Genetic Modification of Hematopoietic Stem Cells as a Therapy for HIV/AIDS

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Preliminary Results
2.1. Studies in Murine and Nonhuman Primate Models
2.2. Hematopoietic Stem cell Transplantation in HIV-1 Infected Patients & Ongoing Clinical Trials

3. Establishing an ‘AIDS Patient Model’ in Nonhuman Primates
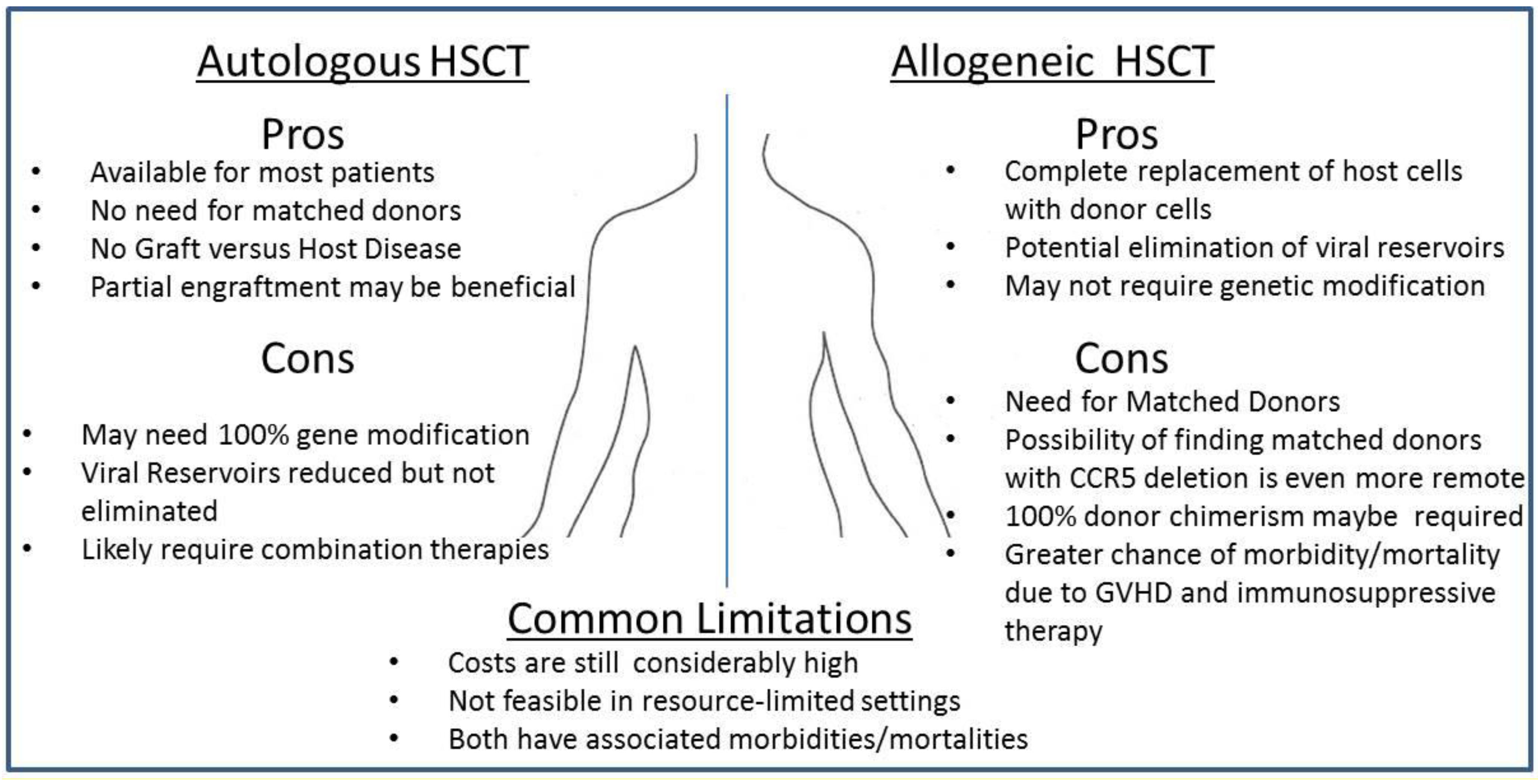
3.1. Autologous Transplantation and the Missing Ingredient: Graft-Versus-Host Disease?
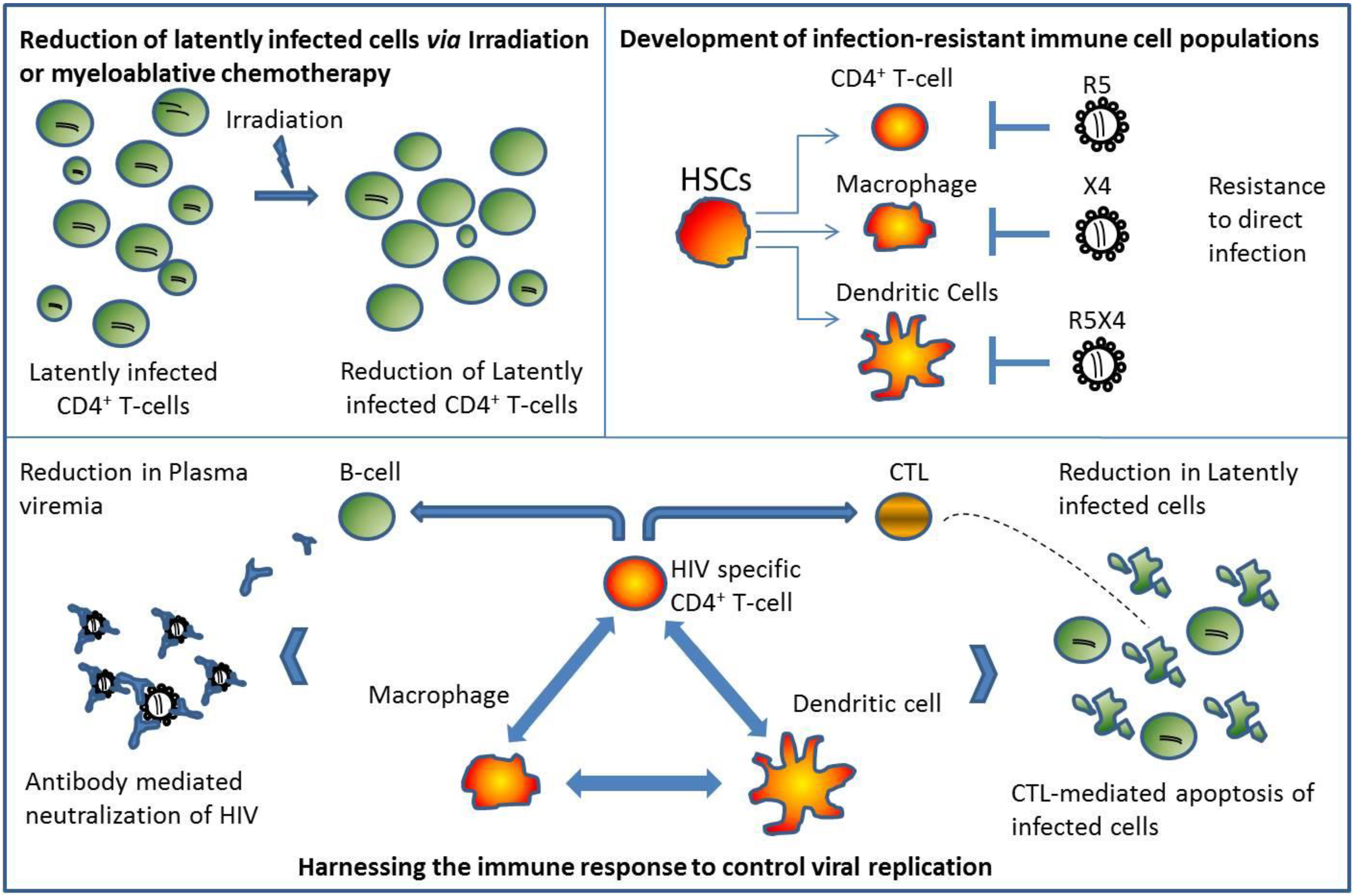
3.2. Reduction of Viral Reservoirs following Myeloablative Conditioning & Non-HSC Derived Macrophages
3.3. The X-factor: Harnessing the Immune System

4. Opinion: Reaching for a Cure
Acknowledgments
Conflicts of Interest
References and Notes
- Walensky, R.P.; Paltiel, A.D.; Losina, E.; Mercincavage, L.M.; Schackman, B.R.; Sax, P.E.; Weinstein, M.C.; Freedberg, K.A. The survival benefits of AIDS treatment in the United States. J. Infect. Dis. 2006, 194, 11–19. [Google Scholar]
- Mills, E.J.; Bakanda, C.; Birungi, J.; Chan, K.; Ford, N.; Cooper, C.L.; Nachega, J.B.; Dybul, M.; Hogg, R.S. Life expectancy of persons receiving combination antiretroviral therapy in low-income countries: a cohort analysis from Uganda. Ann. Intern. Med. 2011, 155, 209–216. [Google Scholar] [CrossRef]
- Eisele, E.; Siliciano, R.F. Redefining the viral reservoirs that prevent HIV-1 eradication (Review). Immunity 2012, 37, 377–388. [Google Scholar] [CrossRef]
- Wong, J.K.; Hezareh, M.; Gunthard, H.F.; Havlir, D.V.; Ignacio, C.C.; Spina, C.A.; Richman, D.D. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 1997, 278, 1291–1295. [Google Scholar]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; Gallant, J.; Markowitz, M.; Ho, D.D.; Richman, D.D.; Siliciano, R.F. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef]
- Chun, T.W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.; Baseler, M.; Lloyd, A.L.; Nowak, M.A.; Fauci, A.S. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. PNAS 1997, 94, 13193–13197. [Google Scholar]
- Kiem, H.-P.; Jerome, K.R.; Deeks, S.G.; McCune, J.M. Hematopoietic-stem-cell-based gene therapy for HIV disease (Review). Cell Stem Cell 2012, 10, 137–147. [Google Scholar] [CrossRef]
- Hutter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Mussig, A.; Allers, K.; Schneider, T.; Hofmann, J.; Kucherer, C.; Blau, O.; Blau, I.W.; Hofmann, W.K.; Thiel, E. Long-term control of HIV by CCR5 Delta32/Delta32 stem-cell transplantation. N. Engl. J. Med. 2009, 360, 692–698. [Google Scholar] [CrossRef]
- Allers, K.; Hutter, G.; Hofmann, J.; Loddenkemper, C.; Rieger, K.; Thiel, E.; Schneider, T. Evidence for the cure of HIV infection by CCR5Δ32/Δ32 stem cell transplantation. Blood 2011, 117, 2791–2799. [Google Scholar] [CrossRef]
- Hutter, G.; Thiel, E. Allogeneic transplantation of CCR5-deficient progenitor cells in a patient with HIV infection: an update after 3 years and the search for patient no. 2. AIDS 2011, 25, 273–274. [Google Scholar]
- Yukl, S.A.; Boritz, E.; Busch, M.; Bentsen, C.; Chun, T.W.; Douek, D.; Eisele, E.; Haase, A.; Ho, Y.C.; Hutter, G.; Justement, J.S.; Keating, S.; Lee, T.H.; Li, P.; Murray, D.; Palmer, S.; Pilcher, C.; Pillai, S.; Price, R.W.; Rothenberger, M.; Schacker, T.; Siliciano, J.; Siliciano, R.; Sinclair, E.; Strain, M.; Wong, J.; Richman, D.; Deeks, S.G. Challenges in detecting HIV persistence during potentially curative interventions: a study of the Berlin patient. PLoS Pathogens 2013, 9, e1003347. [Google Scholar] [CrossRef]
- Hoxie, J.A.; June, C.H. Novel cell and gene therapies for HIV (Review). Cold Spring Harbor Perspectives in Medicine 2012, 2. [Google Scholar]
- Burnett, J.C.; Zaia, J.A.; Rossi, J.J. Creating genetic resistance to HIV (Review). Curr. Opin. Immunol. 2012, 24, 625–632. [Google Scholar] [CrossRef]
- Hutter, G.; Zaia, J.A. Allogeneic haematopoietic stem cell transplantation in patients with human immunodeficiency virus: the experiences of more than 25 years (Review). Clin Exp Immunol 2011, 163, 284–295. [Google Scholar] [CrossRef]
- Resino, S.; Perez, A.; Seoane, E.; Serrano, D.; Berenguer, J.; Balsalobre, P.; Gomez-Chacon, G.F.; Diez-Martin, J.L.; Munoz-Fernandez, M.A. Short communication: Immune reconstitution after autologous peripheral blood stem cell transplantation in HIV-infected patients: might be better than expected? AIDS Res Hum Retroviruses 2007, 23, 543–548. [Google Scholar] [CrossRef]
- Statham, S.; Morgan, R.A. Gene therapy clinical trials for HIV (Review). Current Opinion in Molecular Therapeutics 1999, 1, 430–436. [Google Scholar]
- Simonelli, C.; Zanussi, S.; Pratesi, C.; Rupolo, M.; Talamini, R.; Caffau, C.; Bortolin, M.T.; Tedeschi, R.; Basaglia, G.; Mazzucato, M.; Manuele, R.; Vaccher, E.; Spina, M.; Tirelli, U.; Michieli, M.; de Paoli, P. Immune recovery after autologous stem cell transplantation is not different for HIV-infected versus HIV-uninfected patients with relapsed or refractory lymphoma. Clin. Infect. Dis. 2010, 50, 1672–1679. [Google Scholar]
- Holt, N.; Wang, J.; Kim, K.; Friedman, G.; Wang, X.; Taupin, V.; Crooks, G.M.; Kohn, D.B.; Gregory, P.D.; Holmes, M.C.; Cannon, P.M. Human hematopoietic stem/progenitor cells modified by zinc-finger nucleases targeted to CCR5 control HIV-1 in vivo. Nat. Biotechnol. 2010, 28, 839–847. [Google Scholar] [CrossRef]
- Kimpel, J.; Braun, S.E.; Qiu, G.; Wong, F.E.; Conolle, M.; Schmitz, J.E.; Brendel, C.; Humeau, L.M.; Dropulic, B.; Rossi, J.J.; Berger, A.; von Laer, D.; Johnson, R.P. Survival of the fittest: positive selection of CD4+ T cells expressing a membrane-bound fusion inhibitor following HIV-1 infection. PLoS ONE 2010, 5, e12357. [Google Scholar] [CrossRef]
- Walker, J.E.; Chen, R.X.; McGee, J.; Nacey, C.; Pollard, R.B.; Abedi, M.; Bauer, G.; Nolta, J.A.; Anderson, J.S. Generation of an HIV-1-resistant immune system with CD34(+) hematopoietic stem cells transduced with a triple-combination anti-HIV lentiviral vector. J. Virol. 2012, 86, 5719–5729. [Google Scholar] [CrossRef]
- Younan, P.M.; Polacino, P.; Kowalski, J.P.; Peterson, C.W.; Maurice, N.J.; Williams, N.P.; Ho, O.; Trobridge, G.D.; von Laer, D.; Prlic, M.; Beard, B.C.; DeRosa, S.; Hu, S.L.; Kiem, H.P. Positive selection of mC46-expressing CD4+ T cells and maintenance of virus specific immunity in a primate AIDS model. Blood 2013, 122, 179–187. [Google Scholar] [CrossRef]
- Hassett, J.M.; Zaroulis, C.G.; Greenberg, M.L.; Siegal, F.P. Bone marrow transplantation in AIDS. N. Engl. J. Med. 1983, 309, 665. [Google Scholar]
- Lane, H.C.; Masur, H.; Longo, D.L.; Klein, H.G.; Rook, A.H.; Quinnan, G.V., Jr.; Steis, R.G.; Macher, A.; Whalen, G.; Edgar, L.C. Partial immune reconstitution in a patient with the acquired immunodeficiency syndrome. N. Engl. J. Med. 1984, 311, 1099–1103. [Google Scholar] [CrossRef]
- Wolf, T.; Rickerts, V.; Staszewski, S.; Kriener, S.; Wassmann, B.; Bug, G.; Bickel, M.; Gute, P.; Brodt, H.R.; Martin, H. First case of successful allogeneic stem cell transplantation in an HIV-patient who acquired severe aplastic anemia. Haematologica 2007, 92, e56–e58. [Google Scholar] [CrossRef]
- Avettand-Fenoel, V.; Mahlaoui, N.; Chaix, M.L.; Milliancourt, C.; Burgard, M.; Cavazzana-Calvo, M.; Rouzioux, C.; Blanche, S. Failure of bone marrow transplantation to eradicate HIV reservoir despite efficient HAART. AIDS 2007, 21, 776–777. [Google Scholar] [CrossRef]
- Henrich, T.J.; Hu, Z.; Li, J.Z.; Sciaranghella, G.; Busch, M.P.; Keating, S.M.; Gallien, S.; Lin, N.H.; Giguel, F.F.; Lavoie, L.; Ho, V.T.; Armand, P.; Soiffer, R.J.; Sagar, M.; LaCasce, A.S.; Kuritzkes, D.R. Long-term reduction in peripheral blood HIV type 1 reservoirs following reduced-intensity conditioning allogeneic stem cell transplantation. J. Infect. Dis. 2013, 207, 1694–1702. [Google Scholar] [CrossRef]
- Cillo, A.R.; Krishnan, A.; Mitsuyasu, R.T.; McMahon, D.K.; Li, S.; Rossi, J.J.; Zaia, J.A.; Mellors, J.W. Plasma viremia and cellular HIV-1 DNA persist despite autologous hematopoietic stem cell transplantation for HIV-related lymphoma. J Acquir. Immune Defic. Syndr. 2013, 63, 438–441. [Google Scholar]
- Tomonari, A.; Takahashi, S.; Shimohakamada, Y.; Ooi, J.; Takasugi, K.; Ohno, N.; Konuma, T.; Uchimaru, K.; Tojo, A.; Odawara, T.; Nakamura, T.; Iwamoto, A.; Asano, S. Unrelated cord blood transplantation for a human immunodeficiency virus-1-seropositive patient with acute lymphoblastic leukemia. Bone Marrow Transplant. 2005, 36, 261–262. [Google Scholar]
- Kang, E.M.; De Witte, M.; Malech, H.; Morgan, R.A.; Phang, S.; Carter, C.; Leitman, S.F.; Childs, R.; Barrett, A.J.; Little, R.; Tisdale, J.F. Nonmyeloablative conditioning followed by transplantation of genetically modified HLA-matched peripheral blood progenitor cells for hematologic malignancies in patients with acquired immunodeficiency syndrome. Blood 2002, 99, 698–701. [Google Scholar] [CrossRef]
- Mitsuyasu, R.T.; Zack, J.A.; MacPherson, J.L.; Symonds, G.P. Phase I/II clinical trials using gene-modified adult hematopoietic stem cells for HIV: lessons learnt. Stem Cells International 2011, 2011, 393698. [Google Scholar]
- Evans, D.T.; Silvestri, G. Nonhuman primate models in AIDS research. Current Opinion in HIV and AIDS 2013, 8, 255–261. [Google Scholar]
- Serrano, D.; Carrion, R.; Balsalobre, P.; Miralles, P.; Berenguer, J.; Buno, I.; Gomez-Pineda, A.; Ribera, J.M.; Conde, E.; Diez-Martin, J.L. HIV-associated lymphoma successfully treated with peripheral blood stem cell transplantation. Exp. Hematol. 2005, 33, 487–494. [Google Scholar] [CrossRef]
- Krishnan, A.; Molina, A.; Zaia, J.; Nademanee, A.; Kogut, N.; Rosenthal, J.; Woo, D.; Forman, S.J. Autologous stem cell transplantation for HIV-associated lymphoma. Blood 2001, 98, 3857–3859. [Google Scholar]
- Krishnan, A.; Molina, A.; Zaia, J.; Smith, D.; Vasquez, D.; Kogut, N.; Falk, P.M.; Rosenthal, J.; Alvarnas, J.; Forman, S.J. Durable remissions with autologous stem cell transplantation for high-risk HIV-associated lymphomas. Blood 2005, 105, 874–878. [Google Scholar] [CrossRef]
- Haynes, B.F.; Markert, M.L.; Sempowski, G.D.; Patel, D.D.; Hale, L.P. The role of the thymus in immune reconstitution in aging, bone marrow transplantation, and HIV-1 infection (Review). Annu. Rev. Immunol. 2000, 18, 529–560. [Google Scholar] [CrossRef]
- Napolitano, L.A.; Grant, R.M.; Deeks, S.G.; Schmidt, D.; De Rosa, S.C.; Herzenberg, L.A.; Herndier, B.G.; Andersson, J.; McCune, J.M. Increased production of IL-7 accompanies HIV-1-mediated T-cell depletion: implications for T-cell homeostasis. Nat. Med. 2001, 7, 73–79. [Google Scholar]
- Llano, A.; Barretina, J.; Gutierrez, A.; Blanco, J.; Cabrera, C.; Clotet, B.; Este, J.A. Interleukin-7 in plasma correlates with CD4 T-cell depletion and may be associated with emergence of syncytium-inducing variants in human immunodeficiency virus type 1-positive individuals. J. Virol. 2001, 75, 10319–10325. [Google Scholar] [CrossRef]
- Rethi, B.; Fluur, C.; Atlas, A.; Krzyzowska, M.; Mowafi, F.; Grutzmeier, S.; De Milito, A.; Bellocco, R.; Falk, K.I.; Rajnavolgyi, E.; Chiodi, F. Loss of IL-7Ralpha is associated with CD4 T-cell depletion, high interleukin-7 levels and CD28 down-regulation in HIV infected patients. AIDS 2005, 19, 2077–2086. [Google Scholar] [CrossRef]
- Schluns, K.S.; Kieper, W.C.; Jameson, S.C.; LeFrancois, L. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nat. Immunol. 2000, 1, 426–432. [Google Scholar] [CrossRef]
- Gaardbo, J.C.; Hartling, H.J.; Gerstoft, J.; Nielsen, S.D. Incomplete immune recovery in HIV infection: mechanisms, relevance for clinical care, and possible solutions (Review). Clinical & Developmental Immunology 2012, 2012, 670957. [Google Scholar]
- Akashi, K.; Kondo, M.; von Freeden-Jeffry, U.; Murray, R.; Weissman, I.L. Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Cell 1997, 89, 1033–1041. [Google Scholar] [CrossRef]
- Di Santo, J.P.; Aifantis, I.; Rosmaraki, E.; Garcia, C.; Feinberg, J.; Fehling, H.J.; Fischer, A.; von Boehmer, H.; Rocha, B. The common cytokine receptor gamma chain and the pre-T cell receptor provide independent but critically overlapping signals in early alpha/beta T cell development. J. Exp. Med. 1999, 189, 563–574. [Google Scholar] [CrossRef]
- Pallard, C.; Stegmann, A.P.; van Kleffens, T.; Smart, F.; Venkitaraman, A.; Spits, H. Distinct roles of the phosphatidylinositol 3-kinase and STAT5 pathways in IL-7-mediated development of human thymocyte precursors. Immunity 1999, 10, 525–535. [Google Scholar]
- Chun, T.W.; Finzi, D.; Margolick, J.; Chadwick, K.; Schwartz, D.; Siliciano, R.F. In vivo fate of HIV-1-infected T cells: quantitative analysis of the transition to stable latency. Nat. Med. 1995, 1, 1284–1290. [Google Scholar] [CrossRef]
- Chun, T.W.; Carruth, L.; Finzi, D.; Shen, X.; DiGiuseppe, J.A.; Taylor, H.; Hermankova, M.; Chadwick, K.; Margolick, J.; Quinn, T.C.; Kuo, Y.H.; Brookmeyer, R.; Zeiger, M.A.; Barditch-Crovo, P.; Siliciano, R.F. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nature 1997, 387, 183–188. [Google Scholar]
- Richman, D.D.; Margolis, D.M.; Delaney, M.; Greene, W.C.; Hazuda, D.; Pomerantz, R.J. The challenge of finding a cure for HIV infection (Review). Science 2009, 323, 1304–1307. [Google Scholar] [CrossRef]
- Ramratnam, B.; Mittler, J.E.; Zhang, L.; Boden, D.; Hurley, A.; Fang, F.; Macken, C.A.; Perelson, A.S.; Markowitz, M.; Ho, D.D. The decay of the latent reservoir of replication-competent HIV-1 is inversely correlated with the extent of residual viral replication during prolonged anti-retroviral therapy. Nat. Med. 2000, 6, 82–85. [Google Scholar] [CrossRef]
- Zhang, L.; Ramratnam, B.; Tenner-Racz, K.; He, Y.; Vesanen, M.; Lewin, S.; Talal, A.; Racz, P.; Perelson, A.S.; Korber, B.T.; Markowitz, M.; Ho, D.D. Quantifying residual HIV-1 replication in patients receiving combination antiretroviral therapy. N. Engl. J. Med. 1999, 340, 1605–1613. [Google Scholar] [CrossRef]
- Ho, D.D. Toward HIV eradication or remission: the tasks ahead. Science 1998, 280, 1866–1867. [Google Scholar] [CrossRef]
- Fogg, D.K.; Sibon, C.; Miled, C.; Jung, S.; Aucouturier, P.; Littman, D.R.; Cumano, A.; Geissmann, F. A clonogenic bone marrow progenitor specific for macrophages and dendritic cells. [Erratum appears in Science. 2006 Mar 3;311(5765):1242]. Science 2006, 311, 83–87. [Google Scholar] [CrossRef]
- Liu, K.; Victora, G.D.; Schwickert, T.A.; Guermonprez, P.; Meredith, M.M.; Yao, K.; Chu, F.F.; Randolph, G.J.; Rudensky, A.Y.; Nussenzweig, M. In vivo analysis of dendritic cell development and homeostasis. Science 2009, 324, 392–397. [Google Scholar]
- Schulz, C.; Gomez Perdiguero, E.; Chorro, L.; Szabo-Rogers, H.; Cagnard, N.; Kierdorf, K.; Prinz, M.; Wu, B.; Jacobsen, S.E.; Pollard, J.W.; Frampton, J.; Liu, K.J.; Geissmann, F. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 2012, 336, 86–90. [Google Scholar] [CrossRef]
- Alliot, F.; Godin, I.; Pessac, B. Microglia derive from progenitors, originating from the yolk sac, and which proliferate in the brain. Brain Res. 1999, Dev. Brain Res. 117, 145–152. [Google Scholar] [CrossRef]
- Klein, I.; Cornejo, J.C.; Polakos, N.K.; John, B.; Wuensch, S.A.; Topham, D.J.; Pierce, R.H.; Crispe, I.N. Kupffer cell heterogeneity: functional properties of bone marrow derived and sessile hepatic macrophages. Blood 2007, 110, 4077–4085. [Google Scholar] [CrossRef]
- Chorro, L.; Sarde, A.; Li, M.; Woollard, K.J.; Chambon, P.; Malissen, B.; Kissenpfennig, A.; Barbaroux, J.B.; Groves, R.; Geissmann, F. Langerhans cell (LC) proliferation mediates neonatal development, homeostasis, and inflammation-associated expansion of the epidermal LC network. J. Exp. Med. 2009, 206, 3089–3100. [Google Scholar] [CrossRef]
- Merad, M.; Manz, M.G.; Karsunky, H.; Wagers, A.; Peters, W.; Charo, I.; Weissman, I.L.; Cyster, J.G.; Engleman, E.G. Langerhans cells renew in the skin throughout life under steady-state conditions. [Erratum appears in Nat Immunol 2003 Jan;4(1):92]. Nat. Immunol. 2002, 3, 1135–1141. [Google Scholar]
- Ajami, B.; Bennett, J.L.; Krieger, C.; Tetzlaff, W.; Rossi, F.M. Local self-renewal can sustain CNS microglia maintenance and function throughout adult life. Nature Neuroscience 2007, 10, 1538–1543. [Google Scholar]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; Samokhvalov, I.M.; Merad, M. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef]
- Hunt, P.W.; Martin, J.N.; Sinclair, E.; Bredt, B.; Hagos, E.; Lampiris, H.; Deeks, S.G. T cell activation is associated with lower CD4+ T cell gains in human immunodeficiency virus-infected patients with sustained viral suppression during antiretroviral therapy. J. Infect. Dis. 2003, 187, 1534–1543. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; Blazar, B.R.; Rodriguez, B.; Teixeira-Johnson, L.; Landay, A.; Martin, J.N.; Hecht, F.M.; Picker, L.J.; Lederman, M.M.; Deeks, S.G.; Douek, D.C. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar]
- Estes, J.; Baker, J.V.; Brenchley, J.M.; Khoruts, A.; Barthold, J.L.; Bantle, A.; Reilly, C.S.; Beilman, G.J.; George, M.E.; Douek, D.C.; Haase, A.T.; Schacker, T.W. Collagen deposition limits immune reconstitution in the gut. J. Infect. Dis. 2008, 198, 456–464. [Google Scholar]
- Schacker, T.W.; Reilly, C.; Beilman, G.J.; Taylor, J.; Skarda, D.; Krason, D.; Larson, M.; Haase, A.T. Amount of lymphatic tissue fibrosis in HIV infection predicts magnitude of HAART-associated change in peripheral CD4 cell count. AIDS 2005, 19, 2169–2171. [Google Scholar] [CrossRef]
- Stranford, S.A.; Ong, J.C.; Martinez-Marino, B.; Busch, M.; Hecht, F.M.; Kahn, J.; Levy, J.A. Reduction in CD8+ cell noncytotoxic anti-HIV activity in individuals receiving highly active antiretroviral therapy during primary infection. Proc. Natl. Acad. Sci. USA 2001, 98, 597–602. [Google Scholar]
- Conrad, J.A.; Ramalingam, R.K.; Duncan, C.B.; Smith, R.M.; Wei, J.; Barnett, L.; Simons, B.C.; Lorey, S.L.; Kalams, S.A. Antiretroviral therapy reduces the magnitude and T cell receptor repertoire diversity of HIV-specific T cell responses without changing T cell clonotype dominance. J. Virol. 2012, 86, 4213–4221. [Google Scholar]
- Porichis, F.; Kaufmann, D.E. HIV-specific CD4 T cells and immune control of viral replication (Review). Current Opinion in HIV & AIDS 2011, 6, 174–180. [Google Scholar]
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining chronic viral infection (Review). Cell 2009, 138, 30–50. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Lu, B.; Gerard, C.; Iwasaki, A. CD8(+) T lymphocyte mobilization to virus-infected tissue requires CD4(+) T-cell help. Nature 2009, 462, 510–513. [Google Scholar] [CrossRef]
- Herbeuval, J.P.; Smith, N.; Theze, J. Characteristics of plasmacytoid dendritic cell and CD4+ T cell in HIV elite controllers (Review). Clinical & Developmental Immunology 2012, 2012, 869505. [Google Scholar]
- Iannello, A.; Tremblay, C.; Routy, J.P.; Boulassel, M.R.; Toma, E.; Ahmad, A. Decreased levels of circulating IL-21 in HIV-infected AIDS patients: correlation with CD4+ T-cell counts. Viral Immunology 2008, 21, 385–388. [Google Scholar] [CrossRef]
- Iannello, A.; Boulassel, M.R.; Samarani, S.; Debbeche, O.; Tremblay, C.; Toma, E.; Routy, J.P.; Ahmad, A. Dynamics and consequences of IL-21 production in HIV-infected individuals: a longitudinal and cross-sectional study. J. Immunol. 2010, 184, 114–126. [Google Scholar] [CrossRef]
- Yue, F.Y.; Lo, C.; Sakhdari, A.; Lee, E.Y.; Kovacs, C.M.; Benko, E.; Liu, J.; Song, H.; Jones, R.B.; Sheth, P.; Chege, D.; Kaul, R.; Ostrowski, M.A. HIV-specific IL-21 producing CD4+ T cells are induced in acute and chronic progressive HIV infection and are associated with relative viral control. [Erratum appears in J Immunol. 2010 Aug 15;185(4):2632-3]. J. Immunol. 2010, 185, 498–506. [Google Scholar]
- Santoro, M.M.; Perno, C.F. HIV-1 genetic variability and clinical implications. ISRN Microbiology 2013, 2013, 481314. [Google Scholar]
- McIlroy, D. Do HIV-specific CTL continue to have an antiviral function during antiretroviral therapy? If not, why not, and what can be done about it? Frontiers in Immunology 2013, 4, 52. [Google Scholar]
- Deeks, S.G.; Schweighardt, B.; Wrin, T.; Galovich, J.; Hoh, R.; Sinclair, E.; Hunt, P.; McCune, J.M.; Martin, J.N.; Petropoulos, C.J.; Hecht, F.M. Neutralizing antibody responses against autologous and heterologous viruses in acute versus chronic human immunodeficiency virus (HIV) infection: evidence for a constraint on the ability of HIV to completely evade neutralizing antibody responses. J. Virol. 2006, 80, 6155–6164. [Google Scholar]
- Alter, G.; Heckerman, D.; Schneidewind, A.; Fadda, L.; Kadie, C.M.; Carlson, J.M.; Oniangue-Ndza, C.; Martin, M.; Li, B.; Khakoo, S.I.; Carrington, M.; Allen, T.M.; Altfeld, M. HIV-1 adaptation to NK-cell-mediated immune pressure. Nature 2011, 476, 96–100. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Younan, P.; Kowalski, J.; Kiem, H.-P. Genetic Modification of Hematopoietic Stem Cells as a Therapy for HIV/AIDS. Viruses 2013, 5, 2946-2962. https://doi.org/10.3390/v5122946
Younan P, Kowalski J, Kiem H-P. Genetic Modification of Hematopoietic Stem Cells as a Therapy for HIV/AIDS. Viruses. 2013; 5(12):2946-2962. https://doi.org/10.3390/v5122946
Chicago/Turabian StyleYounan, Patrick, John Kowalski, and Hans-Peter Kiem. 2013. "Genetic Modification of Hematopoietic Stem Cells as a Therapy for HIV/AIDS" Viruses 5, no. 12: 2946-2962. https://doi.org/10.3390/v5122946