HIV-1, HIV-2, and SIVagm) appear to recombine more frequently than gammaretroviruses (SNV and MLV).
The cause of these differences in recombination frequencies is puzzling because RTs from SNV, MLV, and HIV-1 have similar template switching rates as mentioned above. This is further supported by the observation that multiple crossovers are often observed in SNV and MLV recombination, even though such events should be exceedingly rare based on their recombination rates [7,81]. Similarly, when examining MLV proviruses that are known to contain sequences from two different parents, frequent recombination can be detected elsewhere in the genomes [82]. These observations led to the conclusion that in SNV and MLV, recombination only occurs in a distinct retroviral population [18]; however, the nature of this population remains to be fully elucidated. Recent studies suggest that the efficiency of the RNA derived from two separate proviruses to copackage into the same particle has a significant impact on the observed recombination rates (discussed in detail below), providing insight into a possible mechanism that could contribute to the observed different recombination rates.
3.2. RNA Copackaging and Its Impact on Recombination Rates
The recombination rates discussed in the previous section (Figure 5) were measured using viruses derived from the same strain or molecular clones. However, recombination rates can be significantly lower between HIV-1 variants from different subtypes [83]. Subsequent analyses revealed that a 6-nt palindromic sequence in the 5' untranslated region termed dimerization initiation signal (DIS) [84,85,86] plays a major role in determining the recombination potential of two HIV-1 variants [83,87,88]. Two variants with different DIS palindromes recombine less frequently than two variants with the same DIS palindrome. HIV-1 from various subtypes can have different DIS sequences; for example, most subtype B HIV-1 variants have GCGCGC in their DIS, whereas most subtype C variants have GTGCAC in their DIS [89].
Viral RNA dimerizes, or selects its copackaged partner, prior to encapsidating into particles [88,90]. Experimental evidence suggests that the two viral RNAs initiate their interactions by forming base-pairing in the palindromic DIS sequences [91,92]. By labeling the RNA in the virion at the single RNA level, it was shown that RNAs derived from two separate HIV-1 genomes can dimerize very efficiently when they contain the same DIS palindrome [93]. Assuming equal expression and random assortment, the theoretical distribution of particles containing two RNAs from parent 1, one RNA from each parent virus, and two RNAs from parent 2 is 25%:50%:25% (Hardy-Weinberg Equilibrium). It was shown that when the two parent RNAs were expressed at similar levels and contained the same GCGCGC palindromes in their DIS, heterozygous particles were generated ~45% of the time, demonstrating that RNA from different viruses are packaged close to random distribution [93]. However, when the two parent viruses contained different DIS palindromes, for example, one had GCGCGC and the other GTGCAC, these RNAs were copackaged together far less frequently (~20%) [93]. Furthermore, recombination rates were significantly lower with HIV-1 containing different DIS palindromes [83]. In another experiment in which two HIV-1 RNAs were modified to contain nonpalindromic DIS sequences that still could interact with each other, for example, one virus has CCCCCC and the other GGGGGG, copackaging of the two RNAs increased beyond that predicted by random assortment (from ~45% to ~70%), and the recombination rate also increased beyond that of two wild-type viruses containing the same palindromes [88]. Together, these experimental results reveal that the ability of the two RNAs to be copackaged directly determines the recombination potential of two HIV-1 variants.
In the previous section we discussed that the recombination rates of gammaretroviruses appear to be lower than those of lentiviruses, although the RTs of these viruses appear to have similar abilities to perform template switching. Since the ability of the RNAs to be copackaged determines recombination potential, if gammretroviruses do not form heterozygous particles as efficiently as lentiviruses, this could explain the lower observed recombination rates. Indeed, experimental evidence suggests that RNAs derived from two gammaretroviruses may not be copackaged together as efficiently as those of two lentiviruses (HIV-1) [94]. It was found that recombination between two MLV vectors that were introduced into producer cells by cotransfection recombine more efficiently than the same two vectors introduced into cells at separate times by step-transfection [95]. As DNAs from cotransfection are more likely to be integrated close to one another than those from step-transfection, it was proposed that perhaps MLV RNA dimerizes very early during biogenesis and is more likely to pair with RNA transcribed from the same provirus, thereby promoting homodimerization and increasing the ratios of homozygous particles in the viral population. This attractive hypothesis has not been proven directly, as the proportion of the heterozygous virus population in MLV and factors that affect these proportions remain unknown.
The prerequisite for copackaging of RNA derived from two proviruses is having cells infected by more than one virus (double infection). Cell culture studies showed that double infection occurs more frequently than expected from the multiplicity of infection (moi) [96]; part of the cause for the increased double infection is that some target T cells express more receptors/coreceptors and are more susceptible to infection [97]. Double infection is more frequent via cell-mediated than the cell-free infection route (ref) as transfer of multiple viruses occurs during cell-mediated infection [96,98]. The frequency of double infected cells in HIV-1-infected patients have also been studied. In an earlier study using in situ hybridization, it was shown that the infected spleen cells of infected patients contain multiple copies of HIV-1 DNA [99]. A recent study used a newly developed single cell assay to study PBMCs of HIV-1 infected patients and found that most of the infected cells contained one copy of HIV-1 DNA [100]; as the infection MOI is unknown, it is unclear how frequently doubly-infected cells are expected. Thus it is difficult to compare this study with the cell culture system. However, the two studies of infected patient samples appear to point to fairly different conclusions. At this time, it is unclear whether double HIV-1 infected cells exist in spleen cells more frequently than in PBMCs or different infected individuals have different distributions of doubly-infected cells. Further studies will be needed to distinguish these possibilities.
3.3. Fidelity of Recombination and Its Contribution to the Overall Mutation Frequency
Retroviral homologous recombination events are generally precise because this process involves the base-pairing of the donor and acceptor template sequences as described in the dynamic copy-choice model. This is further demonstrated by the efficient reconstitution of marker genes during direct repeat deletion as discussed in the previous section. The accuracy of recombination events has also been directly examined for HIV-1. In an intersubtype HIV-1 recombination study [101], 152 recombination events were identified and their junction sequences analyzed. Although most of the recombination events were accurate (143/152), 6% of the junctions (9/152) contained errors. These erroneous events included substitutions (5/9), insertions caused by template misalignment (2/9), and deletions or substitutions likely caused by template misalignment (2/9). These 9 mutations were among the 76 mutations identified in the viral genomes in this study and occupied a significant portion of the total mutations generated. Therefore, recombination events are mostly accurate but do make occasional mistakes. Because recombination occurs very frequently during retroviral replication, these occasional mistakes can accumulate to a significant level [101].
4. Effects of Sequence Diversity and RNA Secondary Structures on Recombination
4.1. The Effects of Sequence Diversity on HIV-1 Recombination
HIV-1 has acquired extensive genetic diversity after its entry into the human population. Although variants from one subtype can dominate HIV-1 infection in certain countries, co-circulation of variants from multiple subtypes can be found in many geographic areas. Therefore, HIV-1 recombination can occur between variants with a wide range of sequence similarities. As depicted in the dynamic copy-choice model (Figure 2), stretches of homologous sequence are required between the newly-synthesized DNA and the acceptor RNA template in the annealing region for template switching to occur, and the larger the region of homology, the more likely recombination can occur (Figure 3). Using single-cycle replication assays, thereby eliminating the effects of natural selection, several studies demonstrated that increased sequence diversity produced reduced levels of recombination in different regions of the HIV-1 genome including gag [102], pol [103], and env [104,105,106]. A detailed study analyzing events in the pol gene in one round of replication showed that the recombination frequency between subtypes B and F (90% nucleotide sequence similarity) was ~30% lower than that between two subtype B variants (96% nucleotide sequence similarity) [103]. Analogous conclusions were obtained in a recent similar study on env recombination, although the different quantification methods make a direct comparison of the results difficult [106]. Recombination in the gag gene in a 0.3-kb target sequence in the capsid region was also studied by detecting Gag expression using flow cytometry. Compared with recombination between variants containing identical target sequences, it was found that recombination was reduced 3-fold, and 10–13-fold when the sequence similarity was reduced to 91%, and 72–73%, respectively [102].
Recombination frequencies between HIV-1 variants from the same subtype, different subtype, and different groups have been studied using a gfp gene as a marker to measure recombination events. Although the target gfp gene sequence is identical in all of these studies, the HIV-1 genomes contain different levels of sequence similarity. Compared with the events from two variants of the same subtype, gfp recombination is 2-fold less in two variants from different HIV-1 subtypes containing the same DIS sequences; these variants have ~85–90% sequence similarity [83,87]. Furthermore, the gfp recombination rate is drastically reduced (~5-fold) between HIV-1 variants from different groups that contain ~70% sequence similarity even though these variants have the same DIS sequences [102]. Therefore, sequence diversity can also indirectly reduce recombination in regions with high local homology. The exact mechanism underlying such an effect remains to be elucidated. Differences in the primary sequences might produce differences in secondary structures between distant strains that impair co-packaging or reduce RNA accessibility to RT during strand switching.
Several studies investigated the correlation between recombination hot-spots and sequence similarity in the HIV-1 genome. While one study could associate the location of hot-spots with areas of high sequence homology [107], subsequent studies indicated that although sequence diversity can produce local fluctuations in crossover frequency, it is unlikely to produce recombination hotspots [103,108,109,110]. The distribution of recombination breakpoints in vivo is more likely the result of the interplay between sequence homology, RNA structure, and RT strand switching that produce complex and different recombination patterns for each pair of HIV-1 strains.
Taken together, these results show that recombinants between highly similar strains, such as those belonging to the same subtype, are generated at the highest frequencies. More diverse viruses, such as variants belonging to different subtypes, recombine with lower frequencies. Finally, very distant HIV-1 strains, such as those classified as different groups, recombine at very low frequencies [102,103].
4.2. The Effects of RNA Secondary Structure on HIV-1 Recombination
RNA secondary structures have been found to be capable of producing recombination hotspots. Different groups have identified recombination hotspots associated with RNA hairpins in gag [111], pol [110] and env [16,112]. The mechanism regulating the effect of RNA structures on recombination is still not completely understood. The dynamic copy model predicts that stalling of the RT at the base of stem-loops, which alters the balance between DNA synthesis and RNA degradation activities of the enzyme, should increase the rate of template-switching and consequently the frequency of recombination. However, secondary structures can induce template switching events even when correlation with pause sites during reverse transcription are not observed [65]. In such instances, characterized by recombination occurring inside the stem-loop rather than at its base, a direct interaction of the hairpin structures in the donor and acceptor templates has been proposed [113]. One model proposed that the single-stranded loops of nascent DNA and acceptor RNA template could anneal and initiate the pairing of the stems, eventually producing a cross-over in the descending stem of the acceptor RNA [112,114].
Recently, the entire structure of full-length HIV-1 RNA was predicted using the selective 2'-hydroxyl acylation analyzed by primer extension (SHAPE) methodology [115]. Comparison of the RNA and genetic structure demonstrate that many RNA structures are located between domains or genes encoded in the HIV-1 genome; this observation suggests that these RNA structures may have a role in modulating the translation or folding of HIV-1 proteins. In light of the SHAPE results, it appears that most of the previously identified recombination hotspots in the env gene, localize to inter-domain or inter-genic junctions, in both intra- and inter-subtype recombinants [15,104,107,110,111,112]. Given the association between RNA stem-loops and recombination described previously, these findings raise the intriguing possibility that the identified structures have two different functions in two relevant steps of viral replication: facilitating recombination and regulating translation. A recent study showed a direct association between crossover location and local inter-domain RNA structures in the env gene of HIV-1 [116] when recombination was limited to one cycle of replication. Furthermore, recombination breakpoints clustered around gene junctions due to the local secondary structure even in the absence of selection. The data obtained for the env gene strongly supports a role for RNA structures in recombination. Although the effect of the RNA structures on the recombination in the env gene is convincing, whether it can be applied to the entire HIV-1 genome is yet to be determined. For example, using a single cycle assay, the junctions of intra- and intersubtype recombination in the pol gene have been studied; when compared with the predicted structures from the SHAPE study, there is no association between RNA structures and breakpoints distribution [103]. However, stem-loops predicted in env are generally more prominent than those in pol [115]. It is possible that the different results obtained in the two regions in the HIV-1 genome are simply caused by the extent of the secondary structures, and only RNA structures with sufficient size or stability can affect recombination, whereas minor structures have no effect or only subtle effects that were not detected in these studies.
5. Fitness of the Progeny Recombinant
Recombination assorts polymorphisms in retroviral genomes. One of the consequences of this process is that the linkage of the co-evolved sequences may be disrupted in many recombinants. Therefore, although recombination increases diversity of the viral population, many of the newly generated recombinants may have altered or reduced replication fitness than their parents, which may predict that they will have a different probability to survive. Using different experimental approaches, two recent papers addressed the fitness of the newly generated recombinants. In one study, the recombination junctions in the env gene were determined and the functions of a set of recombinant Envs were tested using a single cycle assay [106]. It was shown that viruses with these recombinant Env proteins had varied abilities to replicate; although some recombinant Env proteins could generate viral titers similar to those of the parents, some recombinant Env proteins clearly generated far lower titers than those of the parental Env proteins. Comparing the functions of these Env and recombination junctions of the circulating strains suggests that the function of the Env proteins is an important factor for the survival of the recombinants [106].
In another study, the patterns of the observed recombinants in the pol gene were compared with or without the selection pressure on the Pol proteins [103] by analyzing recombinants generated in a single round assay and in a multiple round assay. Regardless of the requirement for Pol function, the recombination patterns between two subtype B variants were very similar; recombination junctions were observed throughout the pol gene. These results indicate that recombination between strains with high sequence similarity are likely to result in genomes that are functional; therefore, the distribution of crossovers is unaffected in the recombinant population regardless whether Pol function is selected. However, a very different picture emerged in the studies of recombination between subtypes B and F. When the Pol function was not required, recombination junctions were observed throughout the pol gene; in contrast, when the Pol function was selected, recombination junctions were observed in only a few clustered regions in the pol gene. These results revealed that although recombination between divergent strains such as subtypes B and F can occur throughout the pol gene, most of the recombinants did not survive multiple rounds of replication in which the Pol function was required.
Taken together, both of these studies revealed that viral replication fitness is an important factor for the survival of the recombinants; many newly generated recombinants, especially those from divergent variants, have altered replication fitness and face a strong purifying selection. These studies also suggest that despite the frequent observation of the circulating recombinant strains (see the following section for detail), there are many more recombinants that are generated but are lost as a result of the purifying selection.
6. Recombination in the HIV-1 Epidemics
6.1. Intersubtype HIV-1 Recombinants: Evidence of Recombination-Generated Variation in Current HIV- Isolates
One of the best examples of how recombination can generate diversity in a virus population is demonstrated by the observation of HIV-1 intersubtype recombinants. Although recombination occurs frequently with variants in the same subtype, these recombination events are often difficult to identify unless the parental viral sequences are known. In contrast, the genetic distance between different group M subtypes makes it easier to identify intersubtype recombinants. The increased co-circulation of different subtypes of HIV-1 in many areas of the world has fostered the generation of a high number of recombinant forms [3,117,118,119]. Several such circulating recombinant forms (CRFs) have become important in the global epidemic [2,120]; to date, 49 CRFs have been classified [121] in the HIV-1 pandemic. In addition, a large number of unique recombinant forms (URFs) have been detected in the infected population. Some of the early identified CRFs have become predominant in their area of origin, probably due to either their early introduction or to fitness advantage [119]. For example, CRF01_AE, a recombinant between subtype A1 and the putative subtype E, is the predominant strain in South-East Asia [122]; CRF01_AG, a variant which has been recently proposed to be in fact the parental subtype G rather than a recombinant [123], is the dominant form in West and Central Africa [120]; and CRFs 07 and 08, which are B/C recombinants that originally appeared among drug-users in China, have now spread to a vast portion of the infected Chinese population [124].
The genetic composition of recombinants usually reflects the level of co-circulation of different strains in a given geographical area. B/F recombinants such as CRFs 12, 17, 28, 29, 38, 39, 40, 44, and 46, are widely spread in South America, where they represent a high percentage of infections. B/G recombinants such as CRFs 14, 20, 23, and 24, have been identified mostly in Cuba, Spain and Portugal [125,126,127], while recombinants involving the CRF01 (CRFs 15, 33, 34, and 48) are mostly common in South-East Asia. On the other hand, most of the CRFs classified so far have been isolated from Central-African patients, due to the extensive co-circulation of different genetic forms in their area of origin [119]. Overall, the most recent estimates place the prevalence of recombinant forms at about 20% of infections worldwide; these surveys also indicate that non-B subtypes and recombinants have been stably introduced into areas previously restricted to infection by the subtype B variants such as North America and Western Europe [3,128]. The possible impact of recombination on the evolution of escape mutants and drug resistant strains has raised concern in the medical community about long term outcome of antiviral treatment and the effects on patient management.
6.2. Detection, Replicative Fitness, and Drug-Resistance of the Intersubtype Recombinants
Most antivirals and detection assays commercially available for HIV-1 have been developed and tested using subtype B strains. The performance of most HIV-1 RNA quantification assays in identifying subtypes different from B are variable between different assays and usually lower compared to subtype B detection [129,130]. As a consequence, depending on the subtypes involved in each recombinant and the patterns of recombination, the levels of circulating virus could be undetectable or grossly underestimated by the currently available kits [131]. Plasma level of viral RNA is an important parameter for determining drug treatment and assessing the efficacy of therapy [132,133]; unreliable RNA determination could have detrimental effects on patient prognosis and treatment outcome.
Although data are available for a limited number of subtypes, HIV-1 variants demonstrate a range of different replicative fitnesses [134]. Thus, it is possible that intersubtype recombinants may acquire a higher fitness than a parental strain; indeed, CRF02_AG was hypothesized to have had such a gain of fitness, thereby allowing this recombinant form to be a predominant variant in West and Central Africa [135,136]. Similarly, subtype B variants were introduced to South-East Asia before CRF01_AE entered this region through the intravenous drug user population [137], but this recombinant quickly became the predominant form in all risk categories [122] possibly due to higher infectivity of the recombinant form [138]. Despite the differences in replicative fitness, direct analyses of diverse HIV-1 strains only detected significant differences in the viral pathogenicity of subtype D infection [139,140,141], indicating that further studies are needed to clarify the link between replicative fitness and pathogenesis in intersubtype recombinants.
Most of our current understanding of mutations associated with drug resistance and their mechanisms are from studies based on subtype B variants. However, it is known that several non-B subtype variants carry polymorphisms associated with low level of drug resistance in subtype B variants [142,143]. Such sequence polymorphisms can produce different responses to specific drugs or classes of drugs in some HIV-1 variants and are associated with various drug-resistance evolutionary pathways [144]. Furthermore, recent studies showed that although most HIV-1 strains remain susceptible to antiretroviral drugs [145,146,147], variants from different HIV-1 subtypes can have intrinsic differences in the level of resistance to antiretrovirals [148]. Recombination offers the potential to bring together existing resistance mutations, or beneficial molecular features of intrinsically-resistant enzymes [149], thereby generating progenies with more selective advantages than their parents. Indeed, mathematical modeling studies demonstrated that by reducing the impact of linkage disequilibrium, recombination allows a faster reassortment of mutations and a higher adaptation rate [150].
6.3. Recombinants and Variation Create Challenges to HIV-1 Vaccine Development
Vaccination is the most effective way of preventing viral diseases. However, the extensive genetic variability and the diverse assortment of antigen epitopes associated with HIV-1 strains create a major challenge for the development of a successful AIDS vaccine. During early infection, HIV-1 evolution is driven by selection and fixation of mutations conferring escape to cytotoxic T-cell responses. HIV-1 variants carrying escape mutations can be selected for in the infected individual, or can be acquired directly through the chain of transmission [151]. Since the initial transmission in an individual most often involves between 1–3 infection events [152], recombination between founder viruses can facilitate escape from cytotoxic T-cell responses by allowing re-assortment of mutations escaping immune response during early HIV-1 infection. The effectiveness of recombination in epitope shuffling has been examined in recent studies of patients infected with more than one strain of HIV-1. It was shown that by the time the first anti-HIV antibody was detected, most of the circulating viruses were already recombinants [153]. Additionally, a recent case study showed that rapid recombination occurred after superinfection between the existing virus and the newly transmitted virus to generate recombinants containing different epitopes in Gag and Env to avoid the pre-existing cytotoxic T cell response. The generation of escape variants caused the failure of the immune response to control HIV-1 replication and viral loads escalated in the patient [154]. Furthermore, in non-human primate models, recombination between the live attenuated SIV vaccine and pathogenic strains of SIV has resulted in breakthrough viremia and the generation of more virulent strains of SIV [155,156,157,158]. These studies highlight the complications that can be caused by recombination events in the development of a successful HIV-1 vaccine.
For a vaccine to be effective on a population level it must elicit sterilizing immune responses that recognize a majority of the circulating strains of HIV-1. Efforts to overcome the problem of HIV-1 diversity include strategies designed to achieve high epitope coverage. One approach uses polyvalent mosaic immunogens derived by in silico recombination of natural strains of HIV. These so-called mosaic vaccines are intended to induce cellular immune responses that recognize genetically diverse virus isolates [159,160]. Another strategy is to design epitopes using the center of the tree (COT) approach [159,160]. Briefly, a COT sequence is calculated from a phylogenetic tree of different HIV-1 isolates, by re-rooting the tree at the point that describes the least-squares distance to all the tips on the tree. In this way a majority of the variety of the sequences in the tree can be included in a single epitope [159,160]. Two recent studies have demonstrated that such mosaic HIV-1 vaccines appear to increase the breadth and depth of cellular immune responses in Rhesus monkeys [161,162]. Whether these strategies will also be effective in preventing HIV-1 infection in humans remains to be demonstrated.
The impact of recombination on HIV-1 detection, pathogenicity, treatment, and vaccine design can have important consequences on patient management and therapy outcome. Most studies investigating these issues are focused on “pure” non-B subtypes, underlining the need for data regarding recombinant forms. Given the increasing global prevalence of recombinants, a better understanding of their biology in vivo would help in devising optimal treatment strategies and development of a vaccine.
7. Conclusions
Retroviruses have one of the highest recombination rates among all viruses. Frequent recombination reassorts viral sequences to generate variants containing different combinations of polymorphic sequences, thereby generating high diversity in the viral population, which improve the odds that some variants in the population can survive the ever changing selection pressure in the environment. There are clear examples of such advantages in the replication of HIV-1. For example, recombination can generate a variant to escape the host cell immune response; similarly, recombination can also assort existing drug-resistance mutations to generate a more resistant virus or a variant that is resistant to more than one drug. Additionally, by assorting sequences, recombination can also preserve diversity outside the selected region. Recombination is a major driving force for viral diversity; understanding the mechanisms and the factors that affect recombination can ultimately help us to develop better treatment and a preventative vaccine to control the AIDS epidemic.
Acknowledgements
This work was supported in part by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
References and Notes
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Anderson, J.P.; Bradac, J.A.; Carr, J.K.; Foley, B.; Funkhouser, R.K.; Gao, F.; Hahn, B.H.; Kalish, M.L.; Kuiken, C.; et al. HIV-1 nomenclature proposal. Science 2000, 288, 55–56. [Google Scholar] [CrossRef]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global trends in molecular epidemiology of HIV-1 during 2000–2007. AIDS 2011, 25, 679–689. [Google Scholar] [CrossRef]
- Coffin, J.M.; Hughes, S.H.; Varmus, H. Retroviruses; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1997; pp. 121–160. [Google Scholar]
- Yu, S.F.; Baldwin, D.N.; Gwynn, S.R.; Yendapalli, S.; Linial, M.L. Human foamy virus replication: A pathway distinct from that of retroviruses and hepadnaviruses. Science 1996, 271, 1579–1582. [Google Scholar] [CrossRef]
- Hulme, A.E.; Perez, O.; Hope, T.J. Complementary assays reveal a relationship between HIV-1 uncoating and reverse transcription. Proc. Natl. Acad. Sci. U. S. A. 2011, 1014522108. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Retroviral recombination and reverse transcription. Science 1990, 250, 1227–1233. [Google Scholar] [CrossRef]
- Yu, H.; Jetzt, A.E.; Ron, Y.; Preston, B.D.; Dougherty, J.P. The nature of human immunodeficiency virus type 1 strand transfers. J. Biol. Chem. 1998, 273, 28384–28391. [Google Scholar] [CrossRef]
- van Wamel, J.L.; Berkhout, B. The first strand transfer during HIV-1 reverse transcription can occur either intramolecularly or intermolecularly. Virology 1998, 244, 245–251. [Google Scholar] [CrossRef]
- Coffin, J.M. Structure, replication, and recombination of retrovirus genomes: Some unifying hypotheses. J. Gen. Virol. 1979, 42, 1–26. [Google Scholar] [CrossRef]
- Hwang, C.K.; Svarovskaia, E.S.; Pathak, V.K. Dynamic copy choice: Steady state between murine leukemia virus polymerase and polymerase-dependent RNase H activity determines frequency of in vivo template switching. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 12209–12214. [Google Scholar] [CrossRef]
- Junghans, R.P.; Boone, L.R.; Skalka, A.M. Products of reverse transcription in avian retrovirus analyzed by electron microscopy. J. Virol. 1982, 43, 544–554. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Genetic consequences of packaging two RNA genomes in one retroviral particle: Pseudodiploidy and high rate of genetic recombination. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 1556–1560. [Google Scholar] [CrossRef]
- Pathak, V.K.; Hu, W.S. "Might as Well Jump!" Template switching by retroviral reverse transcriptase, defective genome formation, and recombination. Semin. Virol. 1997, 8, 141–150. [Google Scholar] [CrossRef]
- Zhuang, J.; Jetzt, A.E.; Sun, G.; Yu, H.; Klarmann, G.; Ron, Y.; Preston, B.D.; Dougherty, J.P. Human immunodeficiency virus type 1 recombination: Rate, fidelity, and putative hot spots. J. Virol. 2002, 76, 11273–11282. [Google Scholar] [CrossRef]
- Galetto, R.; Moumen, A.; Giacomoni, V.; Veron, M.; Charneau, P.; Negroni, M. The structure of HIV-1 genomic RNA in the gp120 gene determines a recombination hot spot in vivo. J. Biol. Chem. 2004, 279, 36625–36632. [Google Scholar] [CrossRef]
- Jetzt, A.E.; Yu, H.; Klarmann, G.J.; Ron, Y.; Preston, B.D.; Dougherty, J.P. High rate of recombination throughout the human immunodeficiency virus type 1 genome. J. Virol. 2000, 74, 1234–1240. [Google Scholar] [CrossRef]
- Hu, W.S.; Bowman, E.H.; Delviks, K.A.; Pathak, V.K. Homologous recombination occurs in a distinct retroviral subpopulation and exhibits high negative interference. J. Virol. 1997, 71, 6028–6036. [Google Scholar] [CrossRef]
- Katz, R.A.; Skalka, A.M. Generation of diversity in retroviruses. Annu. Rev. Genet. 1990, 24, 409–445. [Google Scholar] [CrossRef]
- Temin, H.M. Sex and recombination in retroviruses. Trends Genet. 1991, 7, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.K.; Temin, H.M. Broad spectrum of in vivo forward mutations, hypermutations, and mutational hotspots in a retroviral shuttle vector after a single replication cycle: Deletions and deletions with insertions. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 6024–6028. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, S.; Varela-Echavarria, A.; Ron, Y.; Preston, B.D.; Dougherty, J.P. Genetic rearrangements occurring during a single cycle of murine leukemia virus vector replication: Characterization and implications. J. Virol. 1995, 69, 7991–8000. [Google Scholar] [CrossRef] [PubMed]
- Czernilofsky, A.P.; Levinson, A.D.; Varmus, H.E.; Bishop, J.M.; Tischer, E.; Goodman, H.M. Nucleotide sequence of an avian sarcoma virus oncogene (src) and proposed amino acid sequence for gene product. Nature 1980, 287, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.; Kosik, E. Mutagenesis of the region between env and src of the SR-A strain of Rous sarcoma virus for the purpose of constructing helper-independent vectors. Virology 1984, 136, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Omer, C.A.; Pogue-Geile, K.; Guntaka, R.; Staskus, K.A.; Faras, A.J. Involvement of directly repeated sequences in the generation of deletions of the avian sarcoma virus src gene. J. Virol. 1983, 47, 380–382. [Google Scholar] [CrossRef]
- Rhode, B.W.; Emerman, M.; Temin, H.M. Instability of large direct repeats in retrovirus vectors. J. Virol. 1987, 61, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Lovmand, S.; Kjeldgaard, N.O.; Jorgensen, P.; Pedersen, F.S. Enhancer functions in U3 of Akv virus: A role for cooperativity of a tandem repeat unit and its flanking DNA sequences. J. Virol. 1990, 64, 3185–3191. [Google Scholar] [CrossRef]
- Pedersen, K.; Lovmand, S.; Jorgensen, E.C.; Pedersen, F.S.; Jorgensen, P. Efficient replication and expression of murine leukemia virus with major deletions in the enhancer region of U3. Virology 1992, 187, 821–824. [Google Scholar] [CrossRef]
- Pathak, V.K.; Temin, H.M. Broad spectrum of in vivo forward mutations, hypermutations, and mutational hotspots in a retroviral shuttle vector after a single replication cycle: Substitutions, frameshifts, and hypermutations. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 6019–6023. [Google Scholar] [CrossRef]
- Julias, J.G.; Hash, D.; Pathak, V.K. E- vectors: Development of novel self-inactivating and self-activating retroviral vectors for safer gene therapy. J. Virol. 1995, 69, 6839–6846. [Google Scholar] [CrossRef]
- Delviks, K.A.; Pathak, V.K. Effect of distance between homologous sequences and 3' homology on the frequency of retroviral reverse transcriptase template switching. J. Virol. 1999, 73, 7923–7932. [Google Scholar] [CrossRef]
- Bowman, R.R.; Hu, W.S.; Pathak, V.K. Relative rates of retroviral reverse transcriptase template switching during RNA- and DNA-dependent DNA synthesis. J. Virol. 1998, 72, 5198–5206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Temin, H.M. Retrovirus recombination depends on the length of sequence identity and is not error prone. J. Virol. 1994, 68, 2409–2414. [Google Scholar] [CrossRef]
- An, W.; Telesnitsky, A. Effects of varying sequence similarity on the frequency of repeat deletion during reverse transcription of a human immunodeficiency virus type 1 vector. J. Virol. 2002, 76, 7897–7902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Temin, H.M. Rate and mechanism of nonhomologous recombination during a single cycle of retroviral replication. Science 1993, 259, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Delviks, K.A.; Hwang, C.K.; Pathak, V.K. Structural determinants of murine leukemia virus reverse transcriptase that affect the frequency of template switching. J. Virol. 2000, 74, 7171–7178. [Google Scholar] [CrossRef]
- Nikolenko, G.N.; Svarovskaia, E.S.; Delviks, K.A.; Pathak, V.K. Antiretroviral drug resistance mutations in human immunodeficiency virus type 1 reverse transcriptase increase template-switching frequency. J. Virol. 2004, 78, 8761–8770. [Google Scholar] [CrossRef]
- Julias, J.G.; Pathak, V.K. Deoxyribonucleoside triphosphate pool imbalances in vivo are associated with an increased retroviral mutation rate. J. Virol. 1998, 72, 7941–7949. [Google Scholar] [CrossRef]
- Pfeiffer, J.K.; Topping, R.S.; Shin, N.H.; Telesnitsky, A. Altering the intracellular environment increases the frequency of tandem repeat deletion during Moloney murine leukemia virus reverse transcription. J. Virol. 1999, 73, 8441–8447. [Google Scholar] [CrossRef]
- An, W.; Telesnitsky, A. Frequency of direct repeat deletion in a human immunodeficiency virus type 1 vector during reverse transcription in human cells. Virology 2001, 286, 475–482. [Google Scholar] [CrossRef]
- Nikolenko, G.N.; Palmer, S.; Maldarelli, F.; Mellors, J.W.; Coffin, J.M.; Pathak, V.K. Mechanism for nucleoside analog-mediated abrogation of HIV-1 replication: Balance between RNase H activity and nucleotide excision. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 2093–2098. [Google Scholar] [CrossRef]
- Gao, L.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Insights into the multiple roles of pausing in HIV-1 reverse transcriptase-promoted strand transfers. J. Biol. Chem. 2007, 282, 6222–6231. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.J.; Buiser, R.G.; Mallaber, L.M.; Fay, P.J.; Bambara, R.A. Parameters that influence processive synthesis and site-specific termination by human immunodeficiency virus reverse transcriptase on RNA and DNA templates. Biochim. Biophys. Acta 1992, 1131, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Harrison, G.P.; Mayo, M.S.; Hunter, E.; Lever, A.M. Pausing of reverse transcriptase on retroviral RNA templates is influenced by secondary structures both 5' and 3' of the catalytic site. Nucl. Acids Res. 1998, 26, 3433–3442. [Google Scholar] [CrossRef] [PubMed]
- Klasens, B.I.; Huthoff, H.T.; Das, A.T.; Jeeninga, R.E.; Berkhout, B. The effect of template RNA structure on elongation by HIV-1 reverse transcriptase. Biochim. Biophys. Acta 1999, 1444, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Lapadat-Tapolsky, M.; De Rocquigny, H.; Van Gent, D.; Roques, B.; Plasterk, R.; Darlix, J.L. Interactions between HIV-1 nucleocapsid protein and viral DNA may have important functions in the viral life cycle. Nucl. Acids Res. 1993, 21, 831–839. [Google Scholar] [CrossRef]
- Suo, Z.; Johnson, K.A. Effect of RNA secondary structure on the kinetics of DNA synthesis catalyzed by HIV-1 reverse transcriptase. Biochemistry 1997, 36, 12459–12467. [Google Scholar] [CrossRef]
- Wu, W.; Blumberg, B.M.; Fay, P.J.; Bambara, R.A. Strand transfer mediated by human immunodeficiency virus reverse transcriptase in vitro is promoted by pausing and results in misincorporation. J. Biol. Chem. 1995, 270, 325–332. [Google Scholar] [CrossRef]
- Zhang, W.H.; Hwang, C.K.; Hu, W.S.; Gorelick, R.J.; Pathak, V.K. Zinc finger domain of murine leukemia virus nucleocapsid protein enhances the rate of viral DNA synthesis in vivo. J. Virol. 2002, 76, 7473–7484. [Google Scholar] [CrossRef]
- Lanciault, C.; Champoux, J.J. Pausing during reverse transcription increases the rate of retroviral recombination. J. Virol. 2006, 80, 2483–2494. [Google Scholar] [CrossRef]
- Klarmann, G.J.; Schauber, C.A.; Preston, B.D. Template-directed pausing of DNA synthesis by HIV-1 reverse transcriptase during polymerization of HIV-1 sequences in vitro. J. Biol. Chem. 1993, 268, 9793–9802. [Google Scholar] [CrossRef]
- Shen, W.; Gao, L.; Balakrishnan, M.; Bambara, R.A. A recombination hot spot in HIV-1 contains guanosine runs that can form a G-quartet structure and promote strand transfer in vitro. J. Biol. Chem. 2009, 284, 33883–33893. [Google Scholar] [CrossRef] [PubMed]
- Brincat, J.L.; Pfeiffer, J.K.; Telesnitsky, A. RNase H activity is required for high-frequency repeat deletion during Moloney murine leukemia virus replication. J. Virol. 2002, 76, 88–95. [Google Scholar] [CrossRef]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Barr, R.; Pathak, V.K. Mutations in human immunodeficiency virus type 1 RNase H primer grip enhance 3'-azido-3'-deoxythymidine resistance. J. Virol. 2007, 81, 6837–6845. [Google Scholar] [CrossRef] [PubMed]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Boyer, P.L.; Hughes, S.H.; Coffin, J.M.; Jere, A.; Pathak, V.K. HIV-1 reverse transcriptase connection subdomain mutations reduce template RNA degradation and enhance AZT excision. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 10943–10948. [Google Scholar] [CrossRef] [PubMed]
- Nikolenko, G.N.; Delviks-Frankenberry, K.A.; Palmer, S.; Maldarelli, F.; Fivash, M.J., Jr.; Coffin, J.M.; Pathak, V.K. Mutations in the connection domain of HIV-1 reverse transcriptase increase 3'-azido-3'-deoxythymidine resistance. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Negroni, M.; Buc, H. Copy-choice recombination by reverse transcriptases: Reshuffling of genetic markers mediated by RNA chaperones. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 6385–6390. [Google Scholar] [CrossRef]
- Roda, R.H.; Balakrishnan, M.; Hanson, M.N.; Wohrl, B.M.; Le Grice, S.F.; Roques, B.P.; Gorelick, R.J.; Bambara, R.A. Role of the reverse transcriptase, nucleocapsid protein, and template structure in the two-step transfer mechanism in retroviral recombination. J. Biol. Chem. 2003, 278, 31536–31546. [Google Scholar] [CrossRef]
- Roda, R.H.; Balakrishnan, M.; Kim, J.K.; Roques, B.P.; Fay, P.J.; Bambara, R.A. Strand transfer occurs in retroviruses by a pause-initiated two-step mechanism. J. Biol. Chem. 2002, 277, 46900–46911. [Google Scholar] [CrossRef]
- Chen, Y.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Steps of the acceptor invasion mechanism for HIV-1 minus strand strong stop transfer. J. Biol. Chem. 2003, 278, 38368–38375. [Google Scholar] [CrossRef]
- Hanson, M.N.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Effects of donor and acceptor RNA structures on the mechanism of strand transfer by HIV-1 reverse transcriptase. J. Mol. Biol. 2005, 353, 772–787. [Google Scholar] [CrossRef]
- DeStefano, J.J.; Mallaber, L.M.; Rodriguez-Rodriguez, L.; Fay, P.J.; Bambara, R.A. Requirements for strand transfer between internal regions of heteropolymer templates by human immunodeficiency virus reverse transcriptase. J. Virol. 1992, 66, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.J.; Bambara, R.A.; Fay, P.J. The mechanism of human immunodeficiency virus reverse transcriptase-catalyzed strand transfer from internal regions of heteropolymeric RNA templates. J. Biol. Chem. 1994, 269, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Operario, D.J.; Balakrishnan, M.; Bambara, R.A.; Kim, B. Reduced dNTP interaction of human immunodeficiency virus type 1 reverse transcriptase promotes strand transfer. J. Biol. Chem. 2006, 281, 32113–32121. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Fay, P.J.; Bambara, R.A. The kissing hairpin sequence promotes recombination within the HIV-I 5' leader region. J. Biol. Chem. 2001, 276, 36482–36492. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Roques, B.P.; Fay, P.J.; Bambara, R.A. Template dimerization promotes an acceptor invasion-induced transfer mechanism during human immunodeficiency virus type 1 minus-strand synthesis. J. Virol. 2003, 77, 4710–4721. [Google Scholar] [CrossRef]
- Tanese, N.; Telesnitsky, A.; Goff, S.P. Abortive reverse transcription by mutants of Moloney murine leukemia virus deficient in the reverse transcriptase-associated RNase H function. J. Virol. 1991, 65, 4387–4397. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, M.; Schulze, T.; Larder, B.A.; Moelling, K. Mutations within the RNase H domain of human immunodeficiency virus type 1 reverse transcriptase abolish virus infectivity. J. Gen. Virol. 1991, 72, 59–66. [Google Scholar] [CrossRef]
- Purohit, V.; Balakrishnan, M.; Kim, B.; Bambara, R.A. Evidence that HIV-1 reverse transcriptase employs the DNA 3' end-directed primary/secondary RNase H cleavage mechanism during synthesis and strand transfer. J. Biol. Chem. 2005, 280, 40534–40543. [Google Scholar] [CrossRef]
- DeStefano, J.J.; Mallaber, L.M.; Fay, P.J.; Bambara, R.A. Determinants of the RNase H cleavage specificity of human immunodeficiency virus reverse transcriptase. Nucl. Acids Res. 1993, 21, 4330–4338. [Google Scholar] [CrossRef]
- Furfine, E.S.; Reardon, J.E. Reverse transcriptase.RNase H from the human immunodeficiency virus. Relationship of the DNA polymerase and RNA hydrolysis activities. J. Biol. Chem. 1991, 266, 406–412. [Google Scholar] [CrossRef]
- Schultz, S.J.; Zhang, M.; Champoux, J.J. Recognition of internal cleavage sites by retroviral RNases H. J. Mol. Biol. 2004, 344, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Balakrishnan, M.; Palaniappan, C.; Fay, P.J.; Bambara, R.A. The sequential mechanism of HIV reverse transcriptase RNase H. J. Biol. Chem. 2000, 275, 37664–37671. [Google Scholar] [CrossRef] [PubMed]
- Delviks, K.A.; Hu, W.S.; Pathak, V.K. Psi- vectors: Murine leukemia virus-based self-inactivating and self-activating retroviral vectors. J. Virol. 1997, 71, 6218–6224. [Google Scholar] [CrossRef] [PubMed]
- Delviks, K.A.; Pathak, V.K. Development of murine leukemia virus-based self-activating vectors that efficiently delete the selectable drug resistance gene during reverse transcription. J. Virol. 1999, 73, 8837–8842. [Google Scholar] [CrossRef]
- Liu, S.; Abbondanzieri, E.A.; Rausch, J.W.; Le Grice, S.F.; Zhuang, X. Slide into action: Dynamic shuttling of HIV reverse transcriptase on nucleic acid substrates. Science 2008, 322, 1092–1097. [Google Scholar] [CrossRef]
- Anderson, J.A.; Bowman, E.H.; Hu, W.S. Retroviral recombination rates do not increase linearly with marker distance and are limited by the size of the recombining subpopulation. J. Virol. 1998, 72, 1195–1202. [Google Scholar] [CrossRef]
- Rhodes, T.; Wargo, H.; Hu, W.S. High rates of human immunodeficiency virus type 1 recombination: Near-random segregation of markers one kilobase apart in one round of viral replication. J. Virol. 2003, 77, 11193–11200. [Google Scholar] [CrossRef]
- Rhodes, T.D.; Nikolaitchik, O.; Chen, J.; Powell, D.; Hu, W.S. Genetic recombination of human immunodeficiency virus type 1 in one round of viral replication: Effects of genetic distance, target cells, accessory genes, and lack of high negative interference in crossover events. J. Virol. 2005, 79, 1666–1677. [Google Scholar] [CrossRef]
- Chen, J.; Powell, D.; Hu, W.S. High frequency of genetic recombination is a common feature of primate lentivirus replication. J. Virol. 2006, 80, 9651–9658. [Google Scholar] [CrossRef]
- Anderson, J.A.; Pathak, V.K.; Hu, W.S. Effect of the murine leukemia virus extended packaging signal on the rates and locations of retroviral recombination. J. Virol. 2000, 74, 6953–6963. [Google Scholar] [CrossRef]
- Zhuang, J.; Mukherjee, S.; Ron, Y.; Dougherty, J.P. High rate of genetic recombination in murine leukemia virus: Implications for influencing proviral ploidy. J. Virol. 2006, 80, 6706–6711. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.P.; Rhodes, T.D.; Chen, J.; Fu, W.; Hu, W.S. Identification of a major restriction in HIV-1 intersubtype recombination. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 9002–9007. [Google Scholar] [CrossRef]
- Clever, J.L.; Wong, M.L.; Parslow, T.G. Requirements for kissing-loop-mediated dimerization of human immunodeficiency virus RNA. J. Virol. 1996, 70, 5902–5908. [Google Scholar] [CrossRef] [PubMed]
- Muriaux, D.; Fosse, P.; Paoletti, J. A kissing complex together with a stable dimer is involved in the HIV-1Lai RNA dimerization process in vitro. Biochemistry 1996, 35, 5075–5082. [Google Scholar] [CrossRef] [PubMed]
- Paillart, J.C.; Skripkin, E.; Ehresmann, B.; Ehresmann, C.; Marquet, R. A loop-loop "kissing" complex is the essential part of the dimer linkage of genomic HIV-1 RNA. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 5572–5577. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.P.S.; Chen, J.; Nikolaitchik, O.A.; Hu, W.-S. Molecular determinants of HIV-1 intersubtype recombination potential. Virology 2007, 363, 437–446. [Google Scholar] [CrossRef]
- Moore, M.D.; Fu, W.; Nikolaitchik, O.; Chen, J.; Ptak, R.G.; Hu, W.S. Dimer initiation signal of human immunodeficiency virus type 1: Its role in partner selection during RNA copackaging and its effects on recombination. J. Virol. 2007, 81, 4002–4011. [Google Scholar] [CrossRef]
- Hussein, I.T.; Ni, N.; Galli, A.; Chen, J.; Moore, M.D.; Hu, W.S. Delineation of the preferences and requirements of the human immunodeficiency virus type 1 dimerization initiation signal by using an in vivo cell-based selection approach. J. Virol. 2010, 84, 6866–6875. [Google Scholar] [CrossRef]
- Moore, M.D.; Nikolaitchik, O.A.; Chen, J.; Hammarskjold, M.L.; Rekosh, D.; Hu, W.S. Probing the HIV-1 genomic RNA trafficking pathway and dimerization by genetic recombination and single virion analyses. PLoS Pathog. 2009, 5, e1000627. [Google Scholar] [CrossRef]
- Moore, M.D.; Hu, W.S. HIV-1 RNA dimerization: It takes two to tango. AIDS Rev. 2009, 11, 91–102. [Google Scholar]
- Paillart, J.C.; Shehu-Xhilaga, M.; Marquet, R.; Mak, J. Dimerization of retroviral RNA genomes: An inseparable pair. Nat. Rev. Microbiol. 2004, 2, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nikolaitchik, O.; Singh, J.; Wright, A.; Bencsics, C.E.; Coffin, J.M.; Ni, N.; Lockett, S.; Pathak, V.K.; Hu, W.S. High efficiency of HIV-1 genomic RNA packaging and heterozygote formation revealed by single virion analysis. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 13535–13540. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.A.; An, W.; King, S.R.; Telesnitsky, A. Nonrandom dimerization of murine leukemia virus genomic RNAs. J. Virol. 2004, 78, 12129–12139. [Google Scholar] [CrossRef] [PubMed]
- Kharytonchyk, S.A.; Kireyeva, A.I.; Osipovich, A.B.; Fomin, I.K. Evidence for preferential copackaging of Moloney murine leukemia virus genomic RNAs transcribed in the same chromosomal site. Retrovirology 2005, 2, 3. [Google Scholar] [CrossRef]
- Dang, Q.; Chen, J.; Unutmaz, D.; Coffin, J.M.; Pathak, V.K.; Powell, D.; KewalRamani, V.N.; Maldarelli, F.; Hu, W.S. Nonrandom HIV-1 infection and double infection via direct and cell-mediated pathways. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 632–637. [Google Scholar] [CrossRef]
- Chen, J.; Dang, Q.; Unutmaz, D.; Pathak, V.K.; Maldarelli, F.; Powell, D.; Hu, W.S. Mechanisms of nonrandom human immunodeficiency virus type 1 infection and double infection: Preference in virus entry is important but is not the sole factor. J. Virol. 2005, 79, 4140–4149. [Google Scholar] [CrossRef]
- Del Portillo, A.; Tripodi, J.; Najfeld, V.; Wodarz, D.; Levy, D.N.; Chen, B.K. Multiploid Inheritance of HIV-1 during Cell-to-Cell Infection. J. Virol. 2011, 85, 7169–7176. [Google Scholar] [CrossRef]
- Jung, A.; Maier, R.; Vartanian, J.P.; Bocharov, G.; Jung, V.; Fischer, U.; Meese, E.; Wain-Hobson, S.; Meyerhans, A. Recombination: Multiply infected spleen cells in HIV patients. Nature 2002, 418, 144. [Google Scholar] [CrossRef]
- Josefsson, L.; King, M.S.; Makitalo, B.; Brannstrom, J.; Shao, W.; Maldarelli, F.; Kearney, M.F.; Hu, W.S.; Chen, J.; Gaines, H.; et al. Majority of CD4+ T cells from peripheral blood of HIV-1-infected individuals contain only one HIV DNA molecule. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 11199–11204. [Google Scholar] [CrossRef]
- Chin, M.P.; Lee, S.K.; Chen, J.; Nikolaitchik, O.A.; Powell, D.A.; Fivash, M.J., Jr.; Hu, W.S. Long-range recombination gradient between HIV-1 subtypes B and C variants caused by sequence differences in the dimerization initiation signal region. J. Mol. Biol. 2008, 377, 1324–1333. [Google Scholar] [CrossRef]
- Nikolaitchik, O.A.; Galli, A.; Moore, M.D.; Pathak, V.K.; Hu, W.-S. Multiple barriers to recombination between divergent HIV-1 variants revealed by a dual-marker recombination assay. J. Mol. Biol. 2011, 407, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Kearney, M.; Nikolaitchik, O.A.; Yu, S.; Chin, M.P.S.; Maldarelli, F.; Coffin, J.M.; Pathak, V.K.; Hu, W.-S. Patterns of human immunodeficiency virus type 1 recombination ex vivo provide evidence for coadaptation of distant sites, resulting in purifying selection for intersubtype recombinants during replication. J. Virol. 2010, 84, 7651–7661. [Google Scholar] [CrossRef] [PubMed]
- Baird, H.A.; Galetto, R.; Gao, Y.; Simon-Loriere, E.; Abreha, M.; Archer, J.; Fan, J.; Robertson, D.L.; Arts, E.J.; Negroni, M. Sequence determinants of breakpoint location during HIV-1 intersubtype recombination. Nucl. Acids Res. 2006, 34, 5203–5216. [Google Scholar] [CrossRef]
- Baird, H.A.; Gao, Y.; Galetto, R.; Lalonde, M.; Anthony, R.M.; Giacomoni, V.; Abreha, M.; Destefano, J.J.; Negroni, M.; Arts, E.J. Influence of sequence identity and unique breakpoints on the frequency of intersubtype HIV-1 recombination. Retrovirology 2006, 3, 91. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Galetto, R.; Hamoudi, M.; Archer, J.; Lefeuvre, P.; Martin, D.P.; Robertson, D.L.; Negroni, M. Molecular mechanisms of recombination restriction in the envelope gene of the human immunodeficiency virus. PLoS Pathog. 2009, 5, e1000418. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Paraskevis, D.; Vandamme, A.M.; Magiorkinis, E.; Sypsa, V.; Hatzakis, A. In vivo characteristics of human immunodeficiency virus type 1 intersubtype recombination: Determination of hot spots and correlation with sequence similarity. J. Gen. Virol. 2003, 84, 2715–2722. [Google Scholar] [CrossRef]
- Archer, J.; Pinney, J.W.; Fan, J.; Simon-Loriere, E.; Arts, E.J.; Negroni, M.; Robertson, D.L. Identifying the important HIV-1 recombination breakpoints. PLoS Comput. Biol. 2008, 4, e1000178. [Google Scholar] [CrossRef]
- Fan, J.; Negroni, M.; Robertson, D.L. The distribution of HIV-1 recombination breakpoints. Infect. Genet. Evol. 2007, 7, 717–723. [Google Scholar] [CrossRef]
- Galli, A.; Lai, A.; Corvasce, S.; Saladini, F.; Riva, C.; Deho, L.; Caramma, I.; Franzetti, M.; Romano, L.; Galli, M.; et al. Recombination analysis and structure prediction show correlation between breakpoint clusters and RNA hairpins in the pol gene of human immunodeficiency virus type 1 unique recombinant forms. J. Gen. Virol. 2008, 89, 3119–3125. [Google Scholar] [CrossRef]
- Dykes, C.; Balakrishnan, M.; Planelles, V.; Zhu, Y.; Bambara, R.A.; Demeter, L.M. Identification of a preferred region for recombination and mutation in HIV-1 gag. Virology 2004, 326, 262–279. [Google Scholar] [CrossRef]
- Galetto, R.; Giacomoni, V.; Veron, M.; Negroni, M. Dissection of a circumscribed recombination hot spot in HIV-1 after a single infectious cycle. J. Biol. Chem. 2006, 281, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Palaniappan, C.; Wu, W.; Fay, P.J.; Bambara, R.A. Evidence for a unique mechanism of strand transfer from the transactivation response region of HIV-1. J. Biol. Chem. 1997, 272, 16769–16777. [Google Scholar] [CrossRef] [PubMed]
- Moumen, A.; Polomack, L.; Unge, T.; Veron, M.; Buc, H.; Negroni, M. Evidence for a mechanism of recombination during reverse transcription dependent on the structure of the acceptor RNA. J. Biol. Chem. 2003, 278, 15973–15982. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.M.; Dang, K.K.; Gorelick, R.J.; Leonard, C.W.; Bess Jr, J.W.; Swanstrom, R.; Burch, C.L.; Weeks, K.M. Architecture and secondary structure of an entire HIV-1 RNA genome. Nature 2009, 460, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Martin, D.P.; Weeks, K.M.; Negroni, M. RNA structures facilitate recombination-mediated gene swapping in HIV-1. J. Virol. 2010, 84, 12675–12682. [Google Scholar] [CrossRef] [PubMed]
- McCutchan, F.E. Global epidemiology of HIV. J. Med. Virol. 2006, 78, S7–S12. [Google Scholar] [CrossRef]
- Thomson, M.M.; Najera, R. Travel and the introduction of human immunodeficiency virus type 1 non-B subtype genetic forms into Western countries. Clin. Infect. Dis. 2001, 32, 1732–1737. [Google Scholar] [CrossRef]
- Thomson, M.M.; Najera, R. Molecular epidemiology of HIV-1 variants in the global AIDS pandemic: An update. AIDS Rev. 2005, 7, 210–224. [Google Scholar]
- Thomson, M.M.; Perez-Alvarez, L.; Najera, R. Molecular epidemiology of HIV-1 genetic forms and its significance for vaccine development and therapy. Lancet Infect. Dis. 2002, 2, 461–471. [Google Scholar] [CrossRef]
- Los Alamos HIV Sequence Database. Available online: http://www.hiv.lanl.gov/ (accessed on 8 August 2011).
- Oelrichs, R.B.; Crowe, S.M. The molecular epidemiology of HIV-1 in South and East Asia. Curr. HIV Res. 2003, 1, 239–248. [Google Scholar] [CrossRef]
- Abecasis, A.B.; Lemey, P.; Vidal, N.; de Oliveira, T.; Peeters, M.; Camacho, R.; Shapiro, B.; Rambaut, A.; Vandamme, A.-M. Recombination confounds the early evolutionary history of human immunodeficiency virus type 1: Subtype G is a circulating recombinant form. J. Virol. 2007, 81, 8543–8551. [Google Scholar] [CrossRef] [PubMed]
- Tee, K.K.; Pybus, O.G.; Li, X.J.; Han, X.; Shang, H.; Kamarulzaman, A.; Takebe, Y. Temporal and spatial dynamics of human immunodeficiency virus type 1 circulating recombinant forms 08_BC and 07_BC in Asia. J. Virol. 2008, 82, 9206–9215. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, M.T.; Ruibal, I.; Villahermosa, M.L.; Diaz, H.; Delgado, E.; Parga, E.V.; Perez-Alvarez, L.; de Armas, M.B.; Cuevas, L.; Medrano, L.; Noa, E.; Osmanov, S.; Najera, R.; Thomson, M.M. High HIV-1 genetic diversity in Cuba. AIDS 2002, 16, 1643–1653. [Google Scholar] [CrossRef]
- Delgado, E.; Thomson, M.M.; Villahermosa, M.L.; Sierra, M.; Ocampo, A.; Miralles, C.; Rodriguez-Perez, R.; Diz-Aren, J.; Ojea-de Castro, R.; Losada, E.; et al. Identification of a newly characterized HIV-1 BG intersubtype circulating recombinant form in Galicia, Spain, which exhibits a pseudotype-like virion structure. J. Acquir. Immune Defic. Syndr. 2002, 29, 536–543. [Google Scholar] [CrossRef]
- Thomson, M.M.; Delgado, E.; Manjón, N.; Ocampo, A.; Villahermosa, M.L.; Mariño, A.; Herrero, I.; Cuevas, M.T.; Vázquez-de Parga, E.; Pérez-Alvarez, L.; et al. Spanish Group for Antiretroviral Studies in Galicia HIV-1 genetic diversity in Galicia Spain: BG intersubtype recombinant viruses circulating among injecting drug users. AIDS 2001, 15, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. AIDS 2006, 20, W13–W23. [Google Scholar] [CrossRef]
- Bourlet, T.; Signori-Schmuck, A.; Roche, L.; Icard, V.; Saoudin, H.; Trabaud, M.A.; Tardy, J.C.; Morand, P.; Pozzetto, B.; Ecochard, R.; et al. HIV-1 load comparison using four commercial real-time assays. J. Clin. Microbiol. 2011, 49, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Church, D.; Gregson, D.; Lloyd, T.; Klein, M.; Beckthold, B.; Laupland, K.; Gill, M.J. Comparison of the RealTime HIV-1, COBAS TaqMan 48 v1.0, Easy Q v1.2, and Versant v3.0 assays for Determination of HIV-1 Viral Loads in a Cohort of Canadian Patients with Diverse HIV Subtype Infections. J. Clin. Microbiol. 2010, 49, 118–124. [Google Scholar] [CrossRef]
- Foglieni, B.; Candotti, D.; Guarnori, I.; Raffaele, L.; Berzuini, A.; Spreafico, M.; Orani, A.; Rossotti, R.; Rossi, D.; Allain, J.P.; et al. A cluster of human immunodeficiency virus Type 1 recombinant form escaping detection by commercial genomic amplification assays. Transfusion 2011, 51, 719–730. [Google Scholar] [CrossRef]
- Korenromp, E.L.; Williams, B.G.; Schmid, G.P.; Dye, C. Clinical prognostic value of RNA viral load and CD4 cell counts during untreated HIV-1 infection—A quantitative review. PLoS One 2009, 4, e5950. [Google Scholar] [CrossRef]
- Mellors, J.W.; Rinaldo, C.R., Jr.; Gupta, P.; White, R.M.; Todd, J.A.; Kingsley, L.A. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science 1996, 272, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Arien, K.K.; Abraha, A.; Quinones-Mateu, M.E.; Kestens, L.; Vanham, G.; Arts, E.J. The replicative fitness of primary human immunodeficiency virus type 1 (HIV-1) group M, HIV-1 group O, and HIV-2 isolates. J. Virol. 2005, 79, 8979–8990. [Google Scholar] [CrossRef] [PubMed]
- Njai, H.F.; Gali, Y.; Vanham, G.; Clybergh, C.; Jennes, W.; Vidal, N.; Butel, C.; Mpoudi-Ngolle, E.; Peeters, M.; Arien, K.K. The predominance of human immunodeficiency virus type 1 (HIV-1) circulating recombinant form 02 (CRF02_AG) in West Central Africa may be related to its replicative fitness. Retrovirology 2006, 3, 40. [Google Scholar] [CrossRef]
- Konings, F.A.; Burda, S.T.; Urbanski, M.M.; Zhong, P.; Nadas, A.; Nyambi, P.N. Human immunodeficiency virus type 1 (HIV-1) circulating recombinant form 02_AG (CRF02_AG) has a higher in vitro replicative capacity than its parental subtypes A and G. J. Med. Virol. 2006, 78, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.Y.; Takebe, Y.; Weniger, B.G.; Luo, C.C.; Kalish, M.L.; Auwanit, W.; Yamazaki, S.; Gayle, H.D.; Young, N.L.; Schochetman, G. Independent introduction of two major HIV-1 genotypes into distinct high-risk populations in Thailand. Lancet 1993, 341, 1171–1174. [Google Scholar] [CrossRef]
- Hudgens, M.G.; Longini, I.M., Jr.; Vanichseni, S.; Hu, D.J.; Kitayaporn, D.; Mock, P.A.; Halloran, M.E.; Satten, G.A.; Choopanya, K.; Mastro, T.D. Subtype-specific transmission probabilities for human immunodeficiency virus type 1 among injecting drug users in Bangkok, Thailand. Am. J. Epidemiol. 2002, 155, 159–168. [Google Scholar] [CrossRef]
- Laurent, C.; Bourgeois, A.; Faye, M.A.; Mougnutou, R.; Seydi, M.; Gueye, M.; Liegeois, F.; Kane, C.T.; Butel, C.; Mbuagbaw, J.; et al. No difference in clinical progression between patients infected with the predominant human immunodeficiency virus type 1 circulating recombinant form (CRF) 02_AG strain and patients not infected with CRF02_AG, in Western and West-Central Africa: A four-year prospective multicenter study. J. Infect. Dis. 2002, 186, 486–492. [Google Scholar]
- Easterbrook, P.J.; Smith, M.; Mullen, J.; O'Shea, S.; Chrystie, I.; de Ruiter, A.; Tatt, I.D.; Geretti, A.M.; Zuckerman, M. Impact of HIV-1 viral subtype on disease progression and response to antiretroviral therapy. J. Int. AIDS Soc. 2010, 13, 4. [Google Scholar] [CrossRef]
- Baeten, J.M.; Chohan, B.; Lavreys, L.; Chohan, V.; McClelland, R.S.; Certain, L.; Mandaliya, K.; Jaoko, W.; Overbaugh, J. HIV-1 subtype D infection is associated with faster disease progression than subtype A in spite of similar plasma HIV-1 loads. J. Infect. Dis. 2007, 195, 1177–1180. [Google Scholar] [CrossRef]
- Vergne, L.; Peeters, M.; Mpoudi-Ngole, E.; Bourgeois, A.; Liegeois, F.; Toure-Kane, C.; Mboup, S.; Mulanga-Kabeya, C.; Saman, E.; Jourdan, J.; et al. Genetic diversity of protease and reverse transcriptase sequences in non-subtype-b human immunodeficiency virus type 1 strains: Evidence of many minor drug resistance mutations in treatment-naive patients. J. Clin. Microbiol. 2000, 38, 3919–3925. [Google Scholar] [CrossRef]
- Kantor, R.; Katzenstein, D.A.; Efron, B.; Carvalho, A.P.; Wynhoven, B.; Cane, P.; Clarke, J.; Sirivichayakul, S.; Soares, M.A.; Snoeck, J.; et al. Impact of HIV-1 subtype and antiretroviral therapy on protease and reverse transcriptase genotype: Results of a global collaboration. PLoS Med. 2005, 2, e112. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M. HIV-1 subtypes: Epidemiology and significance for HIV management. Curr. Opin. Infect. Dis. 2006, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M.; Harrison, L.; Green, H.; Sabin, C.; Hill, T.; Fearnhill, E.; Pillay, D.; Dunn, D. Effect of HIV-1 subtype on virologic and immunologic response to starting highly active antiretroviral therapy. Clin. Infect. Dis. 2009, 48, 1296–1305. [Google Scholar] [CrossRef]
- Bannister, W.P.; Ruiz, L.; Loveday, C.; Vella, S.; Zilmer, K.; Kjaer, J.; Knysz, B.; Phillips, A.N.; Mocroft, A. HIV-1 subtypes and response to combination antiretroviral therapy in Europe. Antivir. Ther. 2006, 11, 707–715. [Google Scholar] [PubMed]
- Bocket, L.; Cheret, A.; Deuffic-Burban, S.; Choisy, P.; Gerard, Y.; de la Tribonniere, X.; Viget, N.; Ajana, F.; Goffard, A.; Barin, F.; Mouton, Y.; Yazdanpanah, Y. Impact of human immunodeficiency virus type 1 subtype on first-line antiretroviral therapy effectiveness. Antivir. Ther. 2005, 10, 247–254. [Google Scholar] [CrossRef]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Maldarelli, F.; Hase, S.; Takebe, Y.; Pathak, V.K. Subtype-specific differences in the human immunodeficiency virus type 1 reverse transcriptase connection subdomain of CRF01_AE are associated with higher levels of resistance to 3'-azido-3'-deoxythymidine. J. Virol. 2009, 83, 8502–8513. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Posada, D.; Crandall, K.A.; Holmes, E.C. The causes and consequences of HIV evolution. Nat. Rev. Genet. 2004, 5, 52–61. [Google Scholar] [CrossRef]
- Batorsky, R.; Kearney, M.F.; Palmer, S.E.; Maldarelli, F.; Rouzine, I.M.; Coffin, J.M. Estimate of effective recombination rate and average selection coefficient for HIV in chronic infection. Proc. Natl. Acad. Sci. U. S. A. 2011. [Google Scholar] [CrossRef]
- Kearney, M.; Maldarelli, F.; Shao, W.; Margolick, J.B.; Daar, E.S.; Mellors, J.W.; Rao, V.; Coffin, J.M.; Palmer, S. Human immunodeficiency virus type 1 population genetics and adaptation in newly infected individuals. J. Virol. 2009, 83, 2715–2727. [Google Scholar] [CrossRef]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 7552–7557. [Google Scholar] [CrossRef]
- Li, H.; Bar, K.J.; Wang, S.; Decker, J.M.; Chen, Y.; Sun, C.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Learn, G.H.; Morgan, C.J.; et al. High Multiplicity Infection by HIV-1 in men who have sex with men. PLoS Pathog. 2010, 6, e1000890. [Google Scholar] [CrossRef] [PubMed]
- Streeck, H.; Li, B.; Poon, A.F.; Schneidewind, A.; Gladden, A.D.; Power, K.A.; Daskalakis, D.; Bazner, S.; Zuniga, R.; Brander, C.; et al. Immune-driven recombination and loss of control after HIV superinfection. J. Exp. Med. 2008, 205, 1789–1796. [Google Scholar] [CrossRef]
- Reynolds, M.R.; Weiler, A.M.; Piaskowski, S.M.; Kolar, H.L.; Hessell, A.J.; Weiker, M.; Weisgrau, K.L.; Leon, E.J.; Rogers, W.E.; Makowsky, R.; et al. Macaques vaccinated with simian immunodeficiency virus SIVmac239Delta nef delay acquisition and control replication after repeated low-dose heterologous SIV challenge. J. Virol. 2010, 84, 9190–9199. [Google Scholar] [CrossRef] [PubMed]
- Gundlach, B.R.; Lewis, M.G.; Sopper, S.; Schnell, T.; Sodroski, J.; Stahl-Hennig, C.; Uberla, K. Evidence for recombination of live, attenuated immunodeficiency virus vaccine with challenge virus to a more virulent strain. J. Virol. 2000, 74, 3537–3542. [Google Scholar] [CrossRef] [PubMed]
- Metzner, K.J.; Moretto, W.J.; Donahoe, S.M.; Jin, X.; Gettie, A.; Montefiori, D.C.; Marx, P.A.; Binley, J.M.; Nixon, D.F.; Connor, R.I. Evaluation of CD8+ T-cell and antibody responses following transient increased viraemia in rhesus macaques infected with live, attenuated simian immunodeficiency virus. J. Gen. Virol. 2005, 86, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.R.; Weiler, A.M.; Weisgrau, K.L.; Piaskowski, S.M.; Furlott, J.R.; Weinfurter, J.T.; Kaizu, M.; Soma, T.; Leon, E.J.; MacNair, C.; et al. Macaques vaccinated with live-attenuated SIV control replication of heterologous virus. J. Exp. Med. 2008, 205, 2537–2550. [Google Scholar] [CrossRef]
- Fischer, W.; Perkins, S.; Theiler, J.; Bhattacharya, T.; Yusim, K.; Funkhouser, R.; Kuiken, C.; Haynes, B.; Letvin, N.L.; Walker, B.D.; et al. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nat. Med. 2007, 13, 100–106. [Google Scholar] [CrossRef]
- Nickle, D.C.; Rolland, M.; Jensen, M.A.; Pond, S.L.; Deng, W.; Seligman, M.; Heckerman, D.; Mullins, J.I.; Jojic, N. Coping with viral diversity in HIV vaccine design. PLoS Comput. Biol. 2007, 3, e75. [Google Scholar] [CrossRef]
- Santra, S.; Liao, H.X.; Zhang, R.; Muldoon, M.; Watson, S.; Fischer, W.; Theiler, J.; Szinger, J.; Balachandran, H.; Buzby, A.; et al. Mosaic vaccines elicit CD8+ T lymphocyte responses that confer enhanced immune coverage of diverse HIV strains in monkeys. Nat. Med. 2010, 16, 324–328. [Google Scholar] [CrossRef]
- Barouch, D.H.; O'Brien, K.L.; Simmons, N.L.; King, S.L.; Abbink, P.; Maxfield, L.F.; Sun, Y.H.; La Porte, A.; Riggs, A.M.; Lynch, D.M.; et al. Mosaic HIV-1 vaccines expand the breadth and depth of cellular immune responses in rhesus monkeys. Nat. Med. 2010, 16, 319–323. [Google Scholar] [CrossRef]
Figure 1.
Reverse Transcription. (A) General structures of retroviral DNA and RNA genomes; (B) Processes required to complete the synthesis of viral DNA from the viral RNA genome. Red, DNA; blue, RNA; green, tRNA primer; arrow, direction of DNA synthesis. RNase H degradation, blue dashes.
Figure 1.
Reverse Transcription. (A) General structures of retroviral DNA and RNA genomes; (B) Processes required to complete the synthesis of viral DNA from the viral RNA genome. Red, DNA; blue, RNA; green, tRNA primer; arrow, direction of DNA synthesis. RNase H degradation, blue dashes.

Figure 2.
Dynamic Copy Choice Model. Shown are two direct repeats (orange box) undergoing an intramolecular template switch during minus-strand DNA synthesis (thick black arrow and line) resulting in deletion of the directly repeated sequence. RNA strand (thin black line) is depicted as “looped”, aligning the homologous directly repeated sequences during reverse transcription. (A) Direct repeat deletion by wild type reverse transcriptase (RT). During reverse transcription, degradation of the RNA template allows the newly synthesized DNA strand to anneal to the acceptor template (upstream direct repeat). The nascent DNA growing point switches templates, reverse transcription continues and one of the direct repeats is deleted; (B) RTs with a slow polymerase will increase template switching; (C) RTs with a slow RNase H will decrease template switching.
Figure 2.
Dynamic Copy Choice Model. Shown are two direct repeats (orange box) undergoing an intramolecular template switch during minus-strand DNA synthesis (thick black arrow and line) resulting in deletion of the directly repeated sequence. RNA strand (thin black line) is depicted as “looped”, aligning the homologous directly repeated sequences during reverse transcription. (A) Direct repeat deletion by wild type reverse transcriptase (RT). During reverse transcription, degradation of the RNA template allows the newly synthesized DNA strand to anneal to the acceptor template (upstream direct repeat). The nascent DNA growing point switches templates, reverse transcription continues and one of the direct repeats is deleted; (B) RTs with a slow polymerase will increase template switching; (C) RTs with a slow RNase H will decrease template switching.

Figure 3.
Relationship Between Direct Repeat Size and Deletion Frequency. Left: Summary table showing the reported deletion frequencies of direct repeats from MLV, SNV and HIV-1; right: correlation between the size of the direct repeat and the frequency of deletion.
Figure 3.
Relationship Between Direct Repeat Size and Deletion Frequency. Left: Summary table showing the reported deletion frequencies of direct repeats from MLV, SNV and HIV-1; right: correlation between the size of the direct repeat and the frequency of deletion.

Figure 4.
Single-cycle Vector to Monitor Viral Infection and the In Vivo Intramolecular Template Switching Rate. An HIV vector containing direct repeats (horizontal arrows) of the middle portion of the gfp gene (the “F” portion) is mobilized by co-transfection with vectors containing gag-pol and envelope. The frequency of a homologous template switch during reverse transcription is measured by reconstitution of the gfp gene. LTR, long terminal repeats.
Figure 4.
Single-cycle Vector to Monitor Viral Infection and the In Vivo Intramolecular Template Switching Rate. An HIV vector containing direct repeats (horizontal arrows) of the middle portion of the gfp gene (the “F” portion) is mobilized by co-transfection with vectors containing gag-pol and envelope. The frequency of a homologous template switch during reverse transcription is measured by reconstitution of the gfp gene. LTR, long terminal repeats.

Figure 5.
Retroviral Recombination Rates. X-axis, nucleotide distances between genetic markers; Y-axis, percent of theoretical maximum observable recombination rate [79]. Measured recombination rates from lentiviruses (dotted oval in blue) and gammretroviruses (dotted oval in black).
Figure 5.
Retroviral Recombination Rates. X-axis, nucleotide distances between genetic markers; Y-axis, percent of theoretical maximum observable recombination rate [79]. Measured recombination rates from lentiviruses (dotted oval in blue) and gammretroviruses (dotted oval in black).

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Delviks-Frankenberry, K.; Galli, A.; Nikolaitchik, O.; Mens, H.; Pathak, V.K.; Hu, W.-S. Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses 2011, 3, 1650-1680. https://doi.org/10.3390/v3091650
AMA Style
Delviks-Frankenberry K, Galli A, Nikolaitchik O, Mens H, Pathak VK, Hu W-S. Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses. 2011; 3(9):1650-1680. https://doi.org/10.3390/v3091650
Chicago/Turabian StyleDelviks-Frankenberry, Krista, Andrea Galli, Olga Nikolaitchik, Helene Mens, Vinay K. Pathak, and Wei-Shau Hu. 2011. "Mechanisms and Factors that Influence High Frequency Retroviral Recombination" Viruses 3, no. 9: 1650-1680. https://doi.org/10.3390/v3091650