Varicella Zoster Virus (VZV)-Human Neuron Interaction

Abstract
:1. Introduction
2. Latent VZV in Human Ganglia
3. Explanted Human Ganglia
4. Human Neurons in SCID Mice
5. Human Neuroblastoma Cells
6. Human Stem Cells
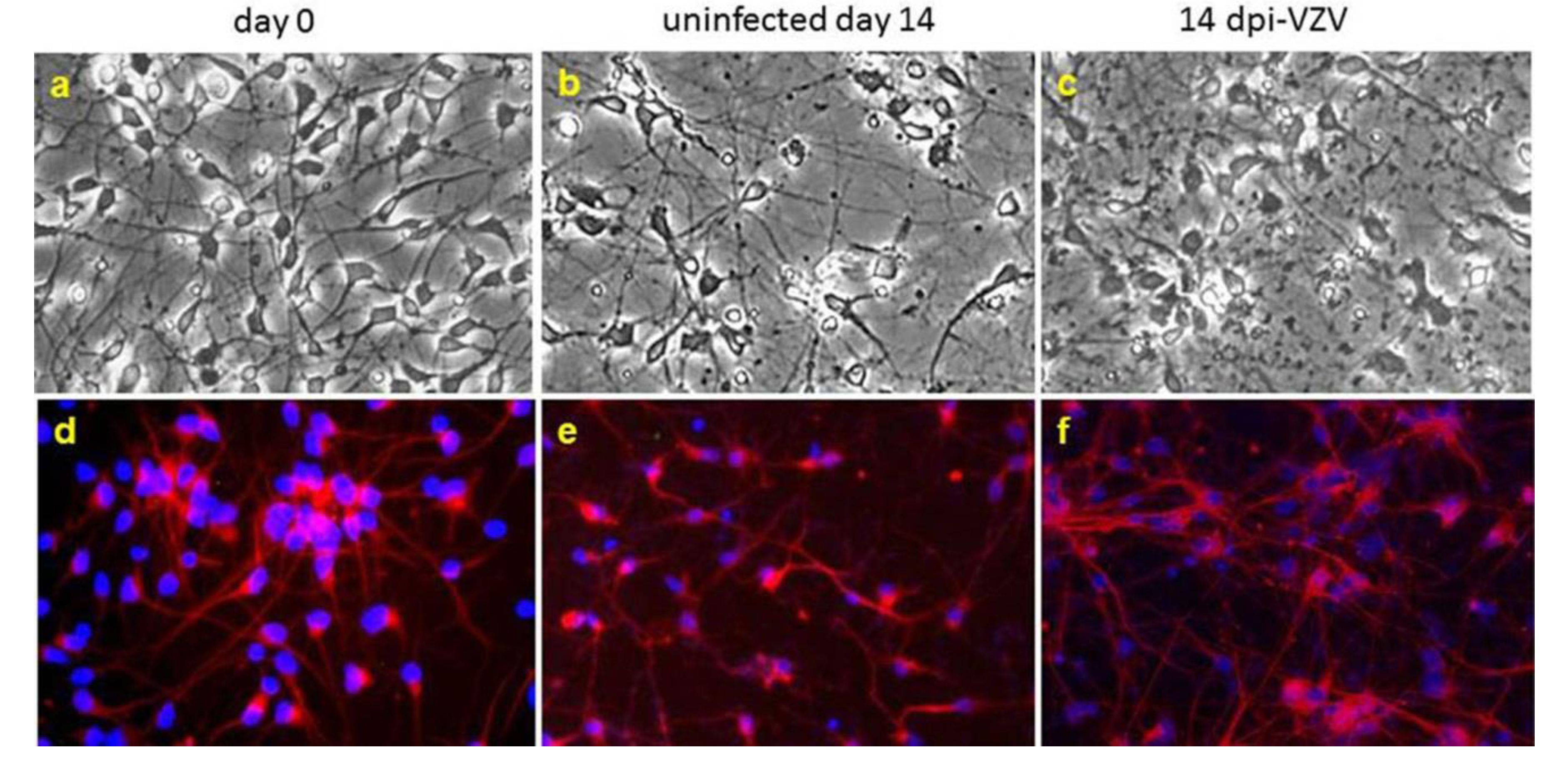
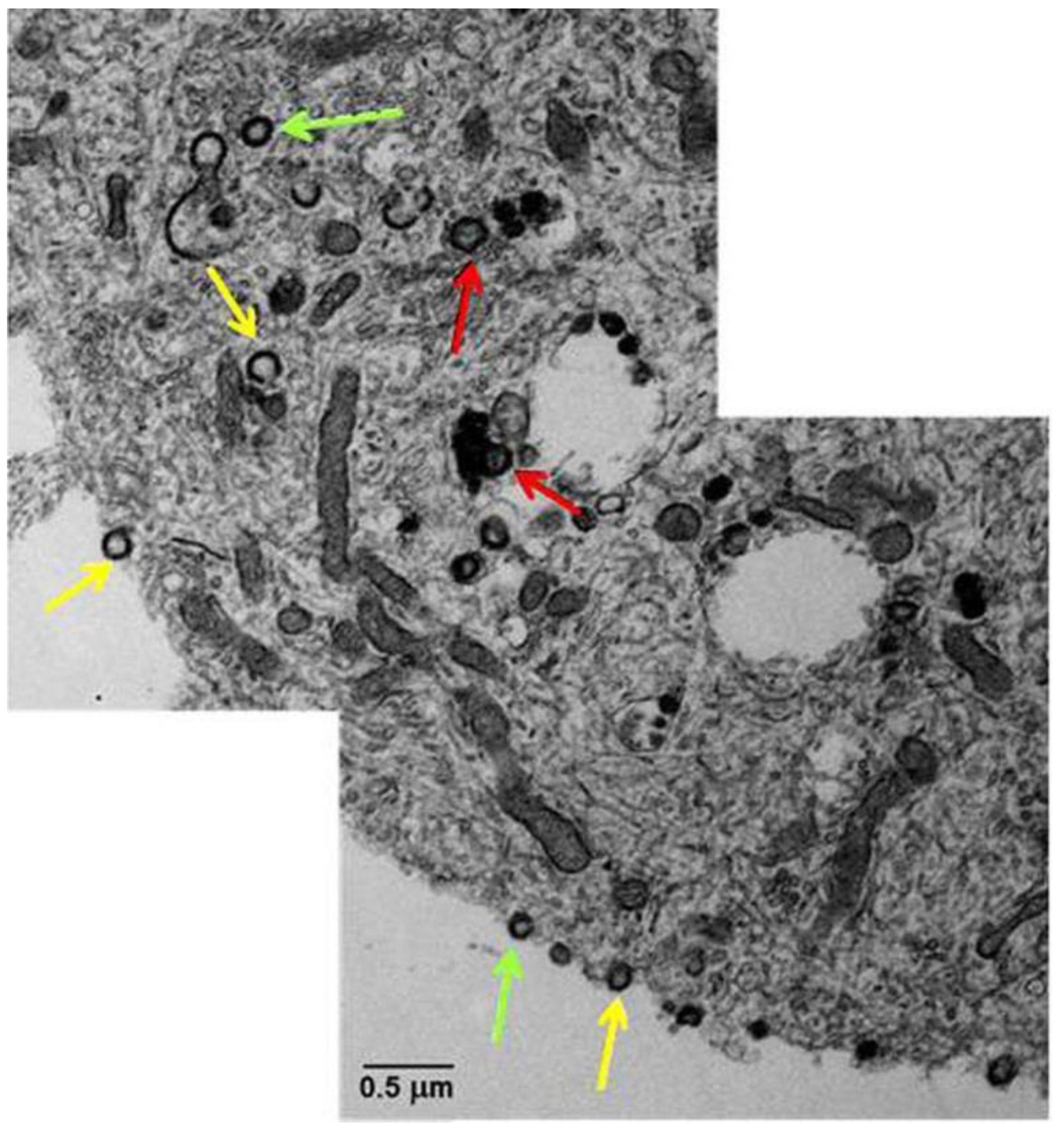
7. Induced Human Pluripotent Stem Cells (iPSC)



8. Normal Human Neural Progenitor (NHNP) Tissue-Like Assemblies (TLA)
9. Conclusions

{kind=link}
{kind=link}
{kind=link}
| Model | Virus inoculum | Results | Reference |
|---|---|---|---|
| fetal neural stem cells implanted into SCID mouse brain | cell-associated | 3 weeks p.i., VZV proteins encoded by ORFs 62, 63 & 47 are detected; VZV gE is rarely detected. | [23] |
| fetal DRG implanted under SCID mouse kidney capsule | cell-associated | 8 weeks p.i., no infectious virus is released; VZV DNA copy number is stable with limited VZV transcription (62/63 only). | [24] |
| embryonic neural stem cell (hNSC) | cell-free | 14 days p.i., no CPE or release of infectious virus, although VZV DNA, RNA, and protein are detectable. | [30] |
| induced pluripotent stem cell | cell-free | 14 days p.i., no CPE or release of infectious virus, although VZV DNA, RNA, & proteins are detected; virions seen - mostly aberrant; no markers of activated apoptotic pathway present. | [33,34] |
| human neural progenitor cells tissue like assemblies | cell-free | first 18 days p.i. show an increase of both VZV DNA and RNA, which plateaus. At 3 months p.i., no CPE is evident and only sporadic release of infectious virus is seen. | [35] |
Acknowledgments
Conflicts of Interest
References
- Gilden, D.H.; Cohrs, R.J.; Mahalingam, R. Clinical and molecular pathogenesis of varicella virus infection. Viral Immunol. 2003, 16, 243–258. [Google Scholar] [CrossRef]
- Gilden, D.H.; Gesser, R.; Smith, J.; Wellish, M.; Laguardia, J.J.; Cohrs, R.J.; Mahalingam, R. Presence of VZV and HSV-1 DNA in human nodose and celiac ganglia. Virus Genes 2001, 23, 145–147. [Google Scholar] [CrossRef]
- Gilden, D.H.; Vafai, A.; Shtram, Y.; Becker, Y.; Devlin, M.; Wellish, M. Varicella-zoster virus DNA in human sensory ganglia. Nature 1983, 306, 478–480. [Google Scholar] [CrossRef]
- Hyman, R.W.; Ecker, J.R.; Tenser, R.B. Varicella-zoster virus RNA in human trigeminal ganglia. Lancet 1983, 2, 814–816. [Google Scholar] [CrossRef]
- Azarkh, Y.; Gilden, D.; Cohrs, R.J. Molecular characterization of varicella zoster virus in latently infected human ganglia: Physical state and abundance of VZV DNA, quantitation of viral transcripts and detection of VZV-specific proteins. Curr. Top. Microbiol. Immunol. 2010, 342, 229–241. [Google Scholar] [CrossRef]
- Gilden, D.; Mahalingam, R.; Nagel, M.A.; Pugazhenthi, S.; Cohrs, R.J. Review: The neurobiology of varicella zoster virus infection. Neuropathol. Appl. Neurobiol. 2011, 37, 441–463. [Google Scholar] [CrossRef]
- Dueland, A.N.; Martin, J.R.; Devlin, M.E.; Wellish, M.; Mahalingam, R.; Cohrs, R.; Soike, K.F.; Gilden, D.H. Acute simian varicella infection. Clinical, laboratory, pathologic, and virologic features. Lab. Investig. 1992, 66, 762–773. [Google Scholar]
- Clarke, P.; Beer, T.; Cohrs, R.; Gilden, D.H. Configuration of latent varicella-zoster virus DNA. J. Virol. 1995, 69, 8151–8154. [Google Scholar]
- Cohrs, R.J.; Randall, J.; Smith, J.; Gilden, D.H.; Dabrowski, C.; van Der Keyl, H.; Tal-Singer, R. Analysis of individual human trigeminal ganglia for latent herpes simplex virus type 1 and varicella-zoster virus nucleic acids using real-time PCR. J. Virol. 2000, 74, 11464–11471. [Google Scholar] [CrossRef]
- Mahalingam, R.; Wellish, M.; Lederer, D.; Forghani, B.; Cohrs, R.; Gilden, D. Quantitation of latent varicella-zoster virus DNA in human trigeminal ganglia by polymerase chain reaction. J. Virol. 1993, 67, 2381–2384. [Google Scholar]
- Pevenstein, S.R.; Williams, R.K.; McChesney, D.; Mont, E.K.; Smialek, J.E.; Straus, S.E. Quantitation of latent varicella-zoster virus and herpes simplex virus genomes in human trigeminal ganglia. J. Virol. 1999, 73, 10514–10518. [Google Scholar]
- Nagel, M.A.; Choe, A.; Traktinskiy, I.; Cordery-Cotter, R.; Gilden, D.; Cohrs, R.J. Varicella-zoster virus transcriptome in latently infected human ganglia. J. Virol. 2011, 85, 2276–2287. [Google Scholar] [CrossRef]
- Nagel, M.A.; Gilden, D.; Shade, T.; Gao, B.; Cohrs, R.J. Rapid and sensitive detection of 68 unique varicella zoster virus gene transcripts in five multiplex reverse transcription-polymerase chain reactions. J. Virol. Methods 2009, 157, 62–68. [Google Scholar] [CrossRef]
- Cohrs, R.J.; Gilden, D.H. Prevalence and abundance of latently transcribed varicella-zoster virus genes in human ganglia. J. Virol. 2007, 81, 2950–2956. [Google Scholar] [CrossRef]
- Ouwendijk, W.J.; Choe, A.; Nagel, M.A.; Gilden, D.; Osterhaus, A.D.; Cohrs, R.J.; Verjans, G.M. Restricted varicella-zoster virus transcription in human trigeminal ganglia obtained soon after death. J. Virol. 2012, 86, 10203–10206. [Google Scholar] [CrossRef]
- Gary, L.; Gilden, D.H.; Cohrs, R.J. Epigenetic regulation of varicella-zoster virus open reading frames 62 and 63 in latently infected human trigeminal ganglia. J. Virol. 2006, 80, 4921–4926. [Google Scholar] [CrossRef]
- Zerboni, L.; Sobel, R.A.; Lai, M.; Triglia, R.; Steain, M.; Abendroth, A.; Arvin, A. Apparent expression of varicella-zoster virus proteins in latency resulting from reactivity of murine and rabbit antibodies with human blood group A determinants in sensory neurons. J. Virol. 2012, 86, 578–583. [Google Scholar] [CrossRef]
- Gilden, D.H.; Wroblewska, Z.; Kindt, V.; Warren, K.G.; Wolinsky, J.S. Varicella-zoster virus infection of human brain cells and ganglion cells in tissue culture. Arch. Virol. 1978, 56, 105–117. [Google Scholar] [CrossRef]
- Wigdahl, B.; Rong, B.L.; Kinney-Thomas, E. Varicella-zoster virus infection of human sensory neurons. Virology 1986, 152, 384–399. [Google Scholar] [CrossRef]
- Hood, C.; Cunningham, A.L.; Slobedman, B.; Boadle, R.A.; Abendroth, A. Varicella-zoster virus-infected human sensory neurons are resistant to apoptosis, yet human foreskin fibroblasts are susceptible: Evidence for a cell-type-specific apoptotic response. J. Virol. 2003, 77, 12852–12864. [Google Scholar] [CrossRef]
- Gowrishankar, K.; Slobedman, B.; Cunningham, A.L.; Miranda-Saksena, M.; Boadle, R.A.; Abendroth, A. Productive varicella-zoster virus infection of cultured intact human ganglia. J. Virol. 2007, 81, 6752–6756. [Google Scholar] [CrossRef]
- Azarkh, Y.; Bos, N.; Gilden, D.; Cohrs, R.J. Human trigeminal ganglionic explants as a model to study alphaherpesvirus reactivation. J. Neurovirol. 2012, 18, 456–461. [Google Scholar] [CrossRef]
- Baiker, A.; Fabel, K.; Cozzio, A.; Zerboni, L.; Fabel, K.; Sommer, M.; Uchida, N.; He, D.; Weissman, I.; Arvin, A.M. Varicella-zoster virus infection of human neural cells in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 10792–10797. [Google Scholar] [CrossRef]
- Zerboni, L.; Ku, C.C.; Jones, C.D.; Zehnder, J.L.; Arvin, A.M. Varicella-zoster virus infection of human dorsal root ganglia in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 6490–6495. [Google Scholar]
- Christensen, J.; Steain, M.; Slobedman, B.; Abendroth, A. Differentiated neuroblastoma cells provide a highly efficient model for studies of productive varicella-zoster virus infection of neuronal cells. J. Virol. 2011, 85, 8436–8442. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Markus, A.; Grigoryan, S.; Sloutskin, A.; Yee, M.B.; Zhu, H.; Yang, I.H.; Thakor, N.V.; Sarid, R.; Kinchington, P.R.; Goldstein, R.S. Varicella-zoster virus (VZV) infection of neurons derived from human embryonic stem cells: Direct demonstration of axonal infection, transport of VZV, and productive neuronal infection. J. Virol. 2011, 85, 6220–6233. [Google Scholar] [CrossRef]
- Dukhovny, A.; Sloutskin, A.; Markus, A.; Yee, M.B.; Kinchington, P.R.; Goldstein, R.S. Varicella-zoster virus infects human embryonic stem cell-derived neurons and neurospheres but not pluripotent embryonic stem cells or early progenitors. J. Virol. 2012, 86, 3211–3218. [Google Scholar] [CrossRef]
- Grigoryan, S.; Kinchington, P.R.; Yang, I.H.; Selariu, A.; Zhu, H.; Yee, M.; Goldstein, R.S. Retrograde axonal transport of VZV: Kinetic studies in hESC-derived neurons. J. Neurovirol. 2012, 18, 462–470. [Google Scholar] [CrossRef]
- Pugazhenthi, S.; Nair, S.; Velmurugan, K.; Liang, Q.; Mahalingam, R.; Cohrs, R.J.; Nagel, M.A.; Gilden, D. Varicella-zoster virus infection of differentiated human neural stem cells. J. Virol. 2011, 85, 6678–6686. [Google Scholar] [CrossRef]
- Sloutskin, A.; Kinchington, P.R.; Goldstein, R.S. Productive vs. non-productive infection by cell-free varicella zoster virus of human neurons derived from embryonic stem cells is dependent upon infectious viral dose. Virology 2013, 443, 285–293. [Google Scholar] [CrossRef]
- Lee, K.S.; Zhou, W.; Scott-McKean, J.J.; Emmerling, K.L.; Cai, G.Y.; Krah, D.L.; Costa, A.C.; Freed, C.R.; Levin, M.J. Human sensory neurons derived from induced pluripotent stem cells support varicella-zoster virus infection. PLoS One 2012, 7, e53010. [Google Scholar]
- Yu, X.; Seitz, S.; Pointon, T.; Bowlin, J.L.; Cohrs, R.J.; Jonjic, S.; Haas, J.; Wellish, M.; Gilden, D. Varicella zoster virus infection of highly pure terminally differentiated human neurons. J. Neurovirol. 2013, 19, 75–81. [Google Scholar] [CrossRef]
- Grose, C.; Yu, X.; Cohrs, R.J.; Carpenter, J.E.; Jacqueline, J.; Gilden, D. Aberrant virion assembly and limited gC production in varicella zoster virus-infected neurons. J. Virol. 2013, 87, 9643–9648. [Google Scholar] [CrossRef]
- Goodwin, T.J.; McCarthy, M.; Osterrieder, N.; Cohrs, R.J.; Kaufer, B.B. Three-dimensional normal human neural progenitor tissure-like assemblies: A model of persisitant varicella-zoster virus infection. PLoS Pathog. 2013, 8, e1003512. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Baird, N.L.; Yu, X.; Cohrs, R.J.; Gilden, D. Varicella Zoster Virus (VZV)-Human Neuron Interaction. Viruses 2013, 5, 2106-2115. https://doi.org/10.3390/v5092106
Baird NL, Yu X, Cohrs RJ, Gilden D. Varicella Zoster Virus (VZV)-Human Neuron Interaction. Viruses. 2013; 5(9):2106-2115. https://doi.org/10.3390/v5092106
Chicago/Turabian StyleBaird, Nicholas L., Xiaoli Yu, Randall J. Cohrs, and Don Gilden. 2013. "Varicella Zoster Virus (VZV)-Human Neuron Interaction" Viruses 5, no. 9: 2106-2115. https://doi.org/10.3390/v5092106