2.1. The HCMV IE1 Gene Disrupts Signaling by Type II Interferon
To identify the HCMV genes involved in disruption of IFN signaling a cDNA library of the HCMV laboratory strain AD169 was constructed [
33]. The human fibrosarcoma cell line 2C4 was used in a preliminary screen for HCMV cDNA clones that have a role in regulating IFN signaling. 2C4 is a fibrosarcoma cell line engineered to express the T-cell antigen CD2 under the control of the promoter element of the Interferon Induced Transmembrane protein 1 (
IFITM1) gene (a.k.a. 9-27, IFI17, CD225) which respond to both type I and II IFNs by increasing cell surface expression of CD2 [
34]. 2C4 cells transfected with any HCMV cDNA clone involved in disruption of IFN signaling would thus be expected to exhibit reduced accumulation of cell surface of CD2. Using this system we observed that transfection of cDNA clone pIE622, harboring the full length cDNA sequence for the HCMV
UL123 gene, was associated with diminished CD2 cell surface levels relative to empty vector-transfected cells after exposure to IFNβ, and to a lesser extent, IFNγ (data not shown). This suggested a role for IE1 in disruption of both type I and type II IFN signaling. The
UL123 gene codes for the HCMV major transcriptional transactivator protein, IE1. In addition to its role as a promiscuous transactivator of viral and cellular genes, IE1 is known to have multiple functions including disruption of ND10 nuclear bodies [
35], antagonism of histone deaceytlase (HDAC3) [
36], anti-apoptotic function [
37], chromatin tethering [
38], and interference with signaling by type I interferons [
21].
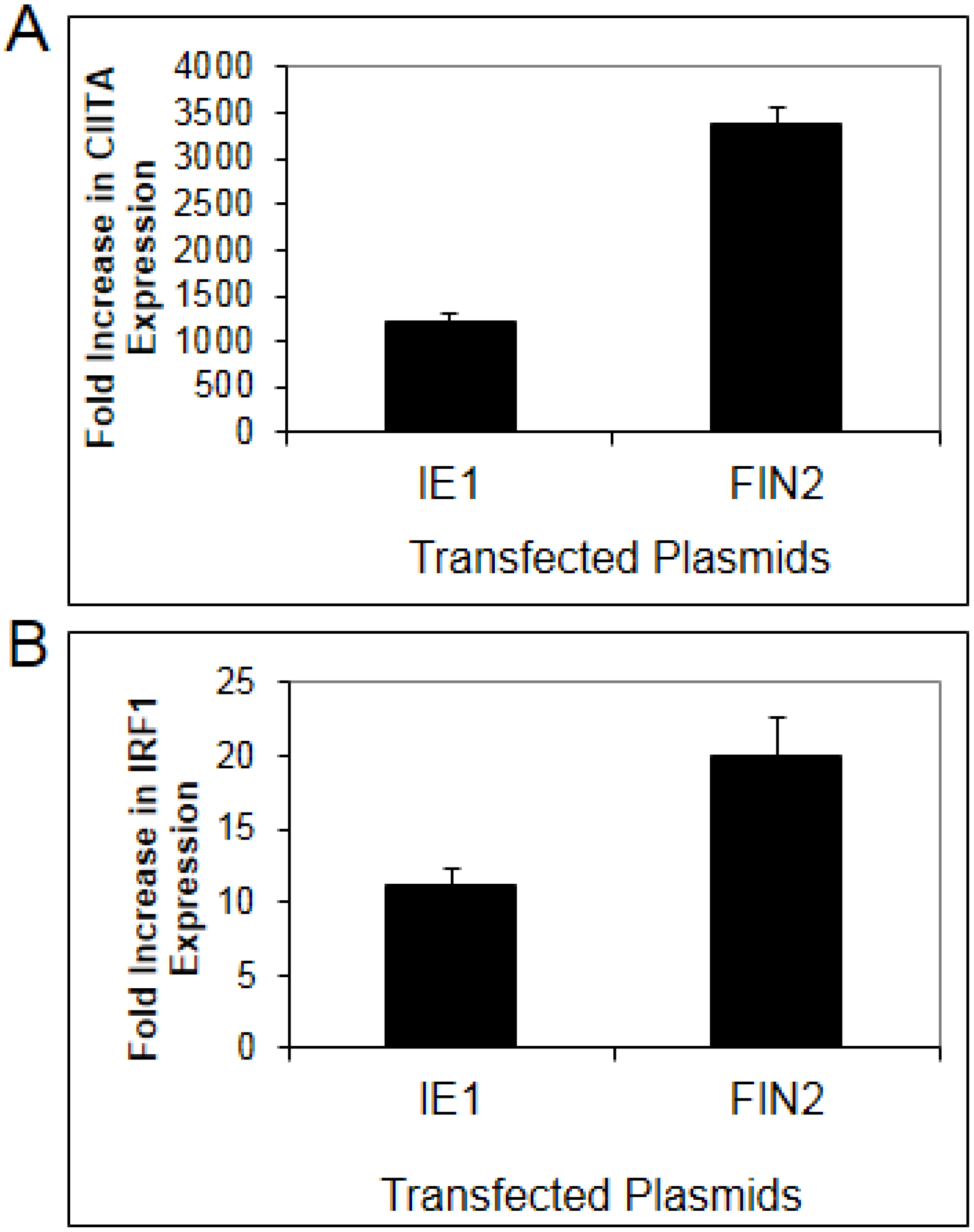
To confirm a role for IE1 in disruption of type II IFN signaling, we examined the effect of IE1 in a more physiologically relevant human fibroblast cell line. Basal expression of MHC II Transcriptional Activator (CIITA) is very low in fibroblasts but becomes highly induced after IFNγ treatment. CIITA transcript levels were quantified by Real-Time RT-PCR in MRC5 fibroblasts after nucleofection with pIE622 expressing the full length IE1. In MRC5 cells, nucleofection resulted in greater than 90% transfection efficiency (data not shown). CIITA mRNA levels were compared in IFNγ treated cells and untreated cells. In untreated cells, expression of IE1 was associated with a slight induction of CIITA or IRF1 (0.9 to 3 fold). In IFNγ treated cells, we found that CIITA induction in IE1 expressing cells was only 39% of that observed in cells nucleofected with empty vector (
Figure 1A). We also examined the IFNγ-induced expression of a second gene, IRF1, in IE1 expressing fibroblasts. We found that the increase in expression of IRF1 in response to IFNγ is reduced by 52% in IE1 expressing cells as compared to cells nucleofected with empty vector (
Figure 1B) relative to untreated cells. These findings confirm that IE1 is able to impair, but not completely block IFNγ-induced upregulation of CIITA and IRF1 transcript levels in human fibroblasts.
2.2. Mapping of the IE1 Protein Region Involved in Disruption of IFN-Induced Gene Expression
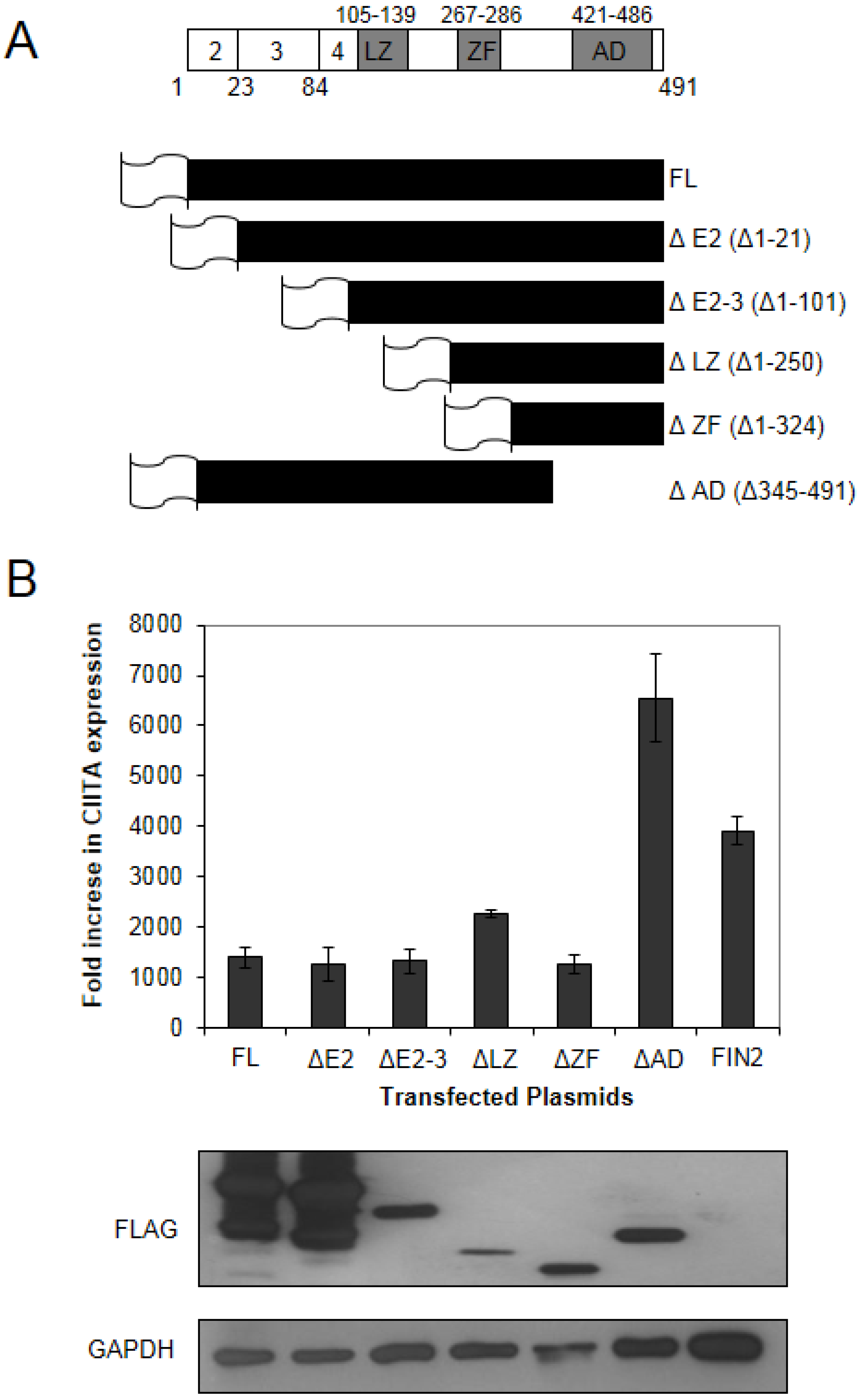
The HCMV IE1 protein is a multifunctional protein with various functions attributed to different domains of the protein (
Figure 2A). We generated a series of FLAG epitope-tagged truncation mutants to identify the protein region required for attenuation of IFNγ-induced gene expression as described in Materials and Methods. We generated five different FLAG-tagged truncation mutants plus a full length FLAG-tagged version of IE1 (
Figure 2A). These six different clones were nucleofected into MRC5 cells, followed by treatment with IFNγ for 6 hours. RNA extracted from these cells was analyzed for levels of CIITA transcript induction by Real-Time RT-PCR. We found that there was a reduction in the IFNγ-induced expression of CIITA in cells expressing full length and truncated versions of IE1, with the exception of the C-terminal truncation missing residues 345–491 (ΔAD) that includes the acidic domain and chromatin tethering domain (
Figure 2B). We consistently observed higher levels of CIITA induction in cells expressing the Δ345–491 protein relative to our vector alone transfections. Although the reason for this is not clear, it may be that the IE1 protein without the C-terminal domain provides a signal that amplifies IFNγ-induced CIITA gene expression. This may also relate to the report linking inducible IE1 expression to STAT1-dependent interferon-like gene induction [
31]. One possibility is that IE1 may interact with cellular proteins that stimulate a type II interferon response, but this activity is masked by the dominant inhibitory role of the C-terminal 147 residues in our studies. These contradictory data may also reflect the different transfection systems or viral strains used in these two studies.
Figure 1.
Diminished interferon-induced gene expression in human fibroblasts expressing IE1. MRC-5 fibroblasts were nucleofected with plasmid pIE622 harboring the UL123 gene specifying IE1, or the pFIN2 empty vector. At 24 hours post-nucleofection, the cells were exposed to 100 U/mL of IFNγ or were left untreated. At 6 hours after treatment, total RNA was isolated and subjected to Real Time RT-PCR analysis as described in Materials and Methods. The fold increase in CIITA (A) (p value = 0.053) or IRF1 (B) (p value 0.017) transcript levels in IFN treated cells relative to untreated cells was determined by the ΔΔCt method. Data shown are the average of three independent experiments.
Figure 1.
Diminished interferon-induced gene expression in human fibroblasts expressing IE1. MRC-5 fibroblasts were nucleofected with plasmid pIE622 harboring the UL123 gene specifying IE1, or the pFIN2 empty vector. At 24 hours post-nucleofection, the cells were exposed to 100 U/mL of IFNγ or were left untreated. At 6 hours after treatment, total RNA was isolated and subjected to Real Time RT-PCR analysis as described in Materials and Methods. The fold increase in CIITA (A) (p value = 0.053) or IRF1 (B) (p value 0.017) transcript levels in IFN treated cells relative to untreated cells was determined by the ΔΔCt method. Data shown are the average of three independent experiments.
Figure 2.
The carboxy-terminal region of IE1 is required for disruption of IFNγ signaling. (A) Schematic of the IE1 protein and truncation mutants generated in this study. Top line: a schematic of the IE1 protein indicating the amino acid positions comprising the coding exons (below) and the positions of the known functional domains (above). The coding exons are shown in white boxes. The functional domains of exon 4 are shown in shaded boxes and include the leucine zipper (L), the zinc finger (ZF) and the acidic domain (AD). A schematic of the five different IE1 N-terminal truncation mutants generated in this study are shown below. The sixth mutant is a C-terminal truncation mutant. The designation of the plasmids harboring the truncation mutants is based on the deletion of relevant exons or functional domains shown on the right, and the deleted amino acid residues are shown in parentheses. The gene sequences were inserted in frame to sequences specifying a FLAG epitope (depicted as a flag). (B) Interferon Stimulated Gene (ISG) induction in fibroblasts expressing truncated IE1 proteins. The indicated plasmids were nucleoporated into MRC5 cells and 24 hours after nucleofection the cells were treated with 100 U/mL IFNγ. At 6 hours after treatment total RNA was isolated. Real Time RT-PCR analysis was carried out for CIITA and 18S rRNA (endogenous control). The fold increase in CIITA transcript levels in IFN treated cells relative to untreated cells was determined by the ΔΔCt method. Shown is the average of two independent experiments. A film image of immunoblot analysis is shown below. In replicate cultures of the first experiment, cells were treated as above and solubilized. Equal amounts of protein from each lysate were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies. Anti-FLAG antibody was used to detect the presence of FLAG tagged IE1 proteins and antibody to GAPDH was used to evaluate protein loading.
Figure 2.
The carboxy-terminal region of IE1 is required for disruption of IFNγ signaling. (A) Schematic of the IE1 protein and truncation mutants generated in this study. Top line: a schematic of the IE1 protein indicating the amino acid positions comprising the coding exons (below) and the positions of the known functional domains (above). The coding exons are shown in white boxes. The functional domains of exon 4 are shown in shaded boxes and include the leucine zipper (L), the zinc finger (ZF) and the acidic domain (AD). A schematic of the five different IE1 N-terminal truncation mutants generated in this study are shown below. The sixth mutant is a C-terminal truncation mutant. The designation of the plasmids harboring the truncation mutants is based on the deletion of relevant exons or functional domains shown on the right, and the deleted amino acid residues are shown in parentheses. The gene sequences were inserted in frame to sequences specifying a FLAG epitope (depicted as a flag). (B) Interferon Stimulated Gene (ISG) induction in fibroblasts expressing truncated IE1 proteins. The indicated plasmids were nucleoporated into MRC5 cells and 24 hours after nucleofection the cells were treated with 100 U/mL IFNγ. At 6 hours after treatment total RNA was isolated. Real Time RT-PCR analysis was carried out for CIITA and 18S rRNA (endogenous control). The fold increase in CIITA transcript levels in IFN treated cells relative to untreated cells was determined by the ΔΔCt method. Shown is the average of two independent experiments. A film image of immunoblot analysis is shown below. In replicate cultures of the first experiment, cells were treated as above and solubilized. Equal amounts of protein from each lysate were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies. Anti-FLAG antibody was used to detect the presence of FLAG tagged IE1 proteins and antibody to GAPDH was used to evaluate protein loading.
![Viruses 06 01502 g002]()
2.3. Disruption of IFNγ Signaling by IE1 Does Not Involve Interaction with PML
Promyelocytic leukemia protein (PML) is involved in transcriptional repression and is a master organizer of nuclear domain (ND)10 structures. Since IE1 has been reported to be involved in the disruption of ND10 structures in the nucleus [
35,
39,
40,
41,
42,
43] and because PML is known to mediate antiviral activities of IFNs in HSV-infected cells [
44] and to confer intrinsic immunity against CMV [
45,
46], we set out to test the hypothesis that the function of IE1 in disrupting IFNγ-induced gene expression is linked to its role in disrupting ND10 structures by immunofluorescence microscopy. We found that in fibroblasts expressing the full length IE1 and the Δ345–491 C-terminal deletion mutant of IE1, PML was dispersed throughout the nucleus, indicating disruption of ND10s (
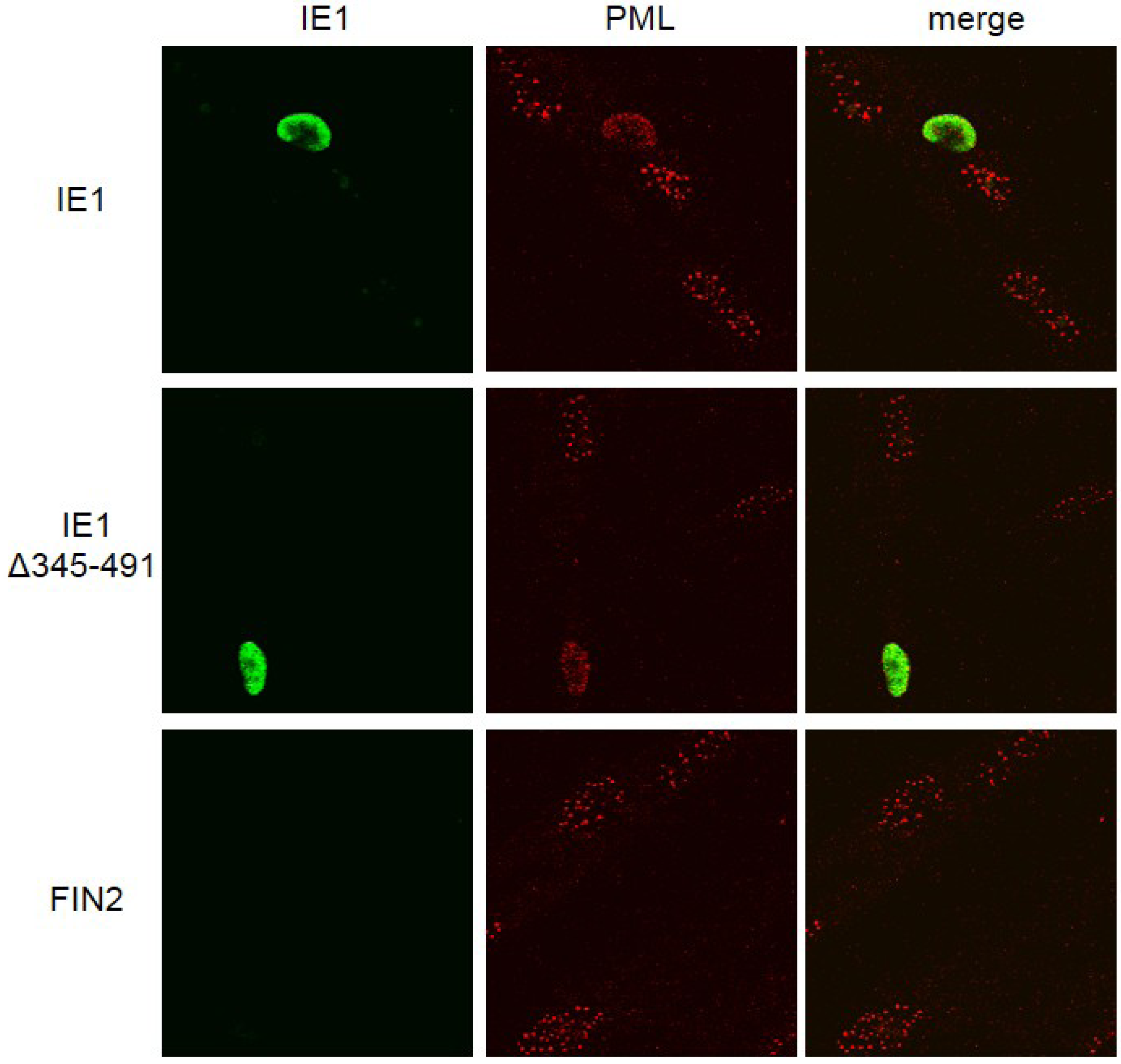
Figure 3). As expected, PML staining was in the form of punctate dots in the nucleus in the absence of IE1. Therefore, the C-terminal deletion mutant retains the ability to disrupt ND10s. We conclude that IE1‑mediated disruption of the IFNγ signaling pathway is not strictly linked to IE1-mediated dispersal of ND10s.
Figure 3.
Disruption of ND10 structures in cells expressing IE1 and the C-terminal truncation mutant. Immunofluorescence images of IE1 and Promyelocytic leukemia protein (PML) in fibroblast cells. MRC5 cells were nucleofected with the indicated constructs. At 24 hours after nucleofection cells were fixed in methanol and reacted with mouse monoclonal anti-IE1 antibody (Mab810) followed by anti-mouse alexafluor 488 as secondary antibody. The cells were then exposed to rhodamine conjugated anti PML antibody (PGM3). IE1 reactivity is visualized in green (left column), PML reactivity is visualized in red (middle column) and merged images are shown in the right column.
Figure 3.
Disruption of ND10 structures in cells expressing IE1 and the C-terminal truncation mutant. Immunofluorescence images of IE1 and Promyelocytic leukemia protein (PML) in fibroblast cells. MRC5 cells were nucleofected with the indicated constructs. At 24 hours after nucleofection cells were fixed in methanol and reacted with mouse monoclonal anti-IE1 antibody (Mab810) followed by anti-mouse alexafluor 488 as secondary antibody. The cells were then exposed to rhodamine conjugated anti PML antibody (PGM3). IE1 reactivity is visualized in green (left column), PML reactivity is visualized in red (middle column) and merged images are shown in the right column.
Earlier there have been conflicting reports, one suggesting that the acidic domain is important for PML targeting and disruption of ND10 structures [
41] and the other suggesting that the acidic domain is not involved in this function of IE1 [
39]. Yet another study revealed an intermediate role for the acidic domain IE1 wherein it was required to target ND10 structures but could not disrupt them [
47]. Our results indicate that a C-terminal truncation mutant that includes the entire acidic domain is able to disrupt ND10s similar to the full length IE1 protein in MRC5 cells. Our results are in agreement the findings of Wilkinson
et al. [
39]. Although these results to do not preclude a role for PML in IE1‑mediated IFN signal disruption, they do demonstrate that this function of IE1 does not involve disruption of ND10 structures.
2.5. Type II Interferon-Induced Binding of STAT1 to GAS Elements Is Reduced in the Presence of IE1
We next determined whether STAT1 molecules in the nucleus were competent to bind the GAS element derived from the promoter of the IRF1 gene by EMSA. The full length IE1, the Δ345–491 mutant (ΔAD), and empty vector (FIN2) were nucleofected into MRC5 cells. These cells were either treated with IFNγ for 30 minutes or left untreated. Nuclear extracts from these cells were incubated with a radiolabeled 22mer IRF1 GAS element probe. The amount of probe shifted in nuclear extracts from IE1-expressing cells is diminished relative to extracts derived from empty vector- or Δ345–491-nucleofected cells (
Figure 6A, compare lane 6 to lanes 5 and 7). These bands could be supershifted using a STAT1 antibody indicating that the shift was caused by a complex containing STAT1 (lanes 11–13). This demonstrates that there is reduced binding of STAT1 to GAS elements in the presence of IE1. Studies were also conducted with nuclear extracts from HCMV-infected cells at 12 hours after infection. We could detect only a minor STAT1-shifted probe band in HCMV infected cells compared to uninfected cells (
Figure 6B, lane 2), which may be related to the much higher levels of IE1 in infected cells relative to cells nucleofected with full length IE1 (data not shown). Together, these data indicate that expression of HCMV IE1 is sufficient to impair STAT1 binding to GAS elements, and that this activity requires the C-terminal region of IE1 including residues 344 to 491. These data also suggest that the impairment of functional STAT1 dimer binding to GAS elements at 12 hours after infection is a result of IE1 expression.
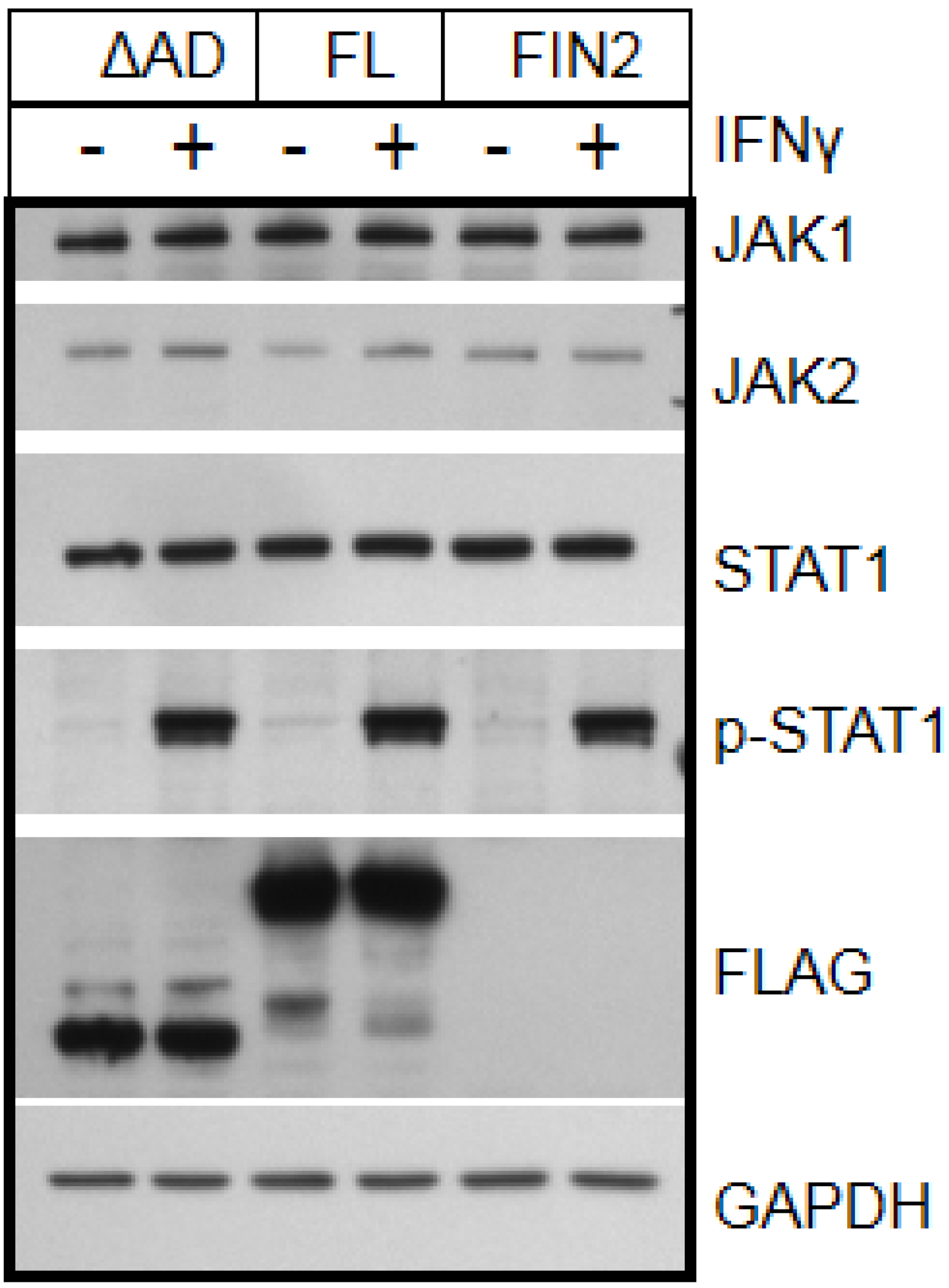
Figure 4.
IE1 does not alter the state or abundance of key signaling molecules of the IFNγ pathway. Film images of electrophoretically separated cell lysates reacted with antibodies to IFNγ pathway signaling components. MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 24 hours after nucleofection, cells were exposed to IFNγ for 30 minutes or left untreated. Cells were solublized and equal amounts of protein from each lysate were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies. Anti-FLAG antibody was used to detect the presence of FLAG tagged IE1 proteins and antibody to GAPDH was used to evaluate protein loading.
Figure 4.
IE1 does not alter the state or abundance of key signaling molecules of the IFNγ pathway. Film images of electrophoretically separated cell lysates reacted with antibodies to IFNγ pathway signaling components. MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 24 hours after nucleofection, cells were exposed to IFNγ for 30 minutes or left untreated. Cells were solublized and equal amounts of protein from each lysate were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies. Anti-FLAG antibody was used to detect the presence of FLAG tagged IE1 proteins and antibody to GAPDH was used to evaluate protein loading.
IE1 has been reported to antagonize HDAC activity [
36]. Although HDAC activity is typically associated with transcriptional repression, it is associated with transcriptional activation of ISGs [
48]. It is tempting to speculate that antagonism of HDAC activity by IE1 is linked to disruption of interferon signaling. Further implicating a role for chromatin organization is that the C-terminal region of IE1 includes the region previously shown to be required for chromatin tethering [
38]. However, this possibility is difficult to reconcile with the observation that IE1 disrupts STAT1-binding to a GAS element probe used in our EMSA assays, which is not dependent on chromatin structure. We therefore propose that a novel function of IE1 impairs STAT1 binding to target GAS elements.
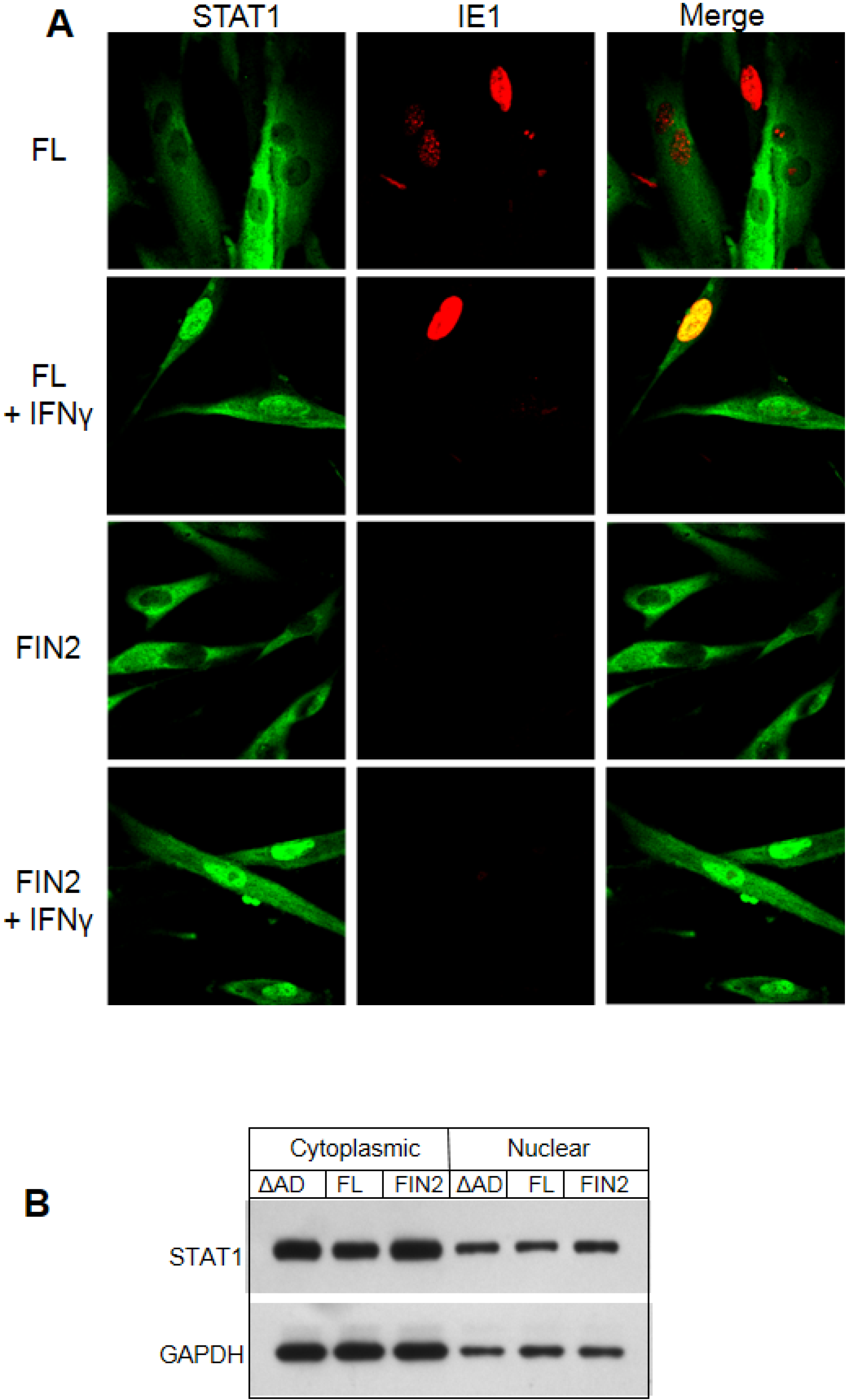
Figure 5.
Nuclear translocation of STAT1 is not altered by IE1. (A) Immunofluorescence images of IE1 and STAT1 in cells with and without IFNγ treatment. MRC5 cells were nucleofected with the full length IE1 (FL) or empty vector (FIN2). After 24 hours cells were exposed to IFNγ for 30 minutes or left untreated. Cells were fixed in methanol and reacted with mouse monoclonal anti-IE1 antibody (Mab810) and rabbit polyclonal anti‑STAT1 antibody followed by anti-mouse alexafluor 543 and anti-rabbit alexafluor 488 as secondary antibodies. STAT1 reactivity is visualized in green (left column), IE1 reactivity is visualized in red (middle column) and merged images are shown in the right column. (B) Film images of electrophoretically separated cell lysates reacted with antibodies to STAT1 and GAPDH (loading control). MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 72 hours after nucleofection, cells were exposed to IFNγ for 30 minutes or left untreated. Cells were harvested and nuclear and cytoplasmic fractions were separated. Equal amounts of protein from the nuclear and cytoplasmic lysates were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies.
Figure 5.
Nuclear translocation of STAT1 is not altered by IE1. (A) Immunofluorescence images of IE1 and STAT1 in cells with and without IFNγ treatment. MRC5 cells were nucleofected with the full length IE1 (FL) or empty vector (FIN2). After 24 hours cells were exposed to IFNγ for 30 minutes or left untreated. Cells were fixed in methanol and reacted with mouse monoclonal anti-IE1 antibody (Mab810) and rabbit polyclonal anti‑STAT1 antibody followed by anti-mouse alexafluor 543 and anti-rabbit alexafluor 488 as secondary antibodies. STAT1 reactivity is visualized in green (left column), IE1 reactivity is visualized in red (middle column) and merged images are shown in the right column. (B) Film images of electrophoretically separated cell lysates reacted with antibodies to STAT1 and GAPDH (loading control). MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 72 hours after nucleofection, cells were exposed to IFNγ for 30 minutes or left untreated. Cells were harvested and nuclear and cytoplasmic fractions were separated. Equal amounts of protein from the nuclear and cytoplasmic lysates were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies.
![Viruses 06 01502 g005]()
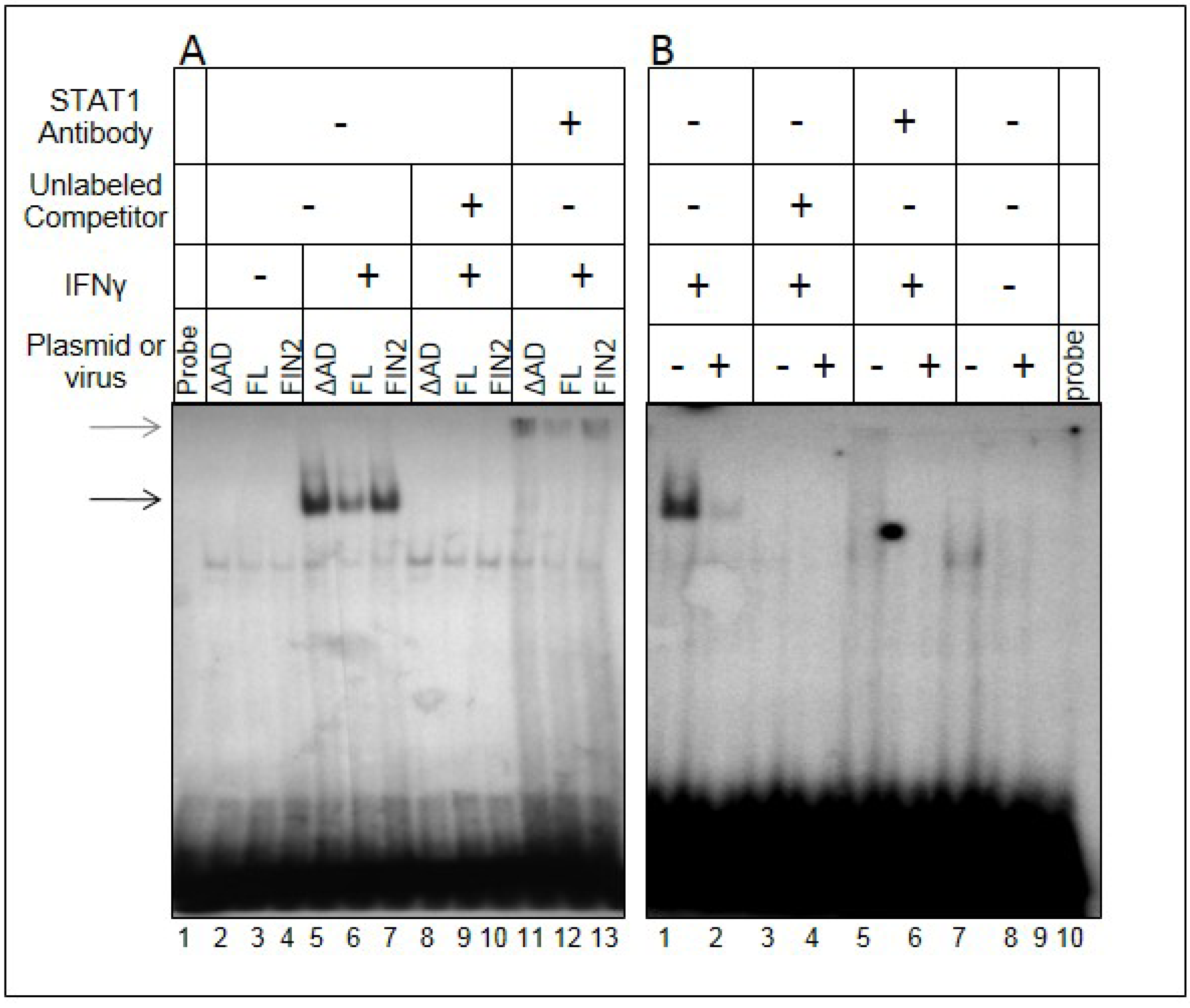
Figure 6.
Reduced binding of STAT1 to GAS elements in the presence of IE1. Film image of an Electrophoretic Mobility Shift Assay shows a reduction in the amount of IRF1 GAS element probe that is shifted upon mixture with nuclear extracts from MRC5 cells ectopically expressing IE1 (A) and HCMV infected cells (B). In A, MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 24 hours after nucleofection, cells were exposed to 100 U/mL of IFNγ or left untreated. At 30 minutes after treatment, cells were solubilized and nuclear extracts were isolated as described in Materials and Methods. Nuclear extracts were mixed with the GAS element of the IRF1 gene with or without addition of unlabeled probe and with or without anti-STAT1 antibody. Resulting complexes were resolved by electrophoresis on a 4.5% native polyacrylamide gel. In a second experiment (B), MRC5 cells were exposed to 1 PFU per cell of the AD169 strain of HCMV or left uninfected. At 12 hours after infection, cells were solubilized and processed as described above. The black arrow indicates the shifted bands and the grey arrow indicates the supershifted bands.
Figure 6.
Reduced binding of STAT1 to GAS elements in the presence of IE1. Film image of an Electrophoretic Mobility Shift Assay shows a reduction in the amount of IRF1 GAS element probe that is shifted upon mixture with nuclear extracts from MRC5 cells ectopically expressing IE1 (A) and HCMV infected cells (B). In A, MRC5 cells were nucleofected with plasmid harboring full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 24 hours after nucleofection, cells were exposed to 100 U/mL of IFNγ or left untreated. At 30 minutes after treatment, cells were solubilized and nuclear extracts were isolated as described in Materials and Methods. Nuclear extracts were mixed with the GAS element of the IRF1 gene with or without addition of unlabeled probe and with or without anti-STAT1 antibody. Resulting complexes were resolved by electrophoresis on a 4.5% native polyacrylamide gel. In a second experiment (B), MRC5 cells were exposed to 1 PFU per cell of the AD169 strain of HCMV or left uninfected. At 12 hours after infection, cells were solubilized and processed as described above. The black arrow indicates the shifted bands and the grey arrow indicates the supershifted bands.
![Viruses 06 01502 g006]()
2.6. STAT1 Fails to Co-Immunoprecipitate with IE1
In the EMSA studies, the size of the shifted band in the presence or absence of STAT1 antibody is the same whether or not IE1 is present. This argues against a scenario in which IE1 acts by directly binding to STAT1-bound GAS elements. In order to test this directly, we carried out co‑immunoprecipitation studies. MRC-5 fibroblast cells were exposed to 3 PFU per cell of the AD169 strain of HCMV. At 12 hours after infection, cells were treated with interferon for 30 minutes. Cells were solubilized and IE1 was isolated using an antibody that recognizes an epitope in exon 2. Under these conditions, we found no evidence of an interaction between STAT1 and IE1 (
Figure 7A). Similarly, STAT1 failed to specifically co-immunoprecipitate with either the full length or the Δ345–491 mutant of IE1 in nucleofection studies (
Figure 7B, left panel). We did observe a small amount of STAT1 isolated from cells expressing both the full length IE1 and the C-terminal mutant upon longer exposure (right panel). However, in several replicate experiments, similar amounts of STAT1 were also observed in immunoprecipitations performed using lysates of empty-vector-transfected cells (data not shown). We therefore conclude that the small amount of STAT1 isolated upon isolation of IE1 and the Δ345–491 mutant IE1 represents nonspecific binding. These studies suggest that IE1does not require a sustained interaction with STAT1 to disrupt IFNγ signaling.
A blockade in IFNγ-mediated regulation of several genes is observed in murine cytomegalovirus (MCMV) infected macrophages [
25], and the MCMV M27 protein has been shown to disrupt IFNγ signaling through a novel, STAT2-dependent mechanism [
26]. Therefore, we examined whether STAT2 co-immunoprecipitates with IE1 in IFNγ treated human fibroblasts. As expected based previously published studies [
21,
22,
23,
24], we did observe an interaction of STAT2 with the full length IE1 in total lysates of nucleofected cells and HCMV infected cells. Similar to previously published results, this interaction maps to the C-terminal region of IE1 and the Δ345–491 mutant does not appear to interact with STAT2 under these conditions (
Figure 7B). In addition, very little or no phosphorylated STAT2 accumulated in nuclei of MRC5 human fibroblasts in response to IFNγ treatment, though phosphorylated STAT2 was observed in nuclei after IFNβ treatment (
Figure 8). Our findings raise the possibility that IFNγ signaling is different in murine and human fibroblasts. Specifically, it seems that murine, but not human fibroblasts, utilize phosphorylated STAT2 to transduce and amplify signals by IFNγ. Together, these studies suggest that IE functions via a novel mechanism in the nucleus to interfere with the type II IFN signal transduction pathway.
An important question raised by our studies is whether IE1 employs the same strategy, or entirely different strategies to disrupt type I and type II interferon signaling. Relevant are the following observations: (i) co-immunoprecipation studies using total lysates indicate an interaction between the HCMV IE1 acidic domain and STAT2 in human cells consistent with other studies ([
21,
22,
23,
24] and this report), and IE1 and STAT2 colocalize in ND10 structures and metaphase chromosomes [
23]; (ii) in human fibroblasts, phophorylated STAT2 does not accumulate in the nucleus upon IFNγ treatment ([
24] and this report); (iii) there are conflicting reports of an interaction between IE1 of the Towne strain and STAT1 [
21,
23] and we were unable to find compelling evidence for a specific interaction between IE1 of strain AD169 and STAT1; and (iv) nevertheless, IE1 expression interferes with binding of STAT1 molecules to GAS elements. Based on these observations, it is tempting to conclude that an interaction between IE1 and STAT2 is necessary and sufficient to interfere with type I IFN signaling. However, such an interaction does not readily account for the activity of IE1 in attenuating binding of STAT1 molecules to GAS elements in response to type II interferon. On the other hand, the ability of IE1 to disrupt STAT1 binding to target DNA elements through an indirect mechanism could impede signals transduced by both type I and type II interferons. We suggest further studies are needed to resolve the role of IE1 in these signaling pathways.
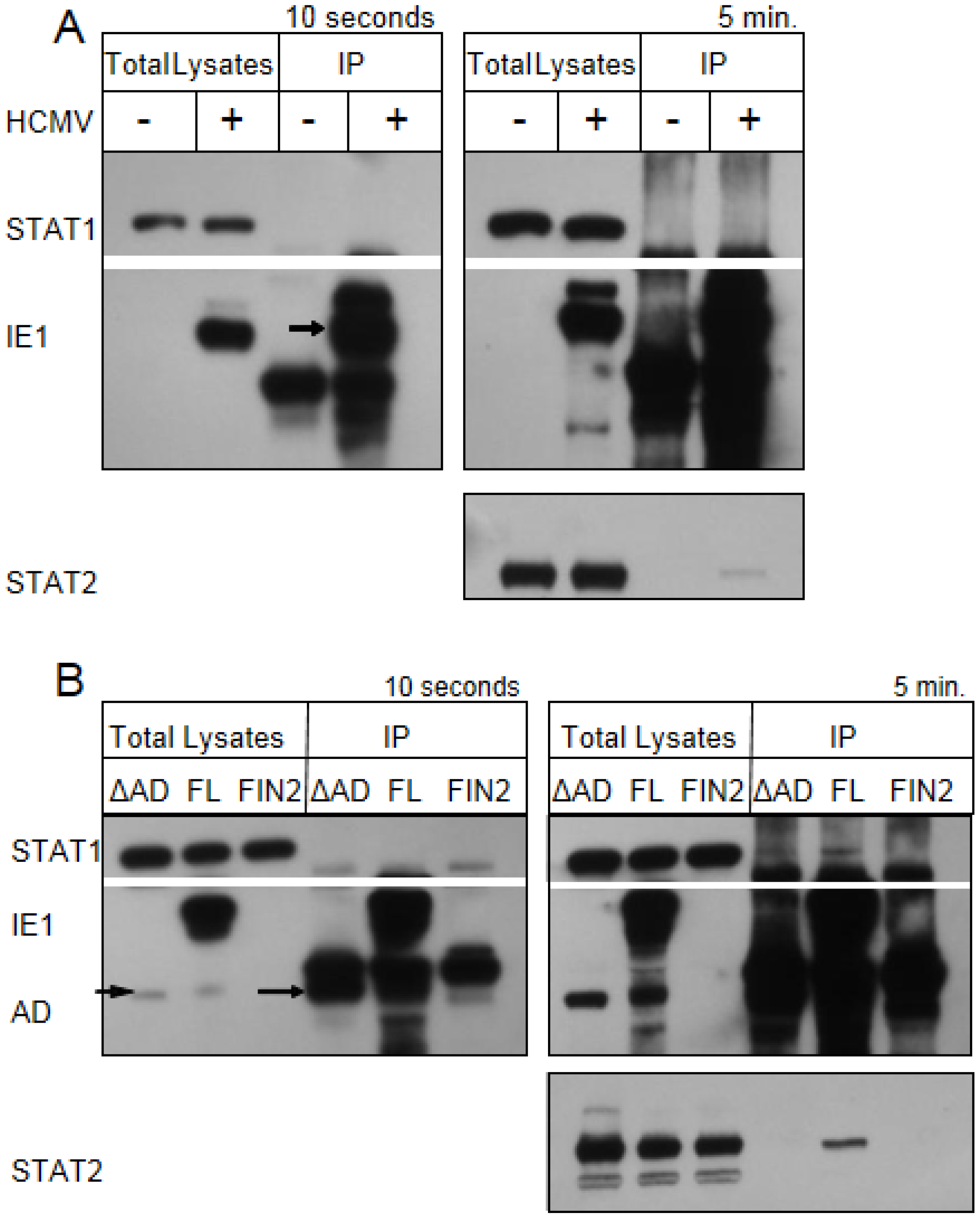
Figure 7.
IE1 does not interact with STAT1 by co-immunoprecipitation. Film images of electrophoretically separated cell lysates and proteins isolated by immunoprecipation. (A) MRC5 cells were exposed to HCMV AD169 at 3 PFU per cell or left untreated. At 12 hours after infection cells were treated with 100 U/mL of IFNγ for 30 minutes. The cells were harvested and solubilized. 350 µg of total protein from each sample was reacted with anti‑IE1 antibody (mouse monoclonal Mab810). The isolated proteins along with the 10 µg total lysates were resolved by denaturing PAGE and transferred to a nitrocellulose membrane. The membrane was reacted with antibodies for STAT1, IE1 and STAT2 and the antibody reactive bands were visualized by chemiluminescence. The left panel shows films exposed for 10 seconds and the right panel shows films exposed for 5 minutes. The arrow indicates the position of the isolated, full length IE1. (B) MRC5 cells were nucleofected with full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). 48 hours after nucleofection cells were exposed to 100 U/mL of IFNγ for 30 minutes. The preparation of cell lysates, immunoprecipitations, PAGE and western hybridization and detection were carried out as above. The left panel shows films exposed for 10 seconds and the right panel shows films exposed for 5 minutes. The short and long arrows indicate the position of the Δ345–491 IE1 in the total lysate and immunoprecipitates respectively.
Figure 7.
IE1 does not interact with STAT1 by co-immunoprecipitation. Film images of electrophoretically separated cell lysates and proteins isolated by immunoprecipation. (A) MRC5 cells were exposed to HCMV AD169 at 3 PFU per cell or left untreated. At 12 hours after infection cells were treated with 100 U/mL of IFNγ for 30 minutes. The cells were harvested and solubilized. 350 µg of total protein from each sample was reacted with anti‑IE1 antibody (mouse monoclonal Mab810). The isolated proteins along with the 10 µg total lysates were resolved by denaturing PAGE and transferred to a nitrocellulose membrane. The membrane was reacted with antibodies for STAT1, IE1 and STAT2 and the antibody reactive bands were visualized by chemiluminescence. The left panel shows films exposed for 10 seconds and the right panel shows films exposed for 5 minutes. The arrow indicates the position of the isolated, full length IE1. (B) MRC5 cells were nucleofected with full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). 48 hours after nucleofection cells were exposed to 100 U/mL of IFNγ for 30 minutes. The preparation of cell lysates, immunoprecipitations, PAGE and western hybridization and detection were carried out as above. The left panel shows films exposed for 10 seconds and the right panel shows films exposed for 5 minutes. The short and long arrows indicate the position of the Δ345–491 IE1 in the total lysate and immunoprecipitates respectively.
![Viruses 06 01502 g007]()
Many DNA viruses encode proteins that target the JAK- STAT signaling pathway including adenovirus E1A [
49,
50], human papilloma virus (HPV) E7 protein [
51], hepatitis B virus (HBV) terminal protein [
52], and polyoma virus large T antigen [
53]. Among the herpesviruses, the EBV immediate early protein BZLF1 has been shown to interfere with STAT1 phosphorylation and nuclear translocation in response to IFNγ, as well as IFNγ receptor expression [
54]. Similarly, herpes simplex virus (HSV) was shown to interfere with type I IFN signaling by decreasing the levels of several signaling molecules, including JAK1 and STAT2, which was in part mediated by the virion host shutoff protein encoded by
UL41 [
55,
56], by inducing suppressor of cytokine signaling-3 (SOCS3) [
57], and by the activity of the ICP27 protein in inhibiting STAT1 phosphorylation and nuclear translocation [
58]. HSV also targets the IFNγ pathway by modifying the IFNγR1 through the activity of the US3 and UL13 protein kinases, and indirectly by the activities of virion host shutoff protein [
59]. With the partial exception of HSV virion host shutoff protein, all of these viruses target early events in the signaling pathway, especially STAT1 phosphorylation and nuclear translocation, to attenuate IFN mediated signaling. Our findings indicate that HCMV does not target these proximal events in IFN signaling but rather it reduces STAT1 binding to target promoter elements in the nucleus. Elucidating the mechanism and consequences of this novel immune evasion strategy may provide important insights into the pathogenesis of HCMV infections and the regulation of interferon-induced gene expression.
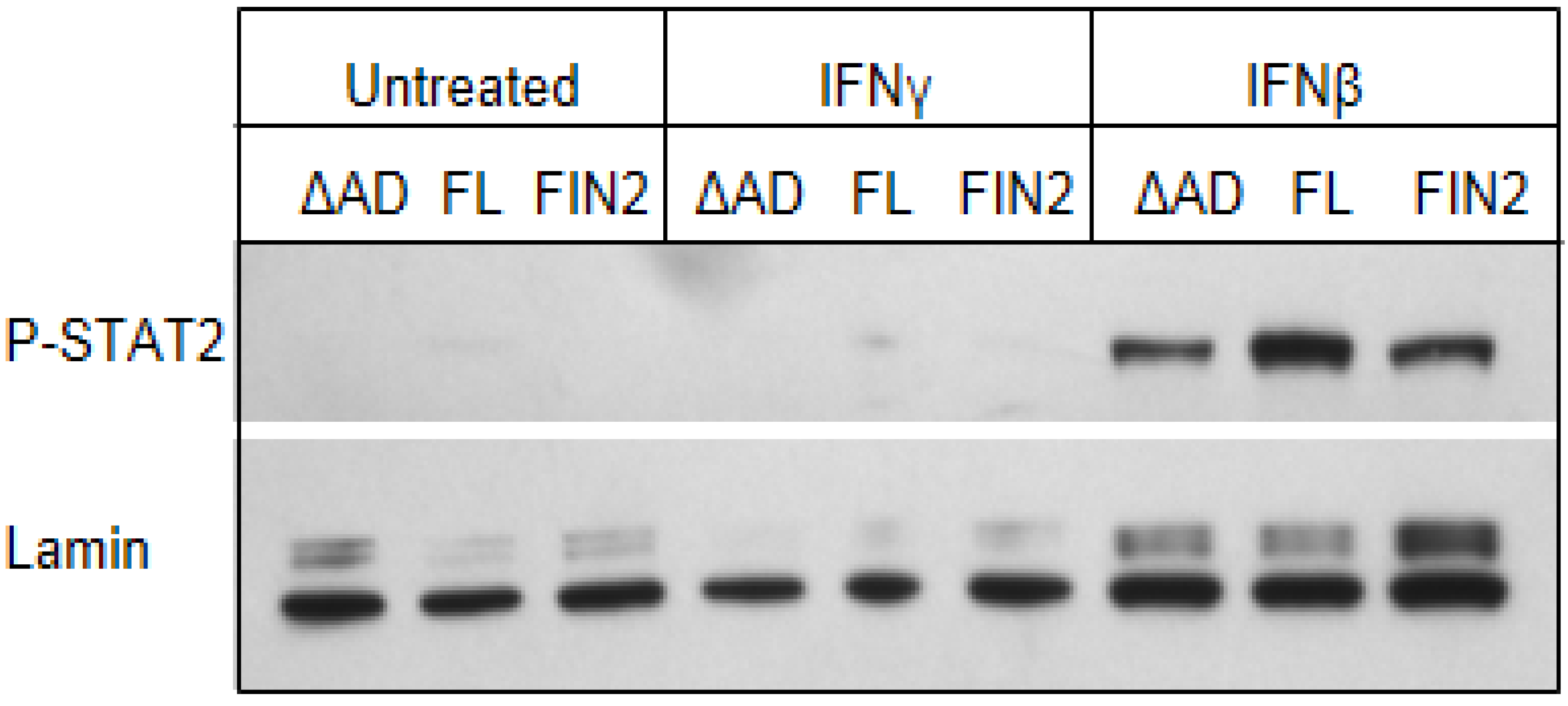
Figure 8.
Phosphorylated STAT2 does not accumulate in the nucleus of IFNγ-treated MRC-5 cells. Film images of electrophoretically separated nuclear lysates reacted with antibodies to p-STAT2 and Lamin (loading control). MRC5 cells were nucleofected with full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 72 hours after nucleofection, cells were exposed to IFNγ or IFNβ for 30 minutes or left untreated. Cells were harvested and nuclear and cytoplasmic fractions were separated. Equal amounts of protein from the nuclear lysates were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies.
Figure 8.
Phosphorylated STAT2 does not accumulate in the nucleus of IFNγ-treated MRC-5 cells. Film images of electrophoretically separated nuclear lysates reacted with antibodies to p-STAT2 and Lamin (loading control). MRC5 cells were nucleofected with full length IE1 (FL), the Δ345–491 C-terminal truncation IE1 (ΔAD) or empty vector (FIN2). At 72 hours after nucleofection, cells were exposed to IFNγ or IFNβ for 30 minutes or left untreated. Cells were harvested and nuclear and cytoplasmic fractions were separated. Equal amounts of protein from the nuclear lysates were subjected to electrophoresis in a denaturing polyacrylamide gel. Proteins were transferred to nitrocellulose sheets and reacted with the indicated antibodies.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







