First Report of Cowpea Mild Mottle Carlavirus on Yardlong Bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
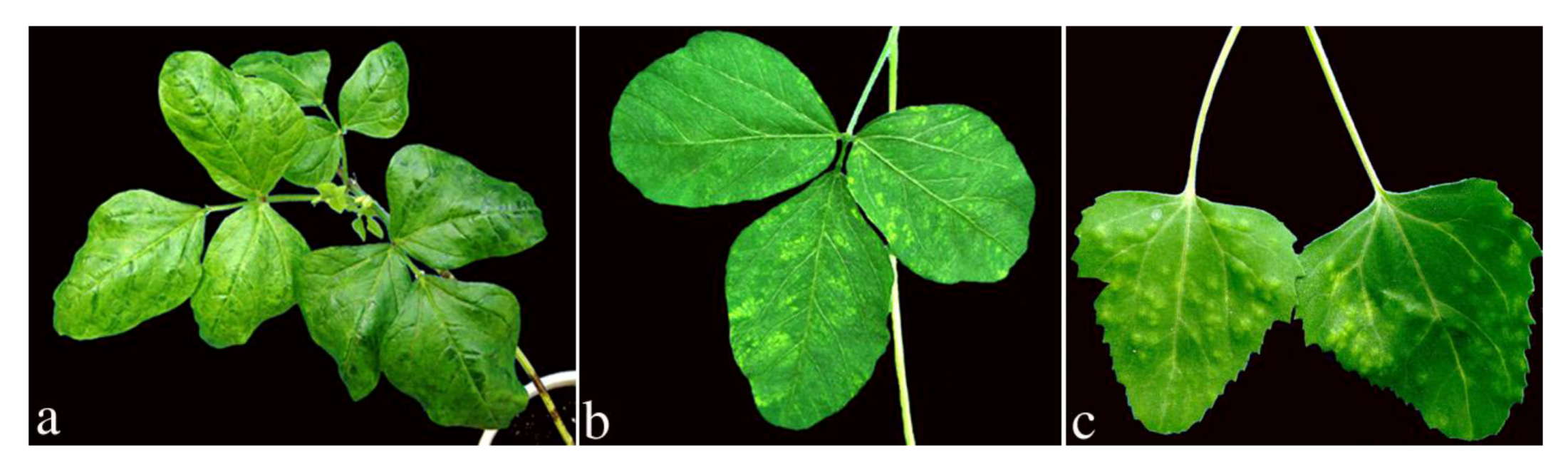
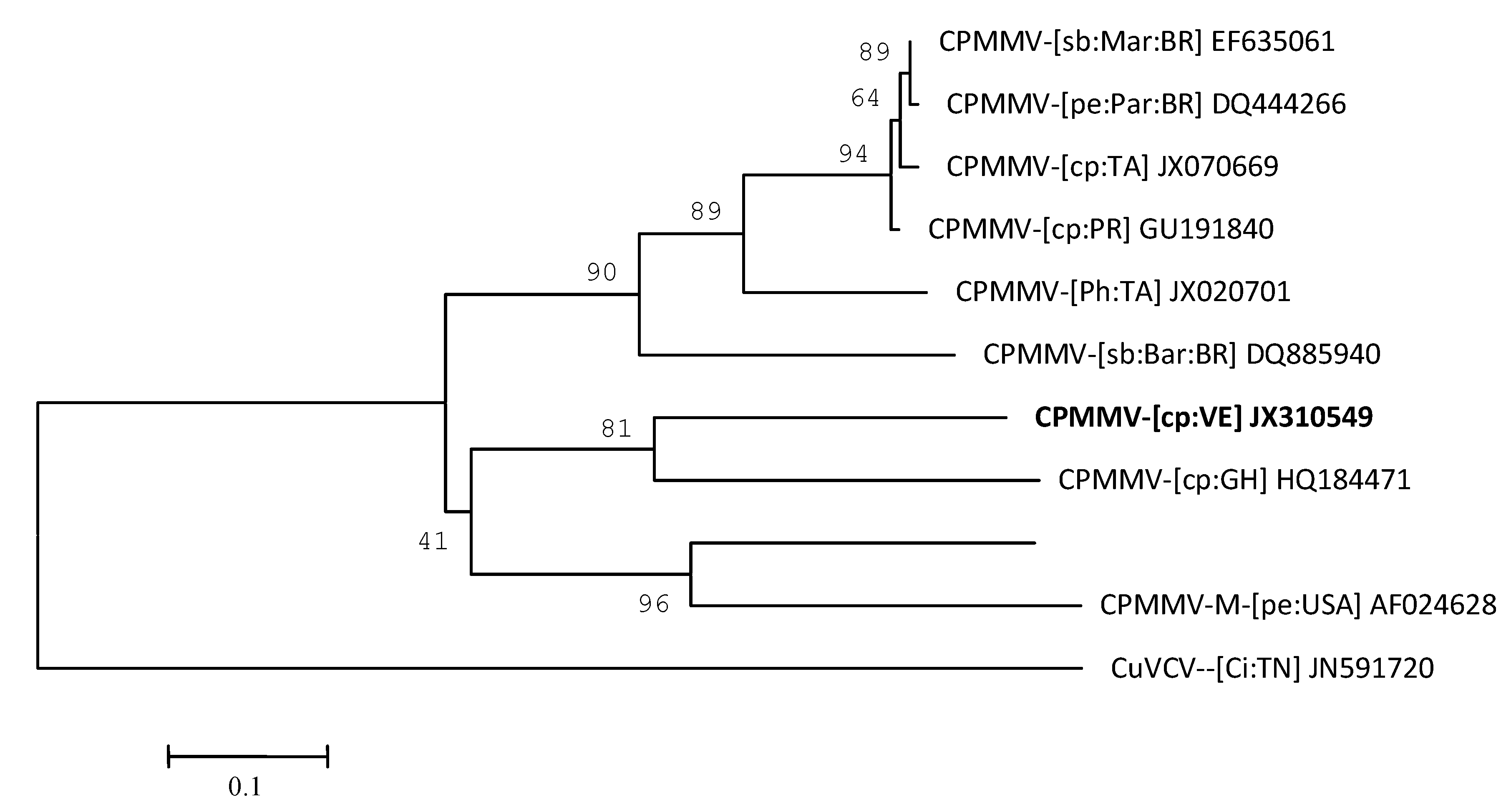
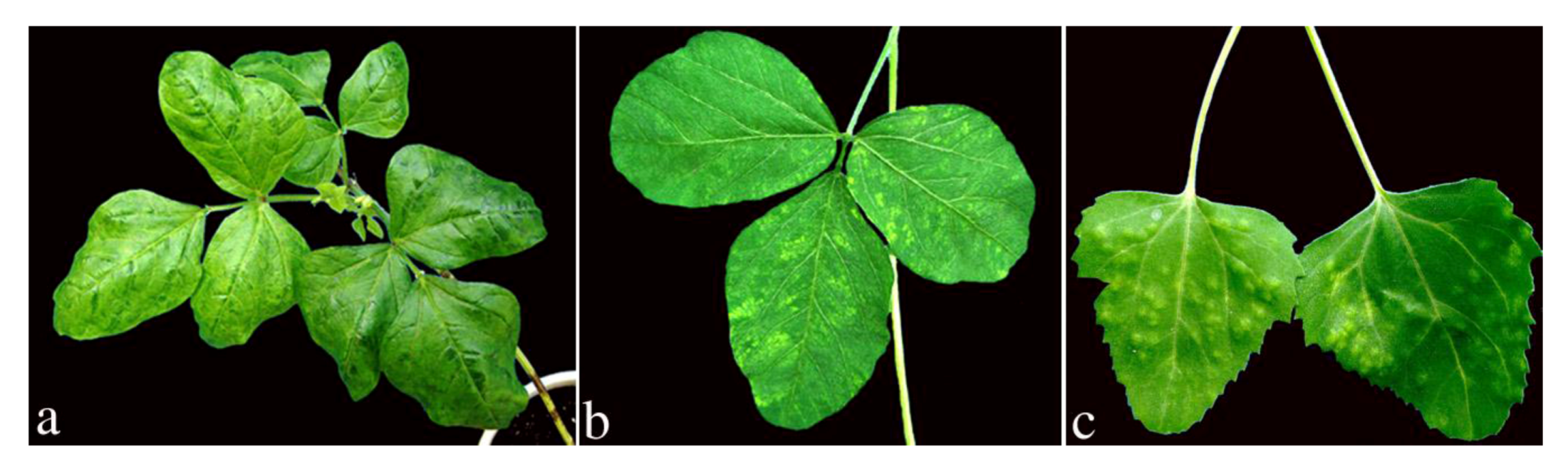
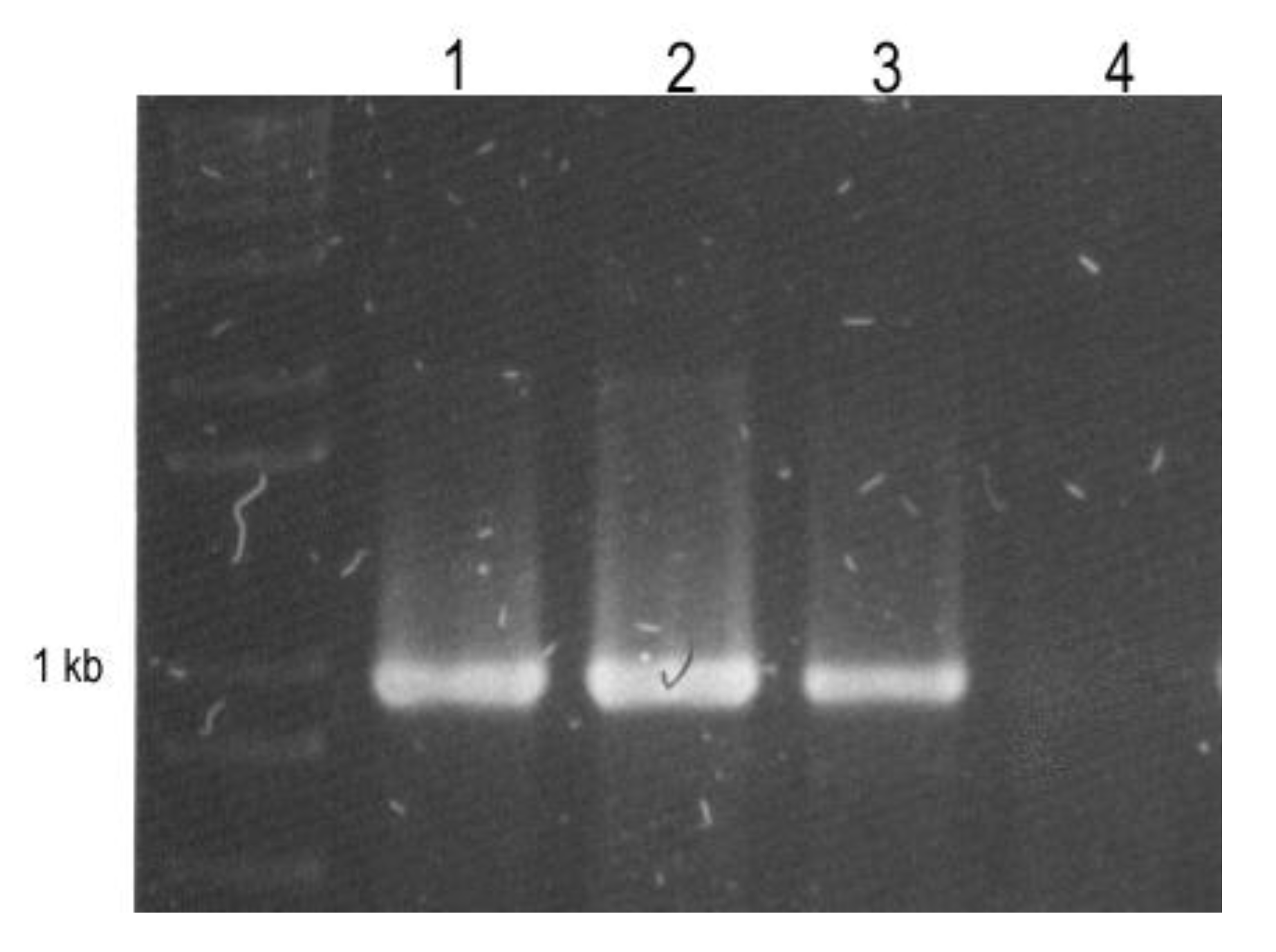
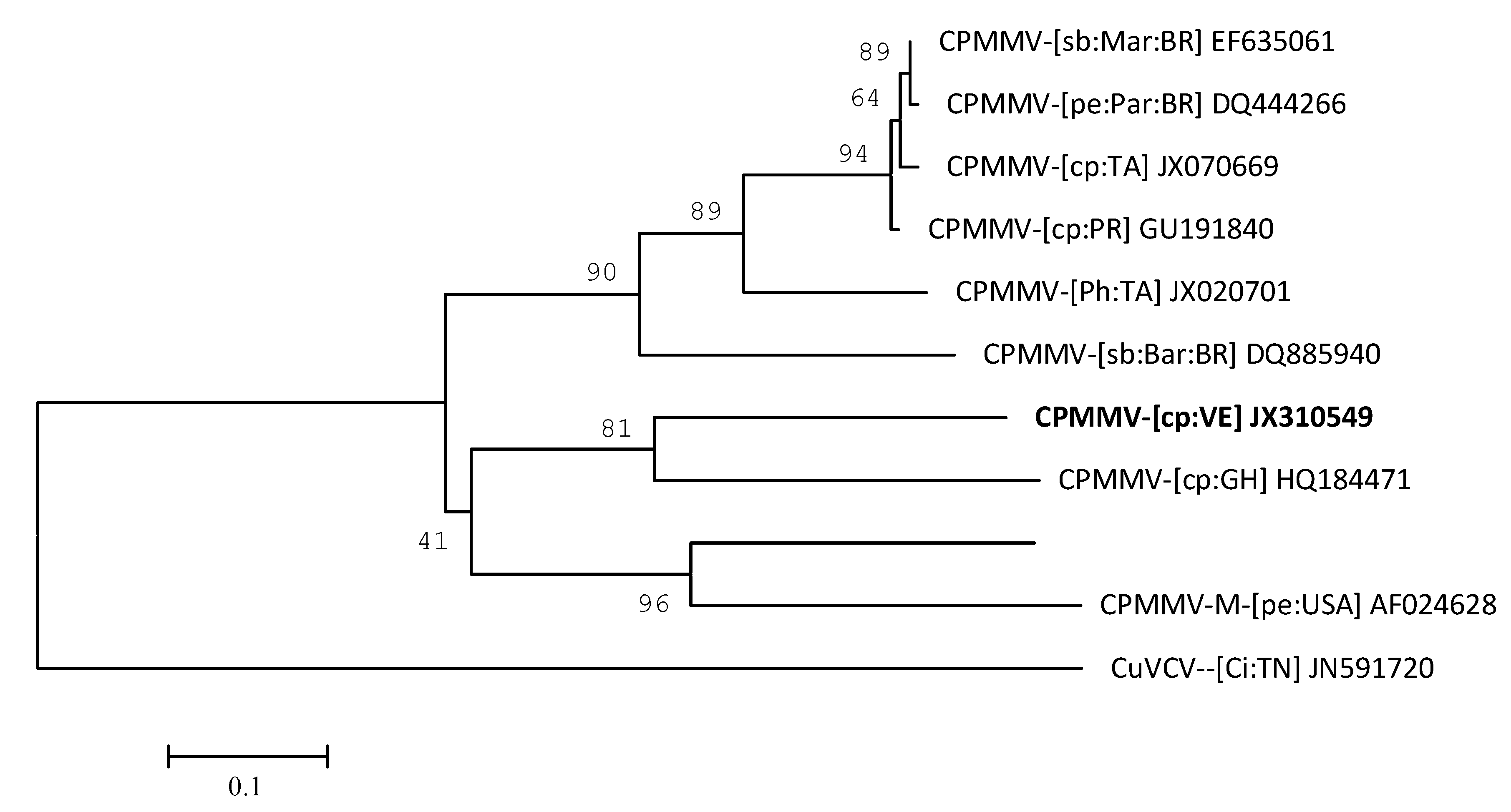
2. Results and Discussion

3. Experimental
3.1. Virus Source and Biological Assays
3.2. Virus Particle and Cytopathology
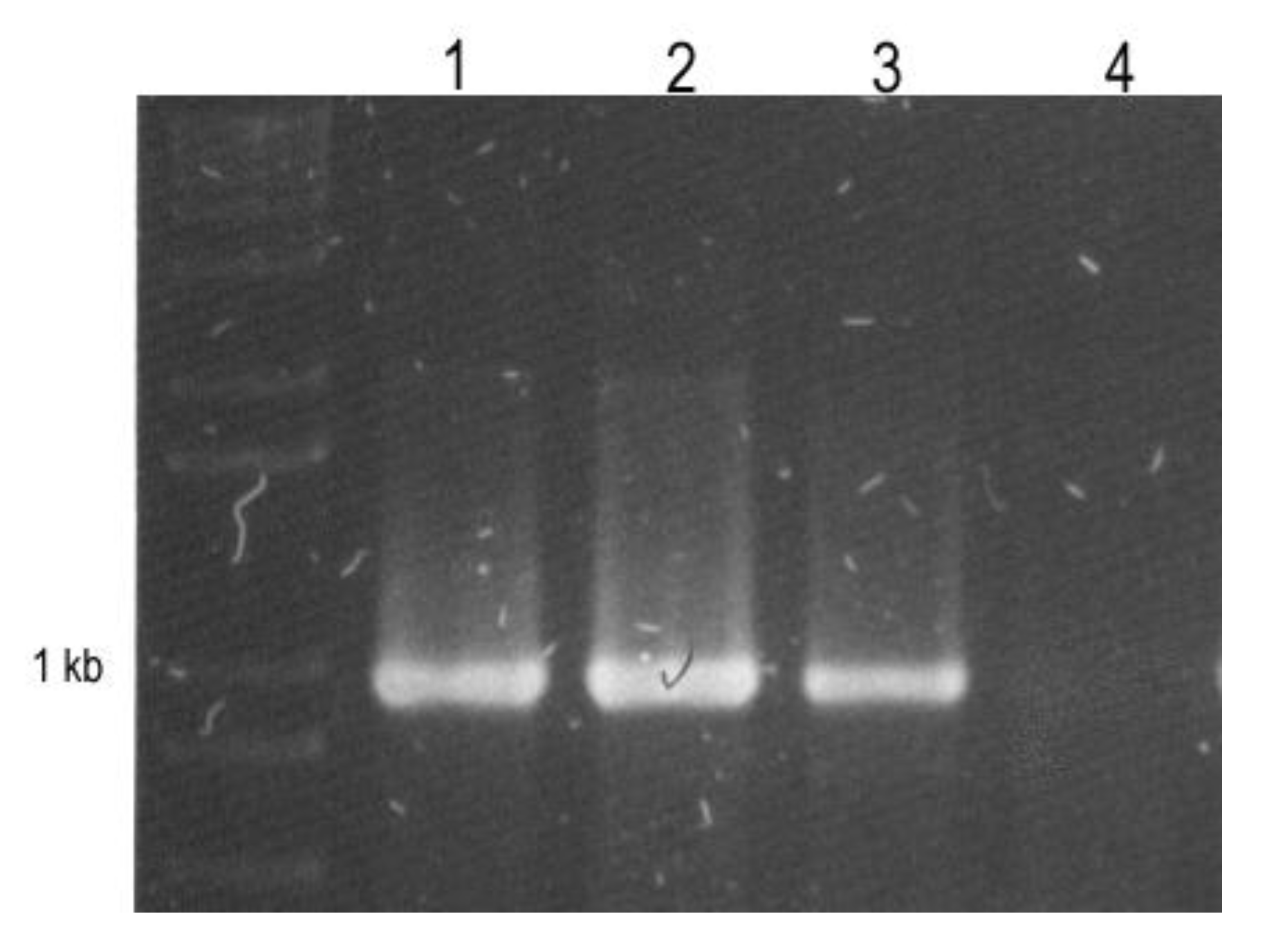
3.3. Reverse Transcription and RT-PCR Detection.
4. Conclusions
Conflict of Interest
Acknowledgements
References and Notes
- Brunt, A.A.; Kenten, R.H. Cowpea mild mottle, a newly recognized virus infecting cowpeas (Vigna unguiculata) in Ghana. Ann. Appl. Biol. 1973, 74, 67–74. [Google Scholar] [CrossRef]
- Anno-Nyako, F.O. Identification and partial characterization of a mild mottle disease in soybean (Glycine max (L.) Meril) in Nigeria.
- Brunt, A.A.; Phillips, S. “Fuzzy vein” a disease of tomato (Lycopersicum esculentum) in Western Nigeria induced by Cowpea mild mottle virus. Trop. Agric. 1981, 58, 177–180. [Google Scholar]
- Mink, G.I.; Keswani, C.L. First report of Cowpea mild mottle virus on bean and mung bean in Tanzania. Plant Dis. 1987, 71, 557. [Google Scholar]
- Salem, N.M.; Ehlers, J.A.; Roberts, P.A.; Ng, J.C.K. Biological and molecular diagnosis of seedborne viruses in cowpea germplasm of geographically diverse sub-Saharan origins. Plant Pathol. 2012, 59, 773–784. [Google Scholar]
- Thouvenel, J.C.; Monsarrat, A.; Fauquet, C. Isolation of cowpea mild mottle virus from diseased soybean in Ivory Cost. Plant Dis. 1982, 66, 336–337. [Google Scholar] [CrossRef]
- Antignus, Y.; Cohen, S. Purification and some properties of a new strain of cowpea mild mottle virus in Israel. Ann. Appl. Biol. 1987, 110, 563–569. [Google Scholar] [CrossRef]
- Nolt, B.L.; Rajeshwari, R. Properties of a Cowpea mild mottle virus isolate from groundnut. Indian Phytopathol. 1987, 40, 22–26. [Google Scholar]
- Reddy, D.V.R. Crop profile. Groundnut viruses and virus diseases: Distribution, identification and control. Rev. Plant Pathol. 1991, 70, 665–678. [Google Scholar]
- Shahraeen, N. Studies on a virus causing mosaic diseases of soybean (cv. MACS-13). Indian Phytopathol. 1989, 42, 338. [Google Scholar]
- Gaspar, J.O.; Beriam, L.O.S.; Alves, M.N.; Oliveira, A.R.; Costa, A.S. Serological identity of bean angular mosaic and cowpea mild mottle viruses. Fitopatologia Brasileira 1985, 10, 195–199. [Google Scholar]
- Hartman, G.L.; Sinclair, J.B.; Rupe, J.C. Compendium of Soybean Disease, 4th ed; The American Phytopathological Society: St. Paul, MN, USA, 1999; p. 182. [Google Scholar]
- Tavasoli, M.; Shahraeen, N.; Ghorbani, S.H. Serological and RT-PCR detection of Cowpea mild mottle carlavirus infecting soybean. J. Gen. Mol. Virol. 2009, 1, 7–11. [Google Scholar]
- Diouf, D. Recent advances in cowpea [Vigna unguiculata (L.) Walp.] “omics” research. Afr. J. Biotechnol. 2011, 10, 2803–2819. [Google Scholar]
- Fery, R.L. New Opportunities in Vigna. Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 224–228. [Google Scholar]
- Hampton, R.O.; Albrechtsen, S.E.; Mathur, S.B. Seed health (viruses) of Vigna unguiculata selections from developing countries. Seed Sci. Technol. 1992, 20, 23–38. [Google Scholar]
- Brunt, A.; Crabtree, K.; Dallwitz, M.; Gibbs, A.; Watson, L. Viruses of Plants: Descriptions and Lists from the VIDE Database. CAB International: Wallingford, UK, 1996. [Google Scholar]
- Badge, J.; Brunt, A.; Carson, R.; Dagless, E.; Karamagioli, M.; Phillips, S.; Seal, S.; Turner, R.; Foster, G.D. A carlavirus-specific PCR primer and partial nucleotide sequence provides further evidence for the recognition of cowpea mild mottle virus as a whitefly-transmitted carlavirus. Eur. J. Plant Pathol. 1996, 102, 305–310. [Google Scholar] [CrossRef]
- Almeida, A.M.R.; Piuga, F.F.; Marin, S.R.; Kitajima, E.W.; Gaspar, J.O.; Moraes, T.G. Detection and partial characterization of a carlavirus causing stem necrosis of soybeans in Brazil. Fitopatologia Brasileira 2005, 30, 191–194. [Google Scholar]
- Laguna, I.G.; Arneodo, J.D.; Rodriguez-Pardina, P.; Kitajima, E.W. Cowpea mild mottle virus infecting soybean crops in North-Western Argentina. Fitopatologia Brasileira 2006, 31, 317. [Google Scholar] [CrossRef]
- Gaspar, J.O.; Belintani, P.; Almeida, A.M.R.; Kitajima, E.W. A degenerate primer allows amplification of part of the 3'-terminus of three distinct carlavirus species. J. Virol. Methods 2008, 481, 283–285. [Google Scholar]
- Costa, A.S.; Gaspar, J.O.; Vega, J. Mosaico angular do feijoeiro Jalo causado por um carlavirus transmitido pela mosca branca Bemisia tabaci. Fitopatologia Brasileira 1983, 8, 325–337. [Google Scholar]
- Zheng, H.Y.; Chen, J.; Adams, M.J.; Chen, J.P. Complete nucleotide sequence and affinities of the genomic RNA of Narcissus common latent virus (genus Carlavirus). Arch. Virol. 2006, 151, 1667–1672. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment, editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 85–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brito, M.; Fernández-Rodríguez, T.; Garrido, M.J.; Mejías, A.; Romano, M.; Marys, E. First Report of Cowpea Mild Mottle Carlavirus on Yardlong Bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela. Viruses 2012, 4, 3804-3811. https://doi.org/10.3390/v4123804
Brito M, Fernández-Rodríguez T, Garrido MJ, Mejías A, Romano M, Marys E. First Report of Cowpea Mild Mottle Carlavirus on Yardlong Bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela. Viruses. 2012; 4(12):3804-3811. https://doi.org/10.3390/v4123804
Chicago/Turabian StyleBrito, Miriam, Thaly Fernández-Rodríguez, Mario José Garrido, Alexander Mejías, Mirtha Romano, and Edgloris Marys. 2012. "First Report of Cowpea Mild Mottle Carlavirus on Yardlong Bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela" Viruses 4, no. 12: 3804-3811. https://doi.org/10.3390/v4123804