
A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate: The Synthesis, Antiviral Activity and Mechanism of Antiviral Action Against Two Alphaherpesviruses

Abstract
:1. Introduction
2. Results
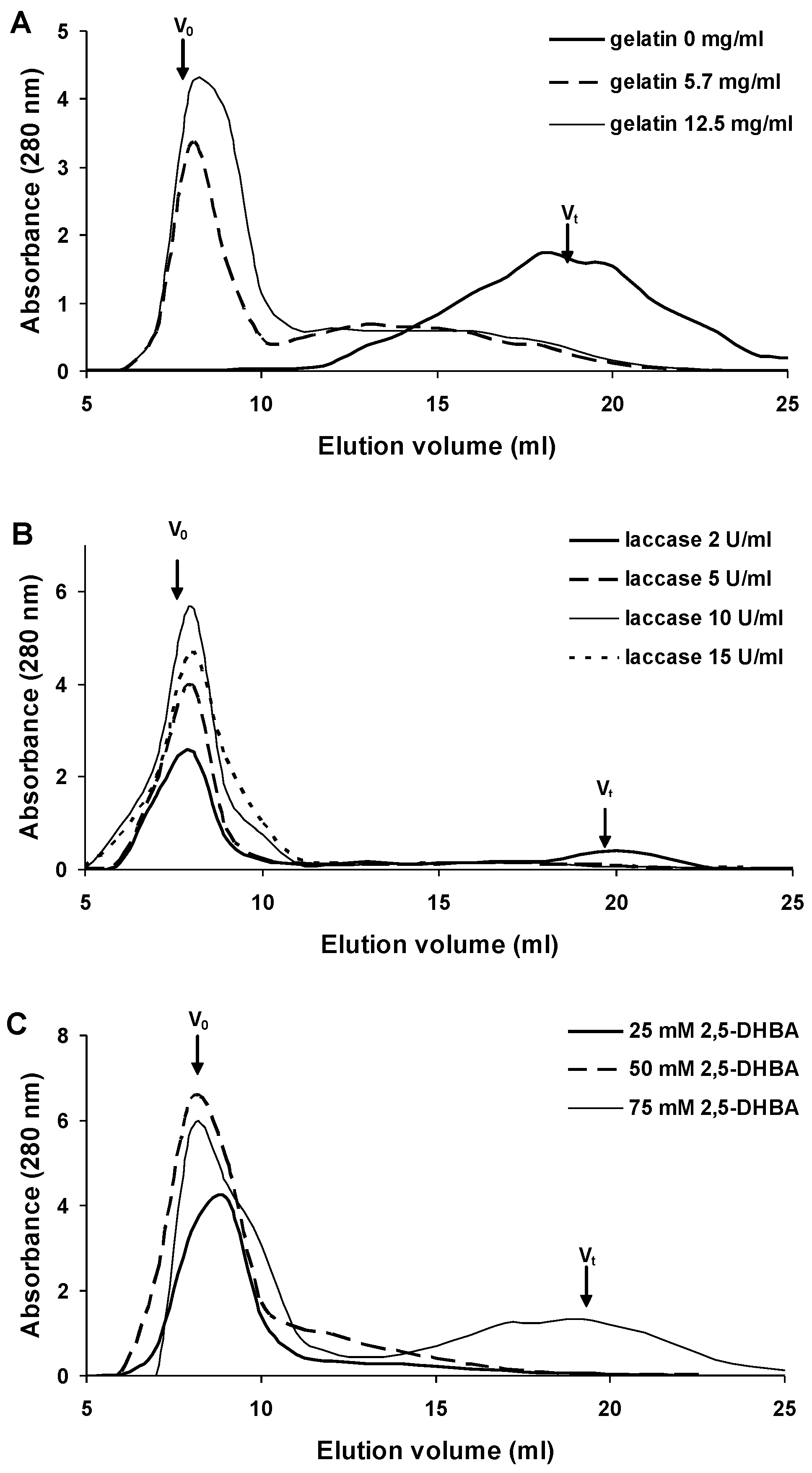
2.1. Synthesis of 2,5-DHBA–Gelatin Conjugate

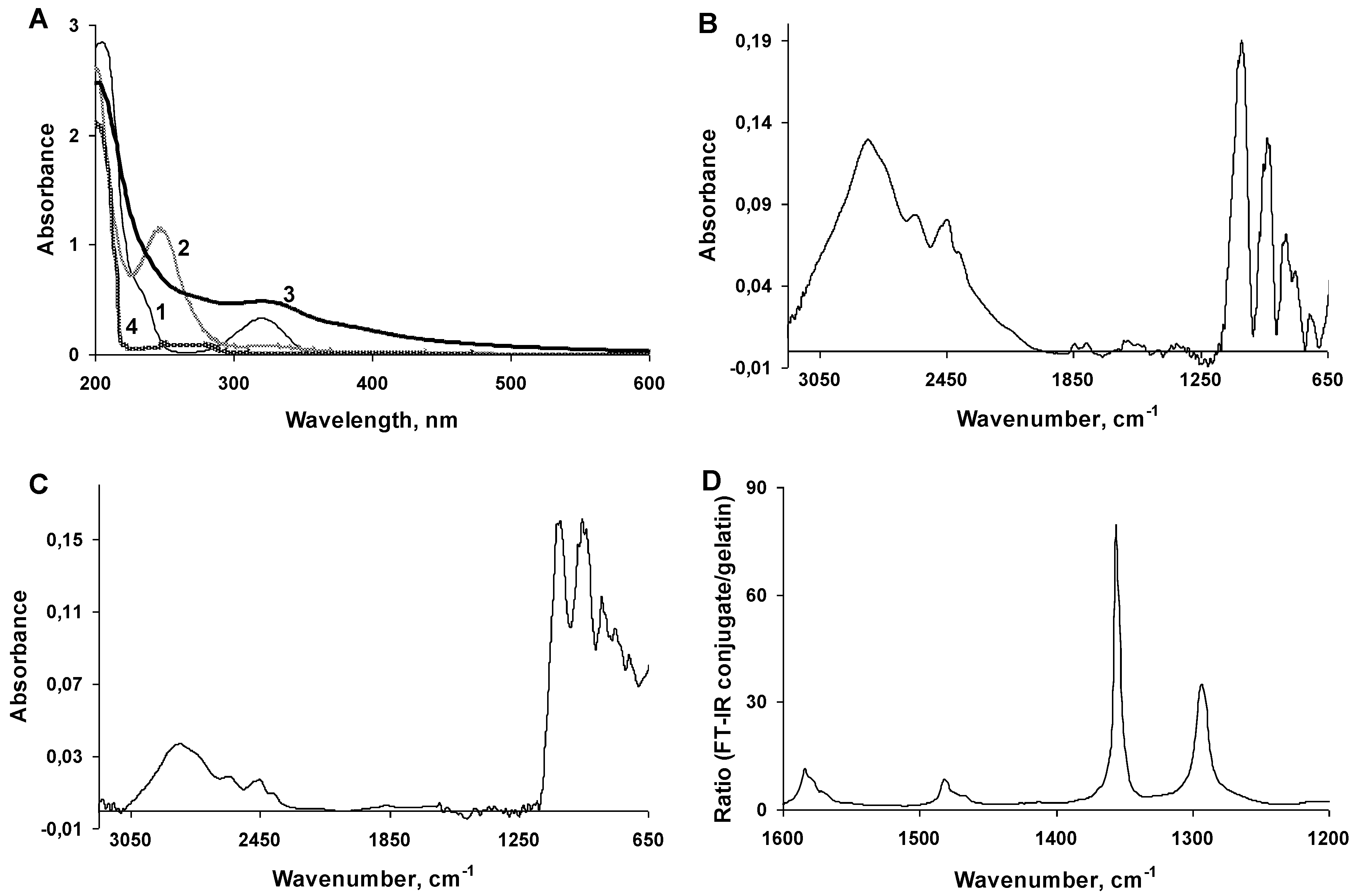
2.2. Characterization of the 2,5-DHBA–Gelatin Conjugate


2.3. Cytotoxicity and Antiproliferative Activity of the 2,5-DHBA–Gelatin Conjugate in Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Concentration, µg/mL | Cytotoxicity (%) a | Antiproliferative Activity (%) a | ||||
|---|---|---|---|---|---|---|---|
| BHK-21 | Vero | MDBK | BHK-21 | Vero | MDBK | ||
| 2,5-DHBA–gelatin | 0 | 100 | 100 | 100 | 100 | 100 | 100 |
| 62.5 | 111 ± 15 | 94 ± 10 | n.d. | 107 ± 7 | 90 ± 16 | n.d. | |
| 125.0 | 95 ± 12 | 99 ± 8 | 107 ± 12 | 102 ± 15 | 92 ± 11 | 96 ± 10 | |
| 500.0 | 103 ± 11 | 91 ± 13 | n.d. | 99 ± 11 | 98 ± 11 | n.d. | |
| 1000.0 | 95 ± 13 | 102 ± 11 | 95 ± 9 | 93 ± 13 | 92 ± 14 | 94 ± 12 | |
| 2,5-DHBA | 50.0 | 93 ± 15 | 103 ± 16 | 111 ± 13 | n.d. | n.d. | n.d. |
2.4. Antiviral Activity of the 2,5-DHBA–Gelatin Conjugate
| Virus | 2,5-DHBA–Gelatin Preparation Number | Antiviral Activity a (IC50, µg/mL) | Selectivity Index b (CC50/IC50) | ||||
|---|---|---|---|---|---|---|---|
| BHK21 | Vero | MDBK | BHK21 | Vero | MDBK | ||
| PRV (Ka) | 1 | 3.7 ± 0.7 | 1.9 ± 0.6 | n.d. | >270 | >526 | n.d. |
| 2 | 5.5 ± 0.9 | 3.1 ± 0.5 | n.d. | >181 | >322 | n.d. | |
| 3 | 2.9 ± 0.5 | 1.5 ± 0.4 | n.d. | >344 | >667 | n.d. | |
| 4 | 2.4 ± 0.7 | 2.8 ± 0.6 | n.d. | >416 | >357 | n.d. | |
| PRV (Bartha K61) | 1 | 12.2 ± 3.3 | 8.8 ± 2.8 | n.d. | >82 | >114 | n.d. |
| 2 | 15.1 ± 4.0 | 8.3 ± 3.0 | n.d. | >66 | >120 | n.d. | |
| 4 | 8.5 ± 2.2 | 10.1 ± 2.7 | n.d. | >117 | >99 | n.d. | |
| BHV-1 (4016) | 3 | n.d. | n.d. | 0.5 ± 0.1 | n.d. | n.d. | >2000 |
| 4 | n.d. | n.d. | 0.7 ± 0.2 | n.d. | n.d. | >1430 | |
| PRV (Ka) | 2,5-DHBA | >50 | n.d. | n.d. | - | - | - |
| BoHV-1 (4016) | 2,5-DHBA | n.d. | n.d. | >50 | - | - | - |
2.5. Direct Virucidal Effect of the 2,5-DHBA–Gelatin Conjugate
| Virus | Temperature of Incubation | Concentration of 2,5-DHBA–Gelatin (µg/mL) | Virus Titer, TCID50 | Residual Infectivity (%) a |
|---|---|---|---|---|
| PRV (Ka) | 4 °C | 1000 | 1.35 × 108 | 101.5 |
| 100 | 1.62 × 108 | 121.8 | ||
| 10 | 1.25 × 108 | 92.3 | ||
| 0 | 1.33 × 108 | 100.0 | ||
| 22 °C | 1000 | 0.95 × 108 | 90.5 | |
| 100 | 1.02 × 108 | 97.1 | ||
| 10 | 0.94 × 108 | 89.5 | ||
| 0 | 1.05 × 108 | 100.0 | ||
| 37 °C | 1000 | 0.78 × 108 | 104.0 | |
| 100 | 0.67 × 108 | 89.3 | ||
| 10 | 0.89 × 108 | 118.8 | ||
| 0 | 0.75 × 108 | 100.0 | ||
| BoHV-1 (4016) | 4 °C | 1000 | 5.6 × 106 | 86.2 |
| 0 | 6.5 × 106 | 100.0 | ||
| 22 °C | 1000 | 5.5 × 106 | 102.0 | |
| 0 | 5.4 × 106 | 100.0 | ||
| 37 °C | 1000 | 2.8 × 106 | 87.5 | |
| 0 | 3.2 × 106 | 100.0 |
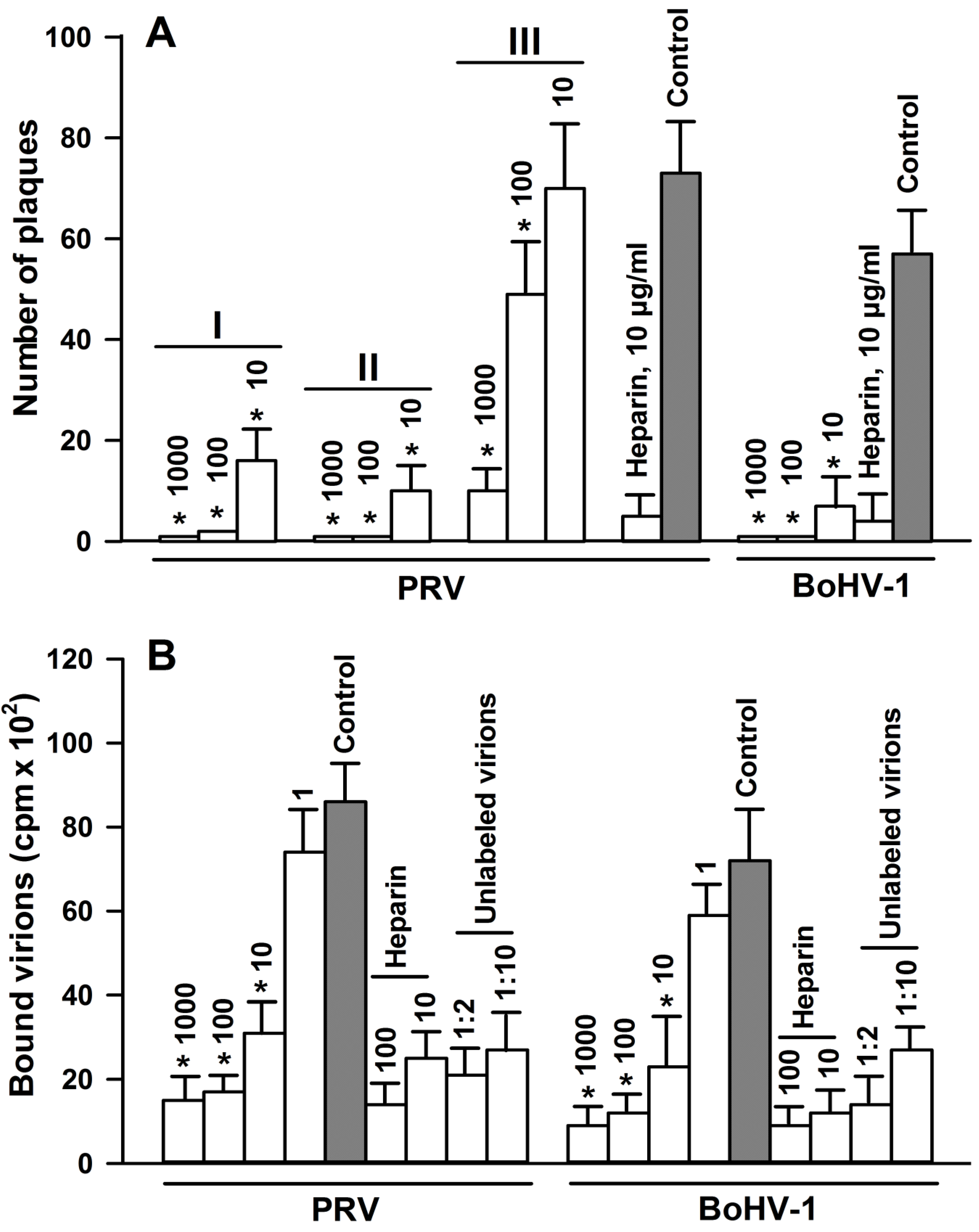
2.6. Effect of 2,5-DHBA–Gelatin on the Attachment of Viruses to Target Cells

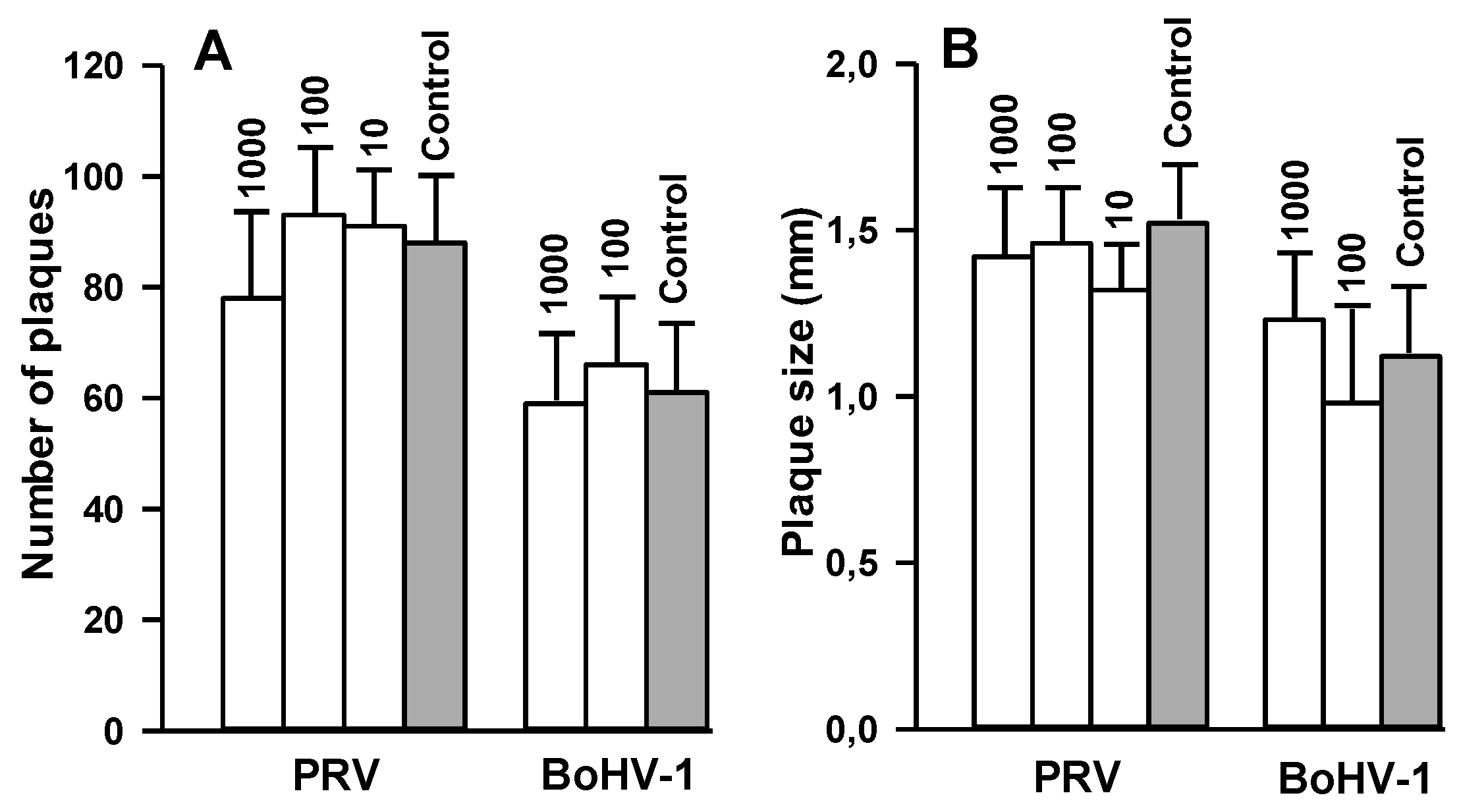
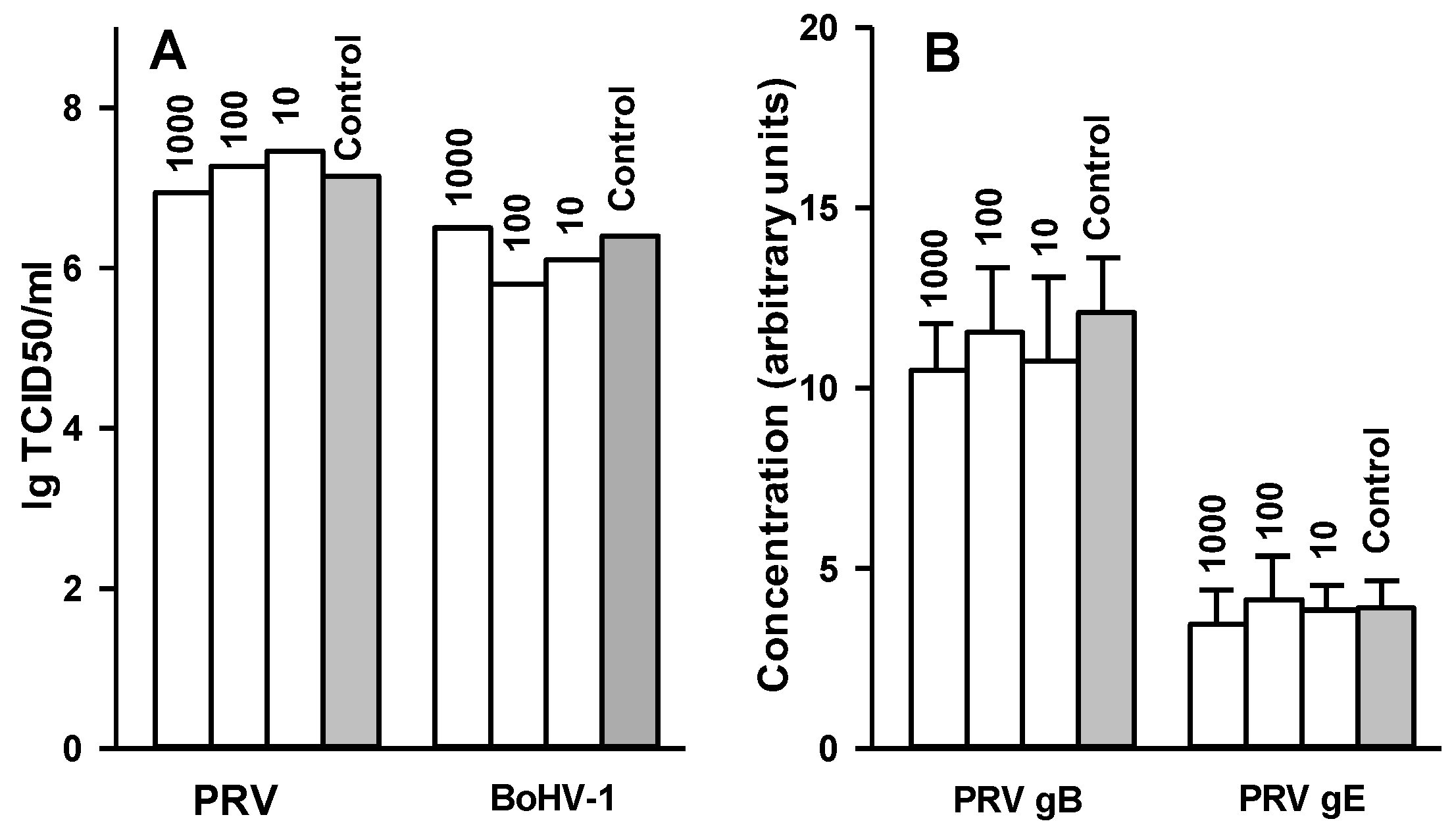
2.7. Effect of 2,5-DHBA–Gelatin on Penetration, Cell-to-Cell Spread, and Production of Infectious Virus Particles


3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Cells and Viruses
4.3. The Synthesis of the 2,5-DHBA–Gelatin Conjugate
4.4. Characterization of the 2,5-DHBA–Gelatin Conjugate
4.5. Determination of In Vitro Cytotoxicity and Antiproliferative Activity of 2,5-DHBA–Gelatin Conjugate
4.6. Virus End-Point Titration Assay
4.7. Plaque Reduction Assay
4.8. Virus Adsorption Assay
4.9. Binding of Radiolabeled Virions to Cells
4.10. Virus Penetration Assay
4.11. Influence of 2,5-DHBA–Gelatin on Cell-to-Cell Spread of PRV and BoHV-1
4.12. Direct Virucidal Effect of 2,5-DHBA–Gelatin on Viruses
4.13. Influence of 2,5-DHBA–Gelatin on Production of Infectious PRV and BoHV-1 Particles and Expression of PRV Glycoproteins B and E
4.14. Statistical Analysis
Author Contributions
Conflicts of Interest
References
- Lüscher-Mattli, M. Polyanions–a lost chance in the fight against HIV and other virus diseases? Antivir. Chem. Chemother. 2000, 11, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Baranova, E.O.; Shastina, N.S.; Shvets, V.I. Polyanionic inhibitors of HIV adsorption. Russ. J. Bioorg. Chem. 2011, 37, 527–542. [Google Scholar] [CrossRef]
- Nahmiasa, A.J.; Kibrick, S. Inhibitory effect of heparin on herpes simplex virus. J. Bacteriol. 1964, 87, 1060–1066. [Google Scholar]
- Harada, H.; Sakagami, H.; Nagata, K.; Oh-hara, T.; Kawazoe, Y.; Ishihama, A.; Hata, N.; Misawa, Y.; Terada, H.; Konno, K. Possible involvement of lignin structure in anti-influenza virus activity. Antivir. Res. 1991, 15, 41–49. [Google Scholar] [CrossRef]
- Ponce, N.M.; Pujol, C.A.; Damonte, E.B.; Flores, M.L.; Stortz, C.A. Fucoidans from the brown seaweed Adenocystisutricularis: extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Ichimura, T.; Otake, T.; Mori, H.; Maruyama, S. HIV-1 protease inhibition and anti-HIV effect of natural and synthetic water-soluble lignin-like substances. Biosci. Biotechnol. Biochem. 1999, 63, 2202–2204. [Google Scholar] [CrossRef] [PubMed]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Snoeck, R.; Pauwels, R.; de Clercq, E. Sulfated polysaccharides are potent and selective inhibitors of various enveloped viruses, including herpes simplex virus, cytomegalovirus, vesicular stomatitis virus, and human immunodeficiency virus. Antimicrob. Agents Chemother. 1988, 32, 1742–1745. [Google Scholar] [CrossRef] [PubMed]
- Thiel, K.-D.; Klöcking, R.; Schweizer, H.; Sprössig, M. In vitro studies of the antiviral activity of ammonium humate against herpes simplex virus type 1 and type 2. Zentralblattfuer Bakteriologie Mikrobiologie und Hygiene 1 Abt Originale A 1977, 239, 304–321. [Google Scholar]
- Ishihara, C.; Yoshimatsu, K.; Tsuji, M.; Arikawa, J.; Saiki, I.; Tokura, S.; Azuma, I. Anti-viral activity of sulfated chitin derivatives against Friend murine leukaemia and herpes simplex type-1 viruses. Vaccine 1993, 11, 670–674. [Google Scholar] [CrossRef]
- Mbemba, E.; Chams, V.; Gluckman, J.C.; Klatzmann, D.; Gattegno, L. Molecular interaction between HIV-1 major envelope glycoprotein and dextran sulfate. Biochim. Biophys. Acta 1992, 1138, 62–67. [Google Scholar] [CrossRef]
- Choi, Y.S.; Yoshida, T.; Mimura, T.; Kaneko, Y.; Nakashima, H.; Yamamoto, N.; Uryu, T. Synthesis of sulfated octadecylribo-oligosaccharides with potent anti-AIDS virus activity by ring-opening polymerization of a 1,4-anhydroribose derivative. Carbohydr. Res. 1996, 282, 113–123. [Google Scholar] [CrossRef]
- Schols, D.; de Clercq, E.; Balzarini, J.; Baba, M.; Witvrouw, M.; Hosoya, M.; Andrei, G.; Snoeck, R.; Neyts, J.; Pauwels, R.; et al. Sulphated polymers are potent and selective inhibitors of various enveloped viruses, including herpes simplex virus, cytomegalovirus, vesicular stomatitis virus, respiratory syncytial virus, and toga-, arena- and retroviruses. Antivir. Chem. Chemother. 1990, 1, 233–240. [Google Scholar] [CrossRef]
- Raghuraman, A.; Tiwari, V.; Zhao, Q.; Shukla, D.; Debnath, A.K.; Desai, U.R. Viral inhibition studies on sulfated lignin, a chemically modified biopolymer and a potential mimic of heparan sulfate. Biomacromolecules 2007, 8, 1759–1763. [Google Scholar] [CrossRef] [PubMed]
- Roner, M.R.; Carraher, C.E.; Shahi, K.; Barot, G. Antiviral activity of metal-containing polymers-organotin and cisplatin-like polymers. Materials 2011, 4, 991–1012. [Google Scholar]
- Rusconi, S.; Moonis, M.; Merrill, D.P.; Pallai, P.V.; Neidhardt, E.A.; Singh, S.K.; Willis, K.J.; Osburne, M.S.; Profy, A.T.; Jenson, J.C.; et al. Naphthalene sulfonate polymers with cd4-blocking and anti-human immunodeficiency virus type 1 activities. Antimicrob. Agents Chemother. 1996, 40, 234–236. [Google Scholar] [PubMed]
- Thakkar, J.N.; Tiwari, V.; Desai, U.R. Nonsulfated, cinnamic acid-based lignins are potent antagonists of HSV-1 entry into cells. Biomacromolecules 2010, 11, 1412–1416. [Google Scholar] [CrossRef]
- Schols, D.; Wutzler, P.; Klöcking, R.; Helbig, B.; de Clercq, E. Selective inhibitory activity of polyhydroxycarboxylates derived from phenolic compounds against human immunodeficiency virus replication. J. Acquir. Immune Defic. Syndr. 1991, 4, 677–685. [Google Scholar] [PubMed]
- Neyts, J.; Snoeck, R.; Wutzler, P.; Cushman, M.; Klöcking, R.; Helbig, B.; Wang, P.; de Clercq, E. Poly(hydroxy)carboxylates as selective inhibitors of cytomegalovirus and herpes simplex virus replication. Antivir. Chem. Chemother. 1992, 3, 215–222. [Google Scholar] [CrossRef]
- Mikolasch, A.; Schauer, F. Fungal laccases as tools for the synthesis of new hybrid molecules and biomaterials. Appl. Microbiol. Biotechnol. 2009, 82, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Bollag, J.M.; Leonowicz, A. Comparative studies of extracellular fungal laccases. Appl. Environ. Microbiol. 1984, 48, 849–854. [Google Scholar] [PubMed]
- Mayer, A.M.; Harel, E. Polyphenol oxidases in plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] [CrossRef]
- Sharma, P.; Goel, R.; Capalash, N. Bacterial laccases. World J. Microbiol. Biotechnol. 2007, 23, 823–832. [Google Scholar] [CrossRef]
- Beckett, R.P.; Zavarzina, A.G.; Liers, C. Oxidoreductases and cellulases in lichens: possible roles in lichen biology and soil organic matter turnover. Fungal Biol. 2013, 117, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Yatsu, J.; Asano, T. Cuticle laccase of the silkworm, Bombyxmori: purification, gene identification and presence of its inactive precursor in the cuticle. Insect Biochem. Mol. Biol. 2009, 39, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Kanitskaya, L.V.; Seleznev, S.N.; Medvedeva, S.A.; Kupriyanovich, Y.N.; Turchaninov, V.K. Dehydrogenation polymerization of 2,6-dimethoxyphenol. Polym. Sci. Ser. A 2003, 45, 92–98. [Google Scholar]
- Aktas, N.; Tanyolac, A. Reaction conditions for laccase-catalyzed polymerization of catechol. Bioresour. Technol. 2003, 87, 209–214. [Google Scholar] [CrossRef]
- Kupriyanovich, Y.N.; Medvedeva, S.A.; Rokhin, A.V.; Kanitskaya, L.V. Regioselectivity of ferulic acid polymerization catalyzed by oxidases. Russ. J. Bioorg. Chem. 2007, 33, 516–522. [Google Scholar] [CrossRef]
- Mai, C.; Schormann, W.; Hüttermann, A. Chemo-enzymatically induced copolymerization of phenolics with acrylate compounds. Appl. Microbiol. Biotechnol. 2001, 55, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Selinheimo, E.; Lampila, P.; Mattinen, M.-L.; Buchert, J. Formation of protein-oligosaccharide conjugates by laccase and tyrosinase. J. Agric. Food Chem. 2008, 56, 3118–3128. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.E.; Kurisawa, M.; Uyama, H.; Kobayashi, S. Enzymatic synthesis and antioxidant property of gelatin-catechin conjugates. Biotechnol. Lett. 2003, 25, 1993–1997. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, G.; Kraemer, K.; Fuessel, S.; Puoci, F.; Curcio, M.; Spizzirri, U.G.; Altimari, I.; Iemma, F. Biological activity of a gallic acid-gelatin conjugate. Biomacromolecules 2010, 11, 3309–3315. [Google Scholar] [CrossRef] [PubMed]
- Roizman, B.; Sears, A.E. Herpes simplex viruses and their replication. In Fields Virology, 3rd ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Chanock, R.M., Melnick, J.L., Monath, T.P., Roizman, B., Straus, S.E., Eds.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1996; pp. 2231–2295. [Google Scholar]
- Herold, B.C.; WuDunn, D.; Soltys, N.; Spear, P.G. Glycoprotein C of herpes simplex virus type 1 plays a principal role in the absorption of virus to cells and in infectivity. J. Virol. 1991, 65, 1090–1098. [Google Scholar]
- Herold, B.C.; Visalli, R.J.; Susmarski, N.; Brandt, C.R.; Spear, P.G. Glycoprotein C-independent binding of herpes simplex virus to cells requires cell surface heparansulphate and glycoprotein B. J. Gen. Virol. 1994, 75, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, X.; van DrunenLittel-van den Hurk, S.; Babiuk, L.A. Characterization of cell-binding properties of bovine herpesvirus 1 glycoproteins B, C, and D: Identification of dual cell-binding function of gB. J. Virol. 1995, 69, 4758–4768. [Google Scholar] [PubMed]
- Mettenleiter, T.C.; Zsak, L.; Zuckermann, F.; Sugg, N.; Kern, H.; Ben-Porat, T. Interaction of glycoprotein gIII with a cellular heparinlike substance mediates adsorption of pseudorabies virus. J. Virol. 1990, 64, 278–286. [Google Scholar] [PubMed]
- Okazaki, K.; Matsuzaki, T.; Sagahara, Y.; Okada, J.; Hasebe, M.; Iwamura, Y.; Ohnishi, M.; Kanno, T.; Shimizu, M.; Honda, E.; Kono, Y. BHV-1 absorption is mediated by the interaction of glycoprotein gIII with heparinlike moiety on the cell surface. Virology 1991, 181, 666–670. [Google Scholar] [CrossRef]
- Trybala, E.; Liljeqvist, J.A.; Svennerholm, B.; Bergström, T.J. Herpes simplex virus types 1 and 2 differ in their interaction with heparan sulfate. Virology 2000, 74, 9106–9114. [Google Scholar] [CrossRef]
- WuDunn, D.; Spear, P.G. Initial interaction of herpes simplex virus with cells is binding to heparan sulfate. J. Virol. 1989, 63, 52–58. [Google Scholar] [PubMed]
- Vrublevskaya, V.V.; Kornev, A.N.; Smirnov, S.V.; Morenkov, O.S. Cell-binding properties of glycoprotein B of Aujeszky's disease virus. Virus Res. 2002, 86, 7–19. [Google Scholar] [CrossRef]
- Campadelli-Fiume, G.; Menotti, L. Entry of alphaherpesviruses into the cell. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 93–111. [Google Scholar]
- Spear, P.G.; Eisenberg, R.J.; Cohen, G.H. Three classes of cell surface receptors for alphaherpesvirus entry. Virology 2000, 275, 1–8. [Google Scholar] [CrossRef]
- Wittman, G. Spread and control of Aujeszky's Disease. Comp. Immun. Microbiol. Infect. Dis. 1991, 14, 165–173. [Google Scholar] [CrossRef]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [PubMed]
- Toscano, G.; Colarieti, M.L.; Greco, G., Jr. Oxidative polymerisation of phenols by a phenol oxidase from green olives. Enz. Microb. Technol. 2003, 33, 47–54. [Google Scholar] [CrossRef]
- Einerson, N.J.; Stevens, K.R.; Kao, W.J. Synthesis and physicochemical analysis of gelatin-based hydrogels for drug carrier matices. Biomaterials 2002, 24, 509–524. [Google Scholar] [CrossRef]
- Hollmann, F.; Arends, I.W.C.E. Enzyme initiated radical polymerizations. Polymers 2012, 4, 759–793. [Google Scholar] [CrossRef]
- Spizzirri, U.G.; Iemma, F.; Puoci, F.; Cirillo, G.; Curcio, M.; Parisi, O.I.; Picci, N. Synthesis of antioxidant polymers by grafting of gallic acid and catechin on gelatin. Biomacromolecules 2009, 10, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Sarkanen, K.V.; Ludwig, C.H. (Eds.) Lignins: Occurrence, Formation, Structure and Reactions; John Wiley and Sons, Inc.: New York, NY, USA, 1971.
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions; Wiley-Interscience: New York, NY, USA, 1982. [Google Scholar]
- Witvrouw, M.; de Clercq, E. Sulfated polysaccharides extracted from sea algae as potent antiviral drugs. Gen. Pharmacol. 1997, 29, 497–511. [Google Scholar] [CrossRef]
- Ben-Porat, T.; DeMarchi, J.M.; Lomniczi, B.; Kaplan, A.S. Role of glycoproteins of pseudorabies virus in eliciting neutralizing antibodies. Virology 1986, 154, 325–334. [Google Scholar] [CrossRef]
- Morenkov, O.S.; Fodor, N.; Sobko, Y.A.; Fodor, I. Immunological characterisation of glycoprotein E of Aujeszky’s disease virus. Virus Res. 1997, 51, 65–79. [Google Scholar] [CrossRef]
- Zaripov, M.M.; Morenkov, O.S.; Siklodi, B.; Barna-Vetro, I.; Gyongyosi-Horvath, A.; Fodor, I. Glycoprotein B of Aujeszky’s disease virus: topographical epitope mapping and epitope-specific antibody response. Res. Virol. 1998, 160, 23–31. [Google Scholar] [CrossRef]
- Ben-Porat, T.; DeMarchi, J.M.; Kaplan, A.S. Characterization of defective interfering viral particles present in a population of pseudorabiesvirions. Virology 1974, 61, 29–37. [Google Scholar] [CrossRef]
- Lisova, Z.A.; Lisov, A.V.; Leontievsky, A.A. Two laccase isoforms of the basidiomycete Cerrena unicolor VKMF-3196. Induction, isolation and properties. J. Basic. Microbiol. 2010, 50, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hygiene 1938, 27, 493–497. [Google Scholar]
- Morenkov, O.S.; Mantsygin, I.A.; Sergeev, V.A.; Sobko, I.A.; Panchenko, O.A.; Zabello, N.E.; Morenkova, M.A. Use of monoclonal antibodies for detecting Aujeszky's disease virus in culture media by an immunoenzyme method. Vopr. Virusol. 1994, 39, 212–214. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisov, A.; Vrublevskaya, V.; Lisova, Z.; Leontievsky, A.; Morenkov, O. A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate: The Synthesis, Antiviral Activity and Mechanism of Antiviral Action Against Two Alphaherpesviruses. Viruses 2015, 7, 5343-5360. https://doi.org/10.3390/v7102878
Lisov A, Vrublevskaya V, Lisova Z, Leontievsky A, Morenkov O. A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate: The Synthesis, Antiviral Activity and Mechanism of Antiviral Action Against Two Alphaherpesviruses. Viruses. 2015; 7(10):5343-5360. https://doi.org/10.3390/v7102878
Chicago/Turabian StyleLisov, Alexander, Veronika Vrublevskaya, Zoy Lisova, Alexey Leontievsky, and Oleg Morenkov. 2015. "A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate: The Synthesis, Antiviral Activity and Mechanism of Antiviral Action Against Two Alphaherpesviruses" Viruses 7, no. 10: 5343-5360. https://doi.org/10.3390/v7102878
APA StyleLisov, A., Vrublevskaya, V., Lisova, Z., Leontievsky, A., & Morenkov, O. (2015). A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate: The Synthesis, Antiviral Activity and Mechanism of Antiviral Action Against Two Alphaherpesviruses. Viruses, 7(10), 5343-5360. https://doi.org/10.3390/v7102878