
The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses


1
Biomedicine Discovery Institute and Department of Microbiology, Monash University, Melbourne, Victoria 3800, Australia
2
Duke-NUS Medical School, Singapore 169857, Singapore
*
Author to whom correspondence should be addressed.
Viruses 2017, 9(5), 102; https://doi.org/10.3390/v9050102
Submission received: 31 March 2017
/
Revised: 5 May 2017
/
Accepted: 7 May 2017
/
Published: 10 May 2017
(This article belongs to the Special Issue Astroviruses)
{kind=link}
{kind=link}
Abstract
:Astroviruses are a diverse family of viruses that infect a wide range of mammalian and avian hosts. Here we describe the phylogenetic diversity and current classification methodology of astroviruses based on the ORF1b and ORF2 genes, highlighting the propensity of astroviruses to undergo interspecies transmission and genetic recombination which greatly increase diversity and complicate attempts at a unified and comprehensive classification strategy.
Keywords:
astrovirus; classification; avian; mammal; genetic diversity; capsid; RNA-dependent-RNA-polymerase; ORF1b; ORF21. Introduction
Astroviruses (AstVs) were first described in 1975 as small viruses that are 28–30 nm in diameter with icosahedral morphology. Astroviruses are named due to the distinct five-pointed or six-pointed star-like appearance of some virions when visualized under an electron microscope (EM); astrovirus is derived from the Greek word astron meaning star [1,2,3]. Astroviruses were first described from human infants with diarrhea and were subsequently identified in the young of numerous mammalian and avian species [4,5].
Human astroviurses (HAstVs) have been recognized as one of the major causes of acute gastroenteritis in children, associated with 2–8% of infections [6]. Transmission of HAstV occurs via the fecal-oral route, person-to-person contact, or contaminated food or water. Following an incubation period of 3–4 days, symptoms including diarrhea, vomiting, abdominal pain, and fever are often reported [7,8]. Whilst primarily associated with asymptomatic or diarrheal disease in humans, there are several reports of central nervous system (CNS) complications such as acute flaccid paralysis [9], meningitis, and encephalitis [10,11]. Animal astroviurses, have been isolated from numerous mammalian and avian species. In animals, astrovirus infection may be asymptomatic or associated with enteric disease and a range of other symptoms indicative of the involvement of other organ systems including hepatitis and nephritis in avian species [12,13], and neurological symptoms in cattle [14,15,16] and mink [17].
1.1. Genome
Astroviruses are classified within the unassigned Astroviridae family and are non-enveloped viruses characterized by a positive sense, single-stranded RNA (ssRNA) genome 6.4–7.9 kb long comprised of a 5′-untranslated region (UTR), three open reading frames (ORFs)—ORF1a, ORF1b, and ORF2, a 3′-UTR, and a poly A tail [18]. The ORF1a region encodes a non-structural polyprotein (serine protease), ORF1b encodes a polyprotein including the RNA-dependent RNA polymerase (RdRp), and ORF2 encodes the viral capsid protein [18]. A further ORF, termed ORFX, has been observed in classic HAstVs and some mammalian astroviurses, overlapping the 5′ end of ORF2 which may be translated through a leaking scanning mechanism [19]. Astroviruses exhibit several distinctive features in addition to a distinctive morphology. The viruses lack a RNA-helicase domain encoded within the genome and utilize a ribosomal frameshifting mechanism to translate the RdRp, which distinguishes the Astroviridae family from other non-enveloped ssRNA virus families such as Picornaviridae and Caliciviridae [20,21]. The greatest diversity in the genome is within the ORF2 region, which is characterized by a highly-conserved N-terminal domain (amino acids (aa) 1–424), a hypervariable domain (aa 425–688) which is believed to form the capsid spike and contribute to receptor binding, and a highly acidic C-terminal domain [22,23].
1.2. Classification
The wide host range of astroviruses and the high degree of genetic diversity present within the Astroviridae family have complicated attempts at a unified classification method. Astroviruses are classified within the unassigned Astroviridae family, which was initially comprised of a single genus (Astrovirus) based on virion morphology [18]. The classification of the Astrovirus genus within the family Astroviridae was recognized by the International Committee for Taxonomy of Viruses (ICTV) in 1995 and the classification scheme has been modified numerous times over the intervening years [24]. In 2004, two genera were recognized; Mamastrovirus (MAstV) and Avastrovirus (AAstV), that were known to infect mammalian and avian species, respectively, and viruses were classified solely on the species of origin [24]. However, the advent of sequence based characterization rendered this approach inadequate, revealing that viruses isolated from different species can be genetically similar (reflecting prevalent interspecies transmission of viruses) and also revealing a large range of diversity of viruses within a single host species. With this in mind, the classification system proposed by the ICTV Astroviridae Study Group in 2010 recommended classification based on the amino acid sequence of the ORF2 genome region, recommending that different strains of the same astrovirus species should share >75% identity in the capsid proteins [25]. Additionally, there are proposals to define distinct variants within a recognized astrovirus species, with a variant defined as sharing <93–95% nucleotide similarity to the reference or prototype strain of each species or if phylogenetic analysis is used, a distance of >0.05 based on analysis of the capsid protein [12,18].
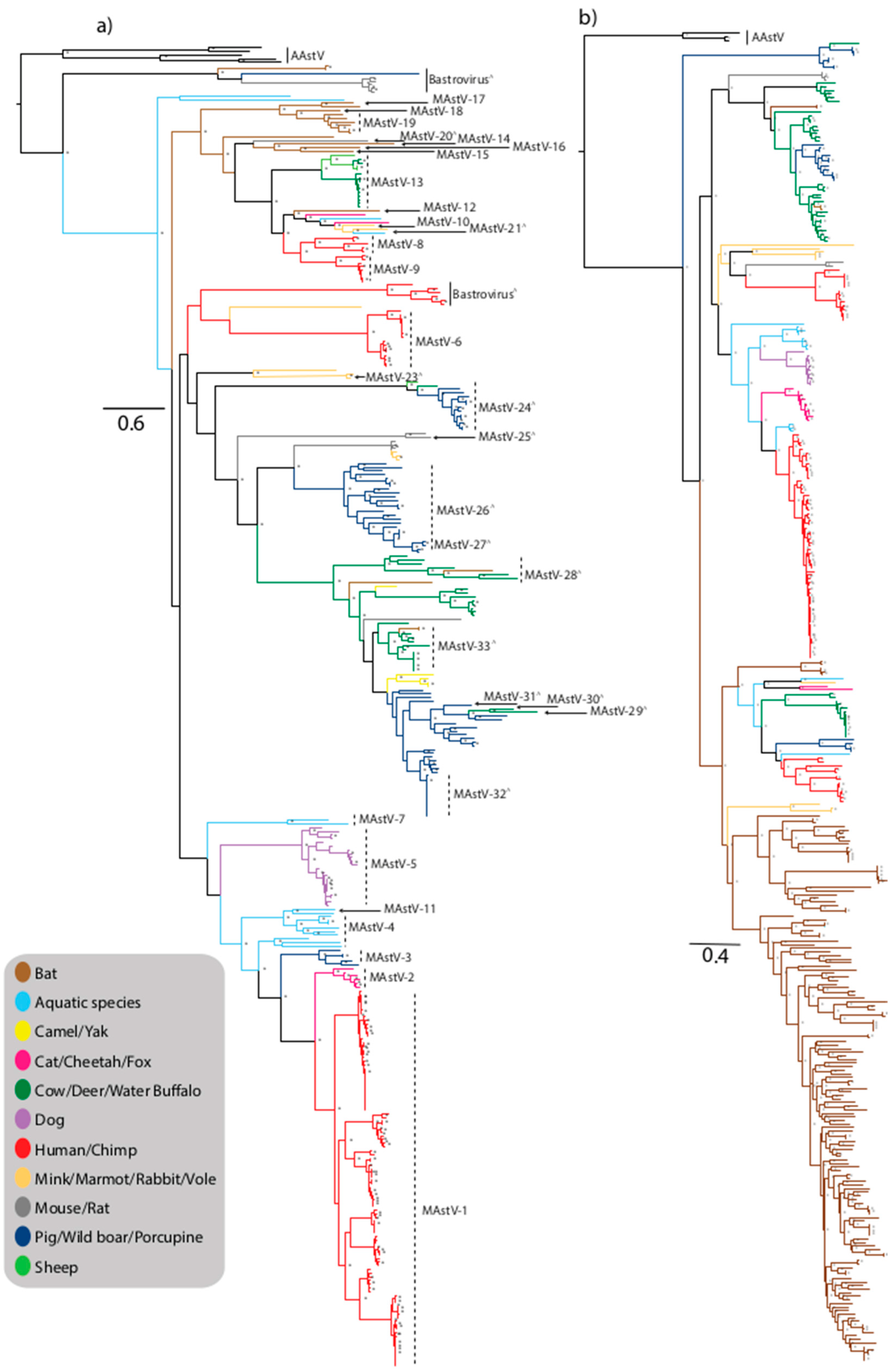
Astroviruses within the Mamastrovirus genus are derived from numerous mammalian species in addition to humans (HAstV), including farmed species such as pigs (PAstV), sheep (OAstV), cattle (BoAstV), domesticated animals including cats (FAstV), and dogs (CaAstV), rodents and small mammals including mink (MiAstV), bats (BAstV), rats (RAstV), mice, rabbit (RabAstV), fox, marmot (HHMastV), porcupine, shrew, vole, and larger species including deer (CcAstV), monkeys, water buffalo (BufAstV), yak, camel (DcAstV), and cheetah (ChAstV) (Figure 1a,b). Viruses from the Mamastrovirus genus have also been characterized from marine mammals including stellar sea lion (SslAstV) and California sea lions (CslAstV), minke whale, orca whale, and bottlenose dolphins (BdAstV) (Figure 1a,b) [26]. The current ICTV classification reveals 19 recognized species of Mamastrovirus (MAstV-1–19) within two genogroups GI and GII; Mamastrovirus 1 (GI.A-human); Mamastrovirus 2 (GI.B-feline); Mamastrovirus 3 (GI.C-porcine); Mamastrovirus 4 (GI.D-California sea lion); Mamastrovirus 5 (GI.E-canine); Mamastrovirus 6 (GI.F-human); Mamastrovirus 7 (GI.G-bottlenose dolphin); Mamastrovirus 8 (GII.A-human); Mamastrovirus 9 (GII.B-human); Mamastrovirus 10 (GII.C-mink); Mamastrovirus 11 (GII.D-California sea lion), Mamastrovirus 12 (GII.E-bat); Mamastrovirus 13 (GII.F-ovine); and Mamastroviruses 14–19 (GII.G to GII.L-bat species), and numerous other strains awaiting classification, some of which are considered as tentative new species (designated by a ∧ symbol in the phylogenetic trees) (Figure 1a) [27,28].
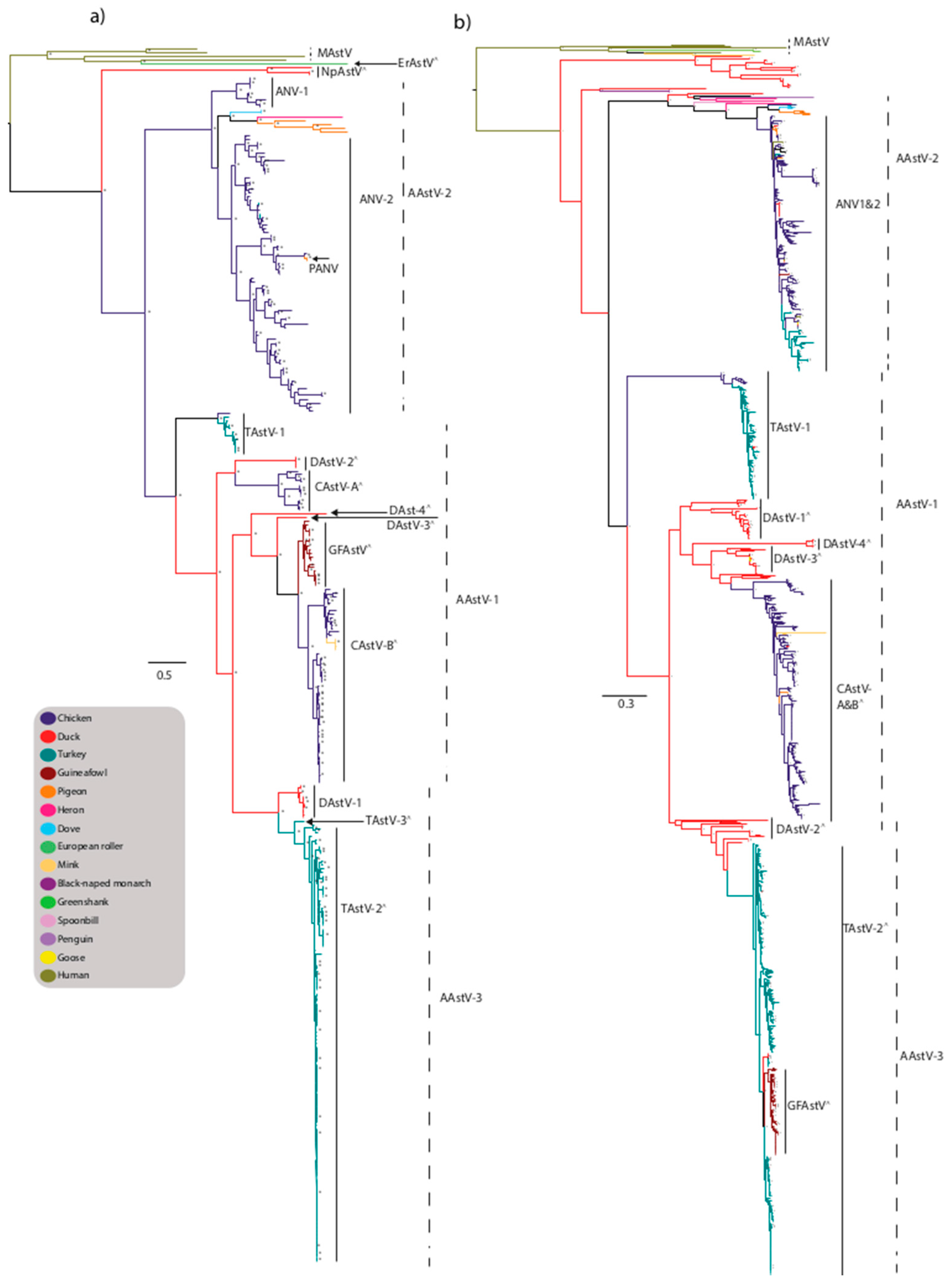
Viruses from the Avastrovirus genus have been characterized from numerous farmed avian species including turkeys (TAstV), ducks (DAstV), chicken (CAstV), guineafowl (GFAstV), pigeon (PiAstV), goose, as well as wild aquatic and terrestrial birds including heron, doves, penguins, and many other species (Figure 2a). The three species originally recognized within the genus were Avastrovirus GI.A comprised of turkey astrovirus 1 (TAstV-1), Avastrovirus GI.B comprised of avian nephritis virus 1 (ANV-1), avian nephritis virus 2 (ANV-2), and Avastrovirus GII.A comprised of turkey astrovirus 2 (TAstV-2) and duck astrovirus DAstV/C-NGB [25]. Avastrovirus GI.A, Avastrovirus GI.B, and Avastrovirus GII.A were renamed Avastrovirus 1 (AAstV-1), Avastrovirus 2 (AAstV-2), and Avastrovirus 3 (AAstV-3), respectively [25].
Currently, classification into species is based on the phylogenetic analysis of the amino acid sequence of the full length ORF2 region of the genome that encodes the capsid. However, the limited number of capsid sequences available compared to RdRp sequences makes consistent classification difficult, especially with some novel viruses incompletely sequenced. There are numerous unclassified astroviruses, particularly isolated from aquatic and terrestrial wild birds which, according to the ICTV, are “related viruses which may be members of the Avastrovirus genus but have not been approved as species” [18].
2. Human Astroviruses
Astrovirus infection in humans has been primarily associated with diarrhea and vomiting, accounting for up to 10% of sporadic gastroenteritis cases in some regions [29]. CNS complications associated with astrovirus infection have been reported in recent years, including acute flaccid paralysis, with some fatalities reported in children with underlying immune disorders [30,31].
Historically human astroviruses (HAstV) were classified into five serotypes in 1984 [32]. Subsequent molecular characterization based on viral reactivity to polyclonal antibodies and nucleotide sequence analysis led to the recognition of eight serotypes (HAstV-1–8), now termed “classic” HAstV [18,33,34]. The relatively recent advent of next generation sequencing (NGS) and metagenomic analysis has led to the identification of numerous novel strains considered “non-classic” HAstV [18]. Currently, HAstVs are classified within the species MAstV-1 (HAstV-1–8), MAstV-6 (MLB1–3), MAstV-8 (VA2/HMO-A, VA4, VA5, BF34), and MAstV-9 (VA1/HMO-C, VA3/HMO-B) [18] (Figure 1a).
2.1. MAstV-1
The MAstV-1 species is comprised of HAstV-1–8, and surveillance has revealed that HAstV-1 is the most commonly detected type in children, followed by HAstV-2–5, whereas HAstV-6–8 have been rarely detected [35]. HAstV-4 and HAstV-8 have been associated with infection of older children and longer duration of diarrhea (>7 days) [36,37]. A HAstV-4 strain was also isolated from an infant with fatal meningoencephalitis [30]. Based upon the phylogenetic analysis of the ORF2 region, different lineages within each HAstV type have been proposed; HAstV-1 (HAstV-1a–d) and HAstV-2 (HAstV-2a–d) have been divided into four lineages, whereas HAstV-3 (HAstV-3a–b) and HAstV-4 (HAstV-4a–c) have been classified into two and three lineages, respectively [38].
2.2. MAstV-6
The first “non-classic” HAstV strain characterized was MLB1, the virus was detected in a stool sample from a 3 year old Australian child with acute diarrhea in 1999; the child had previously received a liver transplant [39]. The majority of MLB1 strains characterized to date have been detected in India, Kenya, and Japan with limited detected in the USA, China, Bhutan, Egypt, Brazil, and Italy and prevalence has been reported in the range of 0.2% to 9% [40]. However, a seroepidemiologic study in the USA revealed that primary exposure to MLB1 occurs in childhood and that seropositivity reached 100% by adulthood suggesting the widespread circulation of the virus in the human population [41]. MLB2 viruses were first identified in Vellore, India [42] with the majority of strains subsequently identified in Japan, The Gambia, and Switzerland with limited detection in Turkey, USA, Kenya, China, and Thailand and prevalence reported in the range of 0.3% to 1.5% [40]. MLB2 has been associated with meningitis and other CNS complications and has been detected in immunocompromised children [10]. MLB3 viruses were first detected in India in 2004 [43], with subsequent detection in Kenya and The Gambia and the prevalence in stools ranges from 0.6% to 3.1% [40,44].
2.3. MAstV-8
There is a dual naming system for some HAstV species due to the simultaneous characterization of these viruses by different researchers; these viruses are termed VA/HMO named for VA—Virginia and HMO—Human-Mink-Ovine-like viruses, due to their genetic relatedness to previously characterized mink and sheep viruses [9]. In 2009, VA2/HMO-A strains were detected in children with non-polio acute flaccid paralysis in Nigeria, Pakistan [9], and India [42]. The prevalence of VA2/HMO-A viruses in stools ranges from 0.3% to 2.3%, with strains also detected in Egypt, Japan, USA, Kenya, and China [40]. VA4 has only been detected in Nepal [43] and the BF34 strain has only detected in Burkina Faso [45].
2.4. MAstV-9
The VA3/HMO-B viruses were first identified in Vellore, India [42] with sporadic detection in Nigeria, Pakistan, and Nepal with prevalence ranging from 0.1% to 2.3% [40]. VA1/HMO-C viruses were first detected in 2009 during an outbreak of diarrhea in children in Virginia, USA [46]. VA1/HMO-C viruses have been detected and associated with encephalitis in immunocompromized children and adults [11,40,47] and acute respiratory disease [48]. The prevalence of VA1/HMO-C viruses in stools ranges from 0.2% to 1.6% in diarrheic and non-diarrheic subjects worldwide, with limited detection in Nepal, Japan, Tanzania, The Gambia, France, and the U.K. [9,44,47]. Seroprevalence of VA1/HMO-C has been reported at 65% in adults [49].
2.5. Bastrovirus
The divergent human astrovirus-like virus tentatively named Bastrovirus was isolated from patients in The Netherlands. The capsid region is homologous to the capsid of HAstV whilst the RdRp region is more closely related to members of the Hepeviridae family. This virus remains to be classified by the ICVT, however the capsid regions clusters closest to the MLB1-3 viruses suggesting an evolutionary relationship to these divergent human astroviruses [50] (Figure 1a). Revealing the geographic and host diversity of this virus, divergent Bastrovirus strains have also been isolated from bat, pig, and rat species in Vietnam, forming a distinct cluster of strains that also await classification (Figure 1a).
3. Non-Human MAstV
Mamastroviruses are capable of infecting a wide range of mammalian species including companion animals (cats and dogs), intensively farmed species (pigs and cattle), as well as terrestrial and aquatic wild mammalian species. Unexpectedly there is no clear clustering of viruses separated by hosts of terrestrial or aquatic origins (Figure 1a,b). In addition to the classified MAstV species, there are numerous divergent viruses isolated from diverse species which likely represent new species awaiting formal classification (Figure 1a) [51].
Not surprisingly, domesticated animals such as cats and dogs harbor astrovirus strains more closely related to HAstV than the viruses harboured by many other animal species. Feline astrovirus was first identified in 1981, and feline strains form a small discrete cluster defined as the species MAstV-2, closely related to the human strains comprising the species MAstV-1 [52] (Figure 1a). Based on evolutionary analysis, an interspecies transmission pathway has been hypothesised whereby porcine strains may have been transmitted to cats and subsequently to humans, possibly involving other intermediary species suggesting sustained interspecies transmission events [53]. Characterization of an outbreak of diarrhea in a group of captive cheetahs in a breeding facility identified an astrovirus strain most closely related to feline strains (Figure 1a), however it is not clear if this is a recent transmission from domesticated cats or if the virus is circulating independently in cheetahs [54]. Despite dogs also being a companion animal, canine astrovirus, first described in the 1980s, are not as closely related to human strains as feline strains appear to be, based on phylogenetic analysis (Figure 1a,b). Canine strains form a small, discrete cluster comprising species MAstV-5, within a lineage comprised of dolphin and sea lion clades representing species MAstV-4 and -11 [55,56], and feline, porcine, and human strains are more distantly related within the same lineage (Figure 1a).
A large diverse lineage, largely comprised of unclassified viruses or tentatively classified viruses awaiting approval encompasses divergent marmot and rabbit strains (MAstV-23), small, discrete clusters of porcine/ovine/bovine strains (MAstV-24), divergent rat strains (MAstV-25), and unclassified mouse and mink strains forming a small discrete cluster (Figure 1a). Numerous porcine strains also cluster within this lineage; PAstV comprise Mamastrovirus 3 and were first detected by EM in pigs in the U.K. and the USA [57], and are now recognized to have worldwide distribution [12]. There is a high prevalence of astrovirus detection in pigs, up to 80% in some studies, suggesting that pigs may be persistently infected with various strains of PAstV [58]. There are five recognized lineages (PAstV1–5) reflecting the diversity of strains, suggesting swine are highly permissive to astrovirus infection often without symptoms of disease (Figure 1a,b). This diversity likely reflects the varying origins of these viruses, in particular highlighting frequent interspecies transmission and recombination events [58,59,60]. A study in the USA revealed the high level of co-infections in pigs (13.9%), revealing the frequent opportunity for recombination, especially between viruses of different lineages [61]. The close clustering between some porcine and humans strains, in particular MAstV-3 and MAstV-1 viruses, reflects the close contact between these species with the close sharing of environments and co-housing documented in some countries facilitating frequent interspecies transmission [62]. Wild boars showing no symptoms of enteric disease, housed in a captive breeding park with no contact with domesticated pigs, were found to harbor astrovirus strains with a high degree of genetic similarity to porcine strains comprising MAstV-26 and suggests that the virus may be derived from commonly circulating porcine strains [63] (Figure 1a,b). Porcupine strains isolated in China also cluster with unclassified PAstV-2 strains suggesting further interspecies transmission of these viruses (Figure 1a) [64].
Unexpectedly, limited astrovirus strains have been isolated from sheep. Ovine astrovirus was first identified by EM in 1977 [4], and OAstV-1 clusters with bovine strains comprising MAstV-13 and a second strain from a healthy sheep characterized in 2009, OAstV-2, clusters with porcine and bovine strains comprising MAstV-24 [65] (Figure 1a). Similarly, astroviruses are not associated with a significant burden of diarrheal disease in bovine species. The first bovine astrovirus was detected in England in 1978 [66] and bovine astrovirus strains have been detection in association with neurological disease, including encephalitis (BoAstV-CH13/NeuroS1) [14,15,16,67,68,69] and diarrheal disease in calves in South Korea [70] and cattle and buffalo calves in China [71]. Two serotypes were previously recognized, BoAstV-1 and BoAstV-2 [72], however based on phylogenetic analysis there are multiple lineages of BoAstV strains circulating in farmed bovine populations, and the close clustering of bovine, porcine, and ovine strains in multiple lineages reflects the common interspecies transmission events that occur between farmed animals (Figure 1a,b). Phylogenetic analysis also reveals that bovine-like astrovirus strains have been isolated from numerous wild species including water buffalo, yak, and European roe deer (CcAstV-1 and CcAstV-2) suffering from gastroenteritis [73]. Unclassified astroviruses from dromedary camels (DcAstV) [27] also cluster in a lineage comprised of porcine and bovine strains, further suggesting multiple interspecies transmission events (Figure 1a).
Substantial astrovirus strain diversity has been observed in small mammals, primarily rodents and bats, forming both species-specific clusters reflecting endemic transmission and co-clustering of strains with other host species reflecting widespread interspecies transmission (Figure 1a,b). Novel murine astrovirus strains have been isolated from laboratory mice in the USA and Japan [74,75]. Divergent viruses have also been detected, such as those detected in rats (MAstV-25), highlighting the need for more detailed detection and characterization of these viruses to better understand the role these animals play in astrovirus transmission between varied species. One of the mammalian species with the highest burden of symptomatic astrovirus infection is mink; infection is associated with pre-weaning diarrhea syndrome and the neurological condition “shaking mink syndrome” [17,76]. Mink viruses cluster within multiple lineages suggesting that the species is permissive to infection with multiple, diverse lineages of viruses (Figure 1a). Bat astroviruses were first detected in 2008 in Hong Kong [77] and subsequently detected in bats in China [78,79], North America [80], Germany [81], Hungary [82], and the Czech Republic [83] from numerous bat species, with detection rates ranging from 36% to 100% in Miniopterus magnater bats and from 50% to 70% of Miniopterus pusillus bats sampled in Hong Kong [77]. A diverse population of viruses appears to be highly prevalent in bats without causing disease [12] (Figure 1 and Figure 2). The majority of bat astroviruses are divergent from other characterized mammalian astroviruses, and display a high degree of genetic diversity forming numerous recognized and proposed species (Figure 1a). Some bat sequences clustered with strains from other species including fox, cattle, and mice, suggesting that bats are highly permissive to infection with diverse astrovirus strains from multiple hosts and play a key role in astrovirus diversity and interspecies transmission (Figure 1a,b).
Astroviruses have been detected in aquatic mammalian species including Californian sea lions, Steller sea lion, bottlenose dolphin, killer whale, and minke whale [26]. The strains from these aquatic species do not cluster together, instead forming multiple discrete clusters, suggesting several transmission events from terrestrial mammals to aquatic mammals (Figure 1a). Minke whale and BdAstV-3 strains are divergent to characterized astrovirus strains, with BdAstV-1 strains comprising MAstV-7 and a diverse group of sea lion viruses comprising the MAstV-4 cluster with porcine and canine strains (Figure 1a).
There are numerous divergent strains that are yet to be classified that may reflect interspecies transmission events. A single divergent feline strain along with a fox strain cluster within a diverse lineage comprised of human HMO strains (MAstV-8 and -9), sea lion (MAstV-11), unclassified sea lion, mink (MAstV-21), and bat (MAstV-12) strains. A Himalayan marmot strain clusters with MAstV-6 strains (Figure 1a,b). There appears to be a large diversity of MAstV strains not captured by the currently available sequences, highlighting the need for more detailed sampling and characterization of animal strains.
4. Avastrovirus
The isolation of astroviruses from avian species predates their isolation in humans, with disease in ducklings described in 1965, however the virus was not formally recognized as an astrovirus until 1984 [5,84]. Avian astroviruses have been documented to cause infection in poultry leading to economic losses in farms and affecting food production worldwide [85]. Avian astroviruses have been associated with a spectrum of disease ranging from subclinical infection in seemingly healthy adult birds to heavy flock losses. Pleomorphic symptoms include enteritis in turkeys, chickens, and guineafowl, mild growth depression and nephritis in chickens, and hepatitis in ducklings [86]. Astrovirus infection has been implicated in pre-hatching mortality in ducklings and goslings [87].
In addition to AAstV-1 comprised of TAstV-1, AAstV-2 comprised of ANV1 and ANV2, and AAstV-3 comprised of TAstV-2 and duck astrovirus 1 (DAstV-1), there are numerous yet-to-be classified viruses, including Turkey astrovirus 3 (TAstV-3), chicken astrovirus A (CAstV-A), chicken astrovirus B (CAstV-B) [88,89], GFAstV, and multiple viruses isolated from ducks including duck hepatitis virus 3 (DHV-3/DAstV-2) [88], DHV-3/DAstV-2-like viruses, DAstV-CPH (DAstV-3) [90], DAstV-4, DAstV-YP/DAstV-4-like, and diverse viruses isolated from wild birds [86].
4.1. TAstV
Avian astrovirus have been detected in outbreaks of enteric disease in turkey poults, and avian astrovirus were first reported as an agent of gastroenteritis and mortality in young turkeys in 1980, associated with a condition known as poult enteritis mortality syndrome (PEMS) [91,92]. Subsequently there have been sporadic reports of astrovirus outbreaks in turkeys, associated with enteritis and growth depression [93]. Based on serological and genetic analysis, two types of TAstV have been recognized (TAstV-1 and TAstV-2). TAstV-1 comprises the Avastrovirus 1 species and was first described in the U.K. [91]. TAstV-1 has limited detection in other avian species with sporadic detection in chicken and ducks and forms a discrete cluster in both the capsid and RdRp phylogenetic analysis (Figure 2a,b). TAstV-2 was identified in 1996 [94], and is likely to be classified within the species Avastrovirus 3 and is primarily associated with PEMS [95]. TAstV-2 is the predominant TAstV lineage, with a wider global circulation and greater genetic diversity compared to TAstV-1, reflected by the co-circulation of multiple sub-lineages (Figure 2a,b). Based on phylogenetic analysis of the RdRp gene there appears to be limited interspecies transmission of TAstV-2-like viruses detected in chicken and duck (Figure 2b). Astroviruses infecting guineafowl are closely related to TAstV-2 strains based on analysis of the RdRp region [96], however the capsid region is distinct to TAstV-2 strains, forming a discrete cluster in a lineage of unassigned viruses suggesting they are closely related to CAstV-B strains (Figure 2a). These GFAstVs were possibly derived from recombination and interspecies events followed by sustained transmission in the guineafowl population. This highlights the limitation of classifying viruses based on a single region of the genome.
4.2. CAstV
Chicken astrovirus has been associated with runting-stunting syndrome (RSS) in chickens characterized by poor weight gain, lower feed conversion, and mortality resulting in economic losses [97], and “white chicks” disease associated with increased mortality of embryos and chicks, weakness, and white plumage [98]. Currently two serotypes of CAstV have been described [97], and both serotypes form discrete clusters in the phylogenetic analysis of the capsid region within a lineage of unclassified viruses (likely to be classified within AAstV-1). The CAstV-A viruses form a smaller, discrete lineage, clustering closest to DAstV-2 strains (Figure 2a). CAstV-B strains form a large lineage clustering closest to GFAstV strains. Based on phylogenetic analysis of the small region of the RdRp gene commonly sequenced, it does not allow CAstV-A and -B strains to form discrete clusters as seen in the capsid analysis, possibly reflecting multiple recombination events in CAstV strains (Figure 2b). CAstV strains have limited detection in other avian species with sporadic detection in pigeon and duck (Figure 2a,b).
4.3. ANV
Avian nephritis virus was identified in association with intestinal nephritis in chickens and growth retardation. The first serotype of ANV (ANV-1) was isolated from a healthy broiler chick in 1976 [99] and was initially regarded as a picornavirus, subsequently reclassified within the Astroviridae family in 2000 [100]. Based on capsid phylogenetic analysis, ANV-1 strains cluster within the AAstV-2 lineage forming a small, discrete, relatively conserved cluster (Figure 2a). The second serotype (ANV-2) was later described from chicks with stunted growth [101]. Based on capsid phylogenetic analysis, ANV-2 strains cluster within the AAstV-2 lineage, forming a larger, more diverse sub-lineage compared to ANV-1. ANV-1 and ANV-2 viruses have also been sporadically detected in ducks and turkeys (Figure 2a). A third serotype (ANV-3) detected in chickens and turkeys with RSS and locomotion impairment has been proposed based on sequencing of the ORF1a region, however complete genome sequences are unavailable for adequate comparisons to the recognized serotypes [102]. The virus designated pigeon ANV (P-ANV) was detected during an outbreak of gastrointestinal illness in young pigeons in China [103]. P-ANV represents an interspecies transmission event from chickens to pigeon; based on phylogenetic analysis, P-ANV strains share a high degree of genetic similarity to ANV-2 strains and cluster closely with strains also circulating in China suggesting a localized transmission event, and these viruses should be considered as ANV-2 viruses and do not require a distinct designation (Figure 2a) [104]. Based on phylogenetic analysis of the small region of the RdRp gene commonly sequenced, it does not allow ANV 1 and 2 strains to form discrete clusters as characterized in the capsid analysis (Figure 2b). Whilst this may reflect common recombination events between ANV strains, the analysis of a small region does not adequately allow for distinct clustering.
4.4. DAstV
Astrovirus infection in ducks has been associated with a highly contagious and fatal hepatitis, historically known as duck hepatitis virus type 2 (DHV-2), which was described in the U.K. and subsequently serotype DHV-3 was isolated in the USA [5,84,105,106]. The 9th ICTV report classified DHV-2 and DHV-3 as DAstV-1 and DAstV-2, respectively [25]. DAstV-1 strains form a small, discrete cluster within the AAstV-3 lineage based on phylogenetic analysis of the capsid region, clustering closest to TAstV-2 strains (Figure 2a). DAstV-2 viruses, currently unclassified, form a discrete, highly conserved cluster closest to CAstV-A strains (Figure 2a). Based on phylogenetic analysis of the RdRp region, DAstV-1 is closely related to other unclassified duck strains whilst DAstV-2 strains are more closely related to TAstV-2 strains (Figure 2b). Further unclassified serotypes have been described; DAstV-3-CPH, DAstV/C-NGB, and DAstV-4 [107]. Duck astrovirus DAstV-3/CPH has been suggested to transmit horizontally and vertically [108] and DAstV-3/CPH viruses have also been detected in goslings [107]. Phylogenetic analysis of the RdRp region highlights the diversity of AAstV strains circulating in duck species (Figure 2b). Strains endemic to ducks have limited detection in other avian species including geese (Figure 2a,b).
4.5. Novel Astroviruses Detected in Wild Birds
Astroviruses have been detected in numerous wild aquatic species including teals, pintails, and shovelers (belonging to the order Anseriformes), sanderlings (order Charadriiformes), and herons and spoonbills (order Pelecaniformes) [86,109]. Fewer land dwelling wild birds have been found to harbor avian astroviruses including doves and pigeons (order Colombiformes), European roller (order Coraciiformes), and black-naped monarch (order Passeriformes) [51,104,110,111]. These viruses are highly divergent and largely unclassified, suggesting these viruses are endemic to the wild bird population (Figure 2a,b). The migratory behavior of some of these species provides ideal conditions for virus dissemination and diversification as during migration there is a high degree of co-mingling and increased density of birds of different species that may originate from varied geographic regions [112].
5. Interspecies Transmission
Phylogenies indicate that the genetic diversity of mamastroviruses has been shaped by extensive interspecies transmission events that have occurred in the past between wild and domestic species and humans. However, the inference of astrovirus interspecies transmission events is hampered due to (a) sequencing of only a small portion of the genome; (b) inadequate sampling; or (c) that the event occurred early during the divergence of astrovirus lineages. A few exceptions where host-jumps are apparent have been in livestock where a greater level of sampling has occurred. This includes hosts with a greater level of interaction (e.g., MAstV-13 in ovine and bovine) or host genetic similarly (e.g., MAstV-26 in wild boars and domestic pigs). With the exception of HAstV-4 strains associated with fatal meningoencephalitis, it is interesting to note that viruses from multiple species, all recognized to cause neurological symptoms, are closely related including human VA1/HMO-C viruses and mink and bovine viruses also associated with neurological symptoms, suggesting that these related viruses may have a distinct phenotype compared to other MAstV strains (Figure 1a,b). Astrovirus strains identified from fecal samples of multiple non-human primate species from wild, captive, and peri-urban environments in Bangladesh and Cambodia reveal multiple interspecies transmission events, with viruses closely related to the VA/HMO lineage of human viruses, and non-human mammalian and avian astroviruses (Figure 1a,b) [28].
Similarly, there appears to be evidence for a high degree of cross species transmission of avian astroviruses between farmed poultry species as described in the above sections. There also appears to be transmission between avian and mammalian species. The highly divergent strains isolated from European roller (Er/SZAL6/HUN/2011 and Er/BMTK529/HUN/2011) exhibited low identity to avian and mammalian astroviruses, cluster within the MAstV lineage, and were likely derived from multiple recombination and interspecies transmission events [51]. The carnivorous diet of this avian species may have facilitated the interspecies transmission event between a rodent or small mammal species. This is the only report of a Mamastrovirus strain in an avian species; in contrast, there have been more reports of Avastrovirus strains detected in mammalian species which may reflect greater sampling density of the mammalian population. A highly divergent group of mink strains detected in China represent a novel clade of astroviruses that were distantly related to previously described mink astrovirus and were closely related to chicken and turkey astroviruses (Figure 2a,b) [113]. These viruses are recombinant strains with the capsid region clustering with CAstV-B strains and the RdRp region clustering with human MAstV strains and CAstV strains (Figure 2a,b). There have been two Avastrovirus strains detected in humans; a strain clustering close to ANV-1 strains detected in turkey and chicken was isolated from a child in The Gambia, and a strain clustering with ANV-2 strains detected in chicken was isolated from a child in Kenya [44]. Serological studies have been used to screen human sera from poultry workers for antibodies to TAstV-2, with up to 26% of participants positive with the highest detection in abattoir workers and turkey growers [114], suggesting avian strains may be readily transmitted to humans under prolonged close contact.
The important role that ecotones play in astrovirus cross-species transmission has been proposed [112]. Ecotones are ecological transition areas such as small and medium sized farms which rear multiple species. The co-rearing of poultry such as domestic ducks, chickens, turkey, and guineafowl can facilitate transmission between these species but also transmission to wild birds [112]. Farms and abattoirs have also been recognized as environments facilitating transmission between livestock species and to farm and abattoir workers [114]. Many other species have contact with livestock in a farming environment; in addition to wild species, companion animals such as cats and dogs and other peri-domestic animals have contact with livestock and their biological waste providing substantial opportunities for cross-species transmission. Astroviruses also persist in bodies of water making the aquatic environment ideal for the transmission of viruses infecting avian species, aquatic mammalian species, and possible transmission between terrestrial and aquatic species. Untreated or inadequately treated sewage and waste water from domestic and farmed areas can reach fresh and marine bodies of water transmitting human and animal viruses. Astroviruses have been detected in the environment and the durability of the virus in this environment may greatly contribute to cross-species transmission within and between terrestrial and aquatic species, generating significant diversity [115].
6. Recombination
In addition to interspecies transmission which generates significant diversity in astrovirus species, both intra-species and inter-species recombination can rapidly generate novel, divergent viruses. Full- and partial-genome sequence analysis has identified multiple strains that have undergone recombination events, which are predominately located within the ORF1b/ORF2 junction region of the genome, which is a region with an RNA secondary structure predicted to contain a stable hairpin structure [21,34,116,117,118,119]. A virus with a recombination event within the ORF1a region has been identified [119]. Some recombinants appear to be highly stable and show widespread detection [21], whilst others are detected sporadically as single strains. Numerous human recombinant strains have been reported between HAstV strains, including strains with HAstV-1 (ORF1b) and HAstV-4 (ORF2) parental viruses, HAstV-1 (ORF1b) and HAstV-3 (ORF2) parental viruses, HAstV-3 (ORF1b) and HAstV-2 (ORF2), and VA2 (ORF1b) and MLB1 (ORF2) parental viruses [116,119,120,121].
Divergent species often represent recombination events between strains of different species. In 2010, the study conducted by Rivera et al. suggested the possibility of a recombination event between human and California sea lion astrovirus strains [26]. A recombinant strain derived from porcine astrovirus and human HAstV-3 strains was reported from piglets and children from various regions of Colombia [62]. ANV strains that appear to be derived from recombination events between ANV-1 (ORF1b) and ANV-2 (ORF2) have been described in the USA [85].
7. Future Considerations for Classification
The increased sampling density of numerous host species combined with the more prevalent use of NGS technologies and viral metagenomic studies will increase the detection of novel strains, further driving the need for a unified, complex, and encompassing classification system. The previously common practice of sequencing a relatively conserved RdRp amplicon of 300–400 bp renders many sequences available in GenBank of little use in detailed phylogenetic analysis, as does the frequent missing metadata regarding species of isolation, country, and date of collection. Sequencing small regions is not adequate to determine if a virus strain is a novel, divergent strain or a recombinant virus. Recommendations should be made to encourage full genome sequencing where possible and the deposition of associated host and demographic information. Although analysis of amino acid sequences of the capsid region is required for classification, it leads to confusion regarding appropriate phylogenetic analysis, with highly inconsistent publication of nucleotide and amino acid trees further complicating attempts to clarify diversity and classification. The topologies of amino acid trees differ to those of nucleotide trees, particularly for the analysis of MAstV, and whilst amino acid trees are required for classification, nucleotide trees may be more appropriate for describing within species diversity (Figures S1 and S2). Incorporating a standardized nomenclature to aid in classification has proven invaluable in the classification of numerous viruses, including rotavirus and influenza [122]. Adopting a nomenclature that records the appropriate metadata associated with sample collection including host, location, date of collection, and determined species and serotype as proposed by Martella and colleagues would vastly improve the usability of strains for more complex analyses [38].
Supplementary Materials
The following are available online at www.mdpi.com/1999-4915/9/5/102/s1, Figure S1: Maximum-likelihood phylogenetic tree of MAstV (a) capsid nucleotide and (b) capsid amino acid sequences, Figure S2: Maximum-likelihood phylogenetic tree of MAstV RdRp, Figure S3: Maximum-likelihood phylogenetic tree of AAstV (a) capsid nucleotide and (b) capsid amino acid sequences, Figure S4: Maximum-likelihood phylogenetic tree of AAstV RdRp.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Madeley, C.R.; Cosgrove, B.P. 28 nm Particles in fæces in infantile gastroenteritis. Lancet 1975, 306, 451–452. [Google Scholar] [CrossRef]
- Risco, C.; Carrascosa, J.L.; Pedregosa, A.M.; Humphrey, C.D.; Sanchez-Fauquier, A. Ultrastructure of human astrovirus serotype 2. J. Gen. Virol. 1995, 76, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Appleton, H.; Higgins, P.G. Letter: Viruses and gastroenteritis in infants. Lancet 1975, 1, 1297. [Google Scholar] [CrossRef]
- Snodgrass, D.R.; Gray, E.W. Detection and transmission of 30 nm virus particles (astroviruses) in faeces of lambs with diarrhoea. Arch. Virol. 1977, 55, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Asplin, F.D. Duck hepatitis: Vaccination against two serological types. Vet. Rec. 1965, 77, 1529. [Google Scholar] [PubMed]
- Jeong, H.S.; Jeong, A.; Cheon, D.S. Epidemiology of astrovirus infection in children. Korean J. Pediatr. 2012, 55, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Glass, R.I.; Noel, J.; Mitchell, D.; Herrmann, J.E.; Blacklow, N.R.; Pickering, L.K.; Dennehy, P.; Ruiz-Palacios, G.; de Guerrero, M.L.; Monroe, S.S. The changing epidemiology of astrovirus-associated gastroenteritis: A review. Arch. Virol. Suppl. 1996, 12, 287–300. [Google Scholar] [PubMed]
- Mitchell, D.K.; Matson, D.O.; Jiang, X.; Berke, T.; Monroe, S.S.; Carter, M.J.; Willcocks, M.M.; Pickering, L.K. Molecular epidemiology of childhood astrovirus infection in child care centers. J. Infect. Dis. 1999, 180, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Li, L.; Victoria, J.; Oderinde, B.; Mason, C.; Pandey, P.; Zaidi, S.Z.; Delwart, E. Multiple novel astrovirus species in human stool. J. Gen. Virol. 2009, 90, 2965–2972. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M.; et al. Astrovirus MLB2, a new gastroenteric virus associated with meningitis and disseminated infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Schultz-Cherry, S. Pathogenesis of astrovirus infection. Viral Immunol. 2005, 18, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Bouzalas, I.G.; Wuthrich, D.; Walland, J.; Drogemuller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Hoper, D.; Beer, M.; Hoffmann, B. Detection of a novel bovine astrovirus in a cow with encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Blomstrom, A.L.; Widen, F.; Hammer, A.S.; Belak, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pintó, R.M.; Guix, S. Human Astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Atkins, J.F. Candidates in astroviruses, seadornaviruses, cytorhabdoviruses and coronaviruses for +1 frame overlapping genes accessed by leaky scanning. Virol. J. 2010, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Monroe, S.S.; Koonin, E.V.; Stine, S.E.; Glass, R.I. RNA sequence of astrovirus: Distinctive genomic organization and a putative retrovirus-like ribosomal frameshifting signal that directs the viral replicase synthesis. Proc. Natl. Acad. Sci. USA 1993, 90, 10539–10543. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.E.; Mitchell, D.K.; Guerrero, M.L.; Berke, T.; Matson, D.O.; Monroe, S.S.; Pickering, L.K.; Ruiz-Palacios, G. Molecular epidemiology of human astrovirus diarrhea among children from a periurban community of Mexico City. J. Infect. Dis. 2001, 183, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.H.; Kakizawa, J.; Wen, L.Y.; Shimizu, M.; Nishio, O.; Fang, Z.Y.; Ushijima, H. Genetic analysis of the capsid region of astroviruses. J. Med. Virol. 2001, 64, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Dong, L.; Mendez, E.; Tao, Y. Crystal structure of the human astrovirus capsid spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, T. Novel human astroviruses: Challenges for developing countries. VirusDisease 2014, 25, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Guix, S.; Krishna, N.K.; Méndez, E.; Monroe, S.S.; Pantin-Jackwood, M.; Schultz-Cherry, S. Family—Astroviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 953–959. [Google Scholar]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.D.; Wellehan, J.F., Jr. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, S.; Xie, J.; Jose, S.; Fan, R.Y.; Wernery, U.; Yuen, K.Y. A novel astrovirus from dromedaries in the Middle East. J. Gen. Virol. 2015, 96, 2697–2707. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, F.A.T.; San, S.; Hasan, M.K.; Wang, D.; Jones-Engel, L.; Schultz-Cherry, S. Non-human Primates harbor diverse mammalian and avian astroviruses including those associated with human infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.A.; Hoang, L.; Pham le, D.; Hoang, K.T.; Mizuguchi, M.; Okitsu, S.; Ushijima, H. Identification of human astrovirus infections among children with acute gastroenteritis in the southern part of Vietnam during 2005–2006. J. Med. Virol. 2008, 80, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Wunderli, W.; Meerbach, A.; Guengoer, T.; Berger, C.; Greiner, O.; Caduff, R.; Trkola, A.; Bossart, W.; Gerlach, D.; Schibler, M.; et al. Astrovirus infection in hospitalized infants with severe combined immunodeficiency after allogeneic hematopoietic stem cell transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, S.; Angez, M.; Alam, M.M.; Jebbink, M.F.; Deijs, M.; Canuti, M.; Sharif, S.; de Vries, M.; Khurshid, A.; Mahmood, T.; et al. Identification and characterization of unrecognized viruses in stool samples of non-polio acute flaccid paralysis children by simplified VIDISCA. Virol. J. 2014, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.B.; Lee, T.W. Human astrovirus serotypes. Lancet 1984, 2, 1405. [Google Scholar] [CrossRef]
- Herrmann, J.E.; Hudson, R.W.; Perron-Henry, D.M.; Kurtz, J.B.; Blacklow, N.R. Antigenic characterization of cell-cultivated astrovirus serotypes and development of astrovirus-specific monoclonal antibodies. J. Infect. Dis. 1988, 158, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Belliot, G.; Laveran, H.; Monroe, S.S. Detection and genetic differentiation of human astroviruses: Phylogenetic grouping varies by coding region. Arch. Virol. 1997, 142, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Gabbay, Y.B.; Leite, J.P.; Oliveira, D.S.; Nakamura, L.S.; Nunes, M.R.; Mascarenhas, J.D.; Heinemann, M.B.; Linhares, A.C. Molecular epidemiology of astrovirus type 1 in Belem, Brazil, as an agent of infantile gastroenteritis, over a period of 18 years (1982–2000): Identification of two possible new lineages. Virus Res. 2007, 129, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Caballero, S.; Villena, C.; Bartolome, R.; Latorre, C.; Rabella, N.; Simo, M.; Bosch, A.; Pinto, R.M. Molecular epidemiology of astrovirus infection in Barcelona, Spain. J. Clin. Microbiol. 2002, 40, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.A.; Cardoso, D.D.; Schreier, E. Molecular characterization of human astroviruses isolated in Brazil, including the complete sequences of astrovirus genotypes 4 and 5. Arch. Virol. 2006, 151, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Pinto, P.; Tummolo, F.; De Grazia, S.; Giammanco, G.M.; Medici, M.C.; Ganesh, B.; L’Homme, Y.; Farkas, T.; Jakab, F.; et al. Analysis of the ORF2 of human astroviruses reveals lineage diversification, recombination and rearrangement and provides the basis for a novel sub-classification system. Arch. Virol. 2014, 159, 3185–3196. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Kirkwood, C.D.; Wang, D. Complete genome sequence of a highly divergent astrovirus isolated from a child with acute diarrhea. Virol. J. 2008, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.-L.; Cordey, S.; Brito, F.; Kaiser, L. Novel human astroviruses: Novel human diseases? J. Clin. Virol. 2016, 82, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Holtz, L.R.; Bauer, I.K.; Jiang, H.; Belshe, R.; Freiden, P.; Schultz-Cherry, S.L.; Wang, D. Seroepidemiology of astrovirus MLB1. Clin. Vaccine Immunol. 2014, 21, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Holtz, L.R.; Jiang, Y.; Rajendran, P.; Franz, C.J.; Zhao, G.; Kang, G.; Wang, D. Human stool contains a previously unrecognized diversity of novel astroviruses. Virol. J. 2009, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Holtz, L.R.; Bauer, I.; Franz, C.J.; Zhao, G.; Bodhidatta, L.; Shrestha, S.K.; Kang, G.; Wang, D. Comparison of novel MLB-clade, VA-clade and classic human astroviruses highlights constrained evolution of the classic human astrovirus nonstructural genes. Virology 2013, 436, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.T.; Bauer, I.K.; Antonio, M.; Adeyemi, M.; Saha, D.; Oundo, J.O.; Ochieng, J.B.; Omore, R.; Stine, O.C.; Wang, D.; et al. Prevalence of classic, MLB-clade and VA-clade Astroviruses in Kenya and The Gambia. Virol. J. 2015, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Nordgren, J.; Ouermi, D.; Simpore, J.; Nitiema, L.W.; Deng, X.; Delwart, E. New astrovirus in human feces from Burkina Faso. J. Clin. Virol. 2014, 60, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Li, Y.; Ruone, S.; Conrardy, C.; Gregoricus, N.; Toney, D.; Virgin, H.W.; Anderson, L.J.; Vinje, J.; Wang, D.; Tong, S. Identification of a novel astrovirus (astrovirus VA1) associated with an outbreak of acute gastroenteritis. J. Virol. 2009, 83, 10836–10839. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.; Anderson, G.; Virasami, A.; et al. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Brito, F.; Vu, D.L.; Turin, L.; Kilowoko, M.; Kyungu, E.; Genton, B.; Zdobnov, E.M.; D’Acremont, V.; Kaiser, L. Astrovirus VA1 identified by next-generation sequencing in a nasopharyngeal specimen of a febrile Tanzanian child with acute respiratory disease of unknown etiology. Emerg. Microbes Infect. 2016, 5, e67. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Ching, K.H.; Esper, F.; Iadarola, M.J.; Delwart, E.; Lipkin, W.I.; Kapoor, A. Serological studies confirm the novel astrovirus HMOAstV-C as a highly prevalent human infectious agent. PLoS ONE 2011, 6, e22576. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Cotten, M.; Canuti, M.; Deijs, M.; Jebbink, M.F.; van Hemert, F.J.; Phan, M.V.T.; Bakker, M.; Jazaeri Farsani, S.M.; Kellam, P.; et al. A novel astrovirus-like RNA virus detected in human stool. Virus Evol. 2016, 2, vew005. [Google Scholar] [CrossRef] [PubMed]
- Pankovics, P.; Boros, A.; Kiss, T.; Delwart, E.; Reuter, G. Detection of a mammalian-like astrovirus in bird, European roller (Coracias garrulus). Infect. Genet. Evol. 2015, 34, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, Y.; Zimmer, J.F.; Moise, N.S.; Scott, F.W. Detection of astroviruses in feces of a cat with diarrhea. Brief report. Arch. Virol. 1981, 70, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Lukashov, V.V.; Goudsmit, J. Evolutionary relationships among Astroviridae. J. Gen. Virol. 2002, 83, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Atkins, A.; Wellehan, J.F.; Childress, A.L.; Archer, L.L.; Fraser, W.A.; Citino, S.B. Characterization of an outbreak of astroviral diarrhea in a group of cheetahs (Acinonyx jubatus). Vet. Microbiol. 2009, 136, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Moschidou, P.; Catella, C.; Larocca, V.; Pinto, P.; Losurdo, M.; Corrente, M.; Lorusso, E.; Banyai, K.; Decaro, N.; et al. Enteric disease in dogs naturally infected by a novel canine astrovirus. J. Clin. Microbiol. 2012, 50, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Williams, F.P., Jr. Astrovirus-like, coronavirus-like, and parvovirus-like particles detected in the diarrheal stools of beagle pups. Arch. Virol. 1980, 66, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhoea. Vet. Rec. 1980, 107, 532–533. [Google Scholar] [PubMed]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Wang, C.; Tong, W.; Zheng, H.; Hua, X.; Yang, S.; Guo, Y.; Zhang, W.; Tong, G. Complete genome of a novel porcine astrovirus. J. Virol. 2012, 86, 13820–13821. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Complete genome sequence of a newly identified porcine astrovirus genotype 3 strain US-MO123. J. Virol. 2012, 86, 13126. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.T.; Gimenez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, J.C.; Gutierrez, M.F. Genomic analysis of two ORF2 segments of new porcine astrovirus isolates and their close relationship with human astroviruses. Can. J. Microbiol. 2010, 56, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Nemes, C.; Boros, A.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Astrovirus in wild boars (Sus scrofa) in Hungary. Arch. Virol. 2012, 157, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chmura, A.A.; Li, J.; Zhu, G.; Desmond, J.S.; Zhang, Y.; Zhang, W.; Epstein, J.H.; Daszak, P.; Shi, Z. Detection of diverse novel astroviruses from small mammals in China. J. Gen. Virol. 2014, 95, 2442–2449. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Delwart, E.; Boros, A. Identification of a novel astrovirus in domestic sheep in Hungary. Arch. Virol. 2012, 157, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med. Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Wuthrich, D.; Boujon, C.L.; Truchet, L.; Selimovic-Hamza, S.; Oevermann, A.; Bouzalas, I.G.; Bruggmann, R.; Seuberlich, T. Exploring the virome of cattle with non-suppurative encephalitis of unknown etiology by metagenomics. Virology 2016, 493, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Seuberlich, T.; Wuthrich, D.; Selimovic-Hamza, S.; Drogemuller, C.; Oevermann, A.; Bruggmann, R.; Bouzalas, I. Identification of a second encephalitis-associated astrovirus in cattle. Emerg. Microbes Infect. 2016, 5, e5. [Google Scholar] [CrossRef] [PubMed]
- Selimovic-Hamza, S.; Bouzalas, I.G.; Vandevelde, M.; Oevermann, A.; Seuberlich, T. Detection of Astrovirus in Historical Cases of European Sporadic Bovine Encephalitis, Switzerland 1958–1976. Front. Vet. Sci. 2016, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Oem, J.K.; An, D.J. Phylogenetic analysis of bovine astrovirus in Korean cattle. Virus Genes 2014, 48, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Alfred, N.; Liu, H.; Li, M.L.; Hong, S.F.; Tang, H.B.; Wei, Z.Z.; Chen, Y.; Li, F.K.; Zhong, Y.Z.; Huang, W.J. Molecular epidemiology and phylogenetic analysis of diverse bovine astroviruses associated with diarrhea in cattle and water buffalo calves in China. J. Vet. Med. Sci. 2015, 77, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Woode, G.N.; Gourley, N.E.; Pohlenz, J.F.; Liebler, E.M.; Mathews, S.L.; Hutchinson, M.P. Serotypes of bovine astrovirus. J. Clin. Microbiol. 1985, 22, 668–670. [Google Scholar] [PubMed]
- Smits, S.L.; van Leeuwen, M.; Kuiken, T.; Hammer, A.S.; Simon, J.H.; Osterhaus, A.D. Identification and characterization of deer astroviruses. J. Gen. Virol. 2010, 91, 2719–2722. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.; Kondov, N.O.; Hayashimoto, N.; Uchida, R.; Cha, Y.; Beyer, A.I.; Wong, W.; Pesavento, P.A.; Suemizu, H.; Muench, M.O.; et al. Identification of an astrovirus commonly infecting laboratory mice in the US and Japan. PLoS ONE 2013, 8, e66937. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, C.C.; Loh, J.; Zhao, G.; Stappenbeck, T.S.; Wang, D.; Huang, H.V.; Virgin, H.W.; Thackray, L.B. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef] [PubMed]
- Englund, L.; Chriel, M.; Dietz, H.H.; Hedlund, K.O. Astrovirus epidemiologically linked to pre-weaning diarrhoea in mink. Vet. Microbiol. 2002, 85, 1–11. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.C.; Chu, D.K.; Liu, W.; Dong, B.Q.; Zhang, S.Y.; Zhang, J.X.; Li, L.F.; Vijaykrishna, D.; Smith, G.J.; Chen, H.L.; et al. Detection of diverse astroviruses from bats in China. J. Gen. Virol. 2009, 90, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Li, J.; Hu, G.; Chen, Z.; Wu, Y.; Chen, Y.; Chen, Z.; Liao, Y.; Zhou, J.; Ke, X.; et al. Isolation and phylogenetic characterization of bat astroviruses in southern China. Arch. Virol. 2011, 156, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Victoria, J.G.; Wang, C.; Jones, M.; Fellers, G.M.; Kunz, T.H.; Delwart, E. Bat guano virome: Predominance of dietary viruses from insects and plants plus novel mammalian viruses. J. Virol. 2010, 84, 6955–6965. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Seebens, A.; Muller, M.A.; Drosten, C. Amplification of emerging viruses in a bat colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Gorfol, T.; Boldogh, S.; Estok, P.; Kurucz, K.; Oldal, M.; Nemeth, V.; Madai, M.; Banyai, K.; et al. Novel European lineages of bat astroviruses identified in Hungary. Acta Virol. 2014, 58, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Dufkova, L.; Strakova, P.; Sirmarova, J.; Salat, J.; Moutelikova, R.; Chrudimsky, T.; Bartonicka, T.; Nowotny, N.; Ruzek, D. Detection of diverse novel bat astrovirus sequences in the Czech Republic. Vector Borne Zoonotic Dis. 2015, 15, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Gough, R.E.; Collins, M.S.; Borland, E.; Keymer, L.F. Astrovirus-like particles associated with hepatitis in ducklings. Vet. Rec. 1984, 114, 279. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Strother, K.O.; Mundt, E.; Zsak, L.; Day, J.M.; Spackman, E. Molecular characterization of avian astroviruses. Arch. Virol. 2011, 156, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Leung, C.Y.; Perera, H.K.; Ng, E.M.; Gilbert, M.; Joyner, P.H.; Grioni, A.; Ades, G.; Guan, Y.; Peiris, J.S.; et al. A novel group of avian astroviruses in wild aquatic birds. J. Virol. 2012, 86, 13772–13778. [Google Scholar] [CrossRef] [PubMed]
- Bidin, M.; Lojkic, I.; Tisljar, M.; Bidin, Z.; Majnaric, D. Astroviruses associated with stunting and pre-hatching mortality in duck and goose embryos. Avian Pathol. 2012, 41, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Wilkinson, D.S.; Jewhurst, H.L.; Wylie, M.; Gordon, A.W.; Adair, B.M. A seroprevalence investigation of chicken astrovirus infections. Avian Pathol. 2009, 38, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.I.; Icard, A.H.; Linnemann, E.; Sellers, H.S.; Mundt, E. Determination of the full length sequence of a chicken astrovirus suggests a different replication mechanism. Virus Genes 2012, 44, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, F.; Zhang, D. Complete sequence of a novel duck astrovirus. Arch. Virol. 2014, 159, 2823–2827. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S.; Curran, W.L.; McFerran, J.B. Detection of astroviruses in turkey faeces by direct electron microscopy. Vet. Rec. 1980, 106, 561. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.A.; Saif, Y.M.; Heggen-Peay, C.L.; Edens, F.W.; Havenstein, G.B. Induction of functional defects in macrophages by a poult enteritis and mortality syndrome-associated turkey astrovirus. Avian Dis. 2001, 45, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.L.; Saif, Y.M. Astrovirus: A cause of an enteric disease in turkey poults. Avian Dis. 1986, 30, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Seal, B.S.; Schultz-Cherry, S. Molecular characterization of an avian astrovirus. J. Virol. 2000, 74, 6173–6177. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Day, J.M.; Jackwood, M.W.; Spackman, E. Enteric viruses detected by molecular methods in commercial chicken and turkey flocks in the United States between 2005 and 2006. Avian Dis. 2008, 52, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Cattoli, G.; De Battisti, C.; Toffan, A.; Salviato, A.; Lavazza, A.; Cerioli, M.; Capua, I. Co-circulation of distinct genetic lineages of astroviruses in turkeys and guinea fowl. Arch. Virol. 2007, 152, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Baxendale, W.; Mebatsion, T. The isolation and characterisation of astroviruses from chickens. Avian Pathol. 2004, 33, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Sajewicz-Krukowska, J.; Domanska-Blicharz, K. Nearly full-length genome sequence of a novel astrovirus isolated from chickens with ‘white chicks’ condition. Arch. Virol. 2016, 161, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Imada, T.; Kawamura, H. Characterization of a picornavirus isolated from broiler chicks. Avian Dis. 1979, 23, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Imada, T.; Yamaguchi, S.; Mase, M.; Tsukamoto, K.; Kubo, M.; Morooka, A. Avian nephritis virus (ANV) as a new member of the family Astroviridae and construction of infectious ANV cDNA. J. Virol. 2000, 74, 8487–8493. [Google Scholar] [CrossRef] [PubMed]
- Takase, K.; Shinohara, K.; Tsuneyoshi, M.; Yamamoto, M.; Yamada, S. Isolation and characterisation of cytopathic avian enteroviruses from broiler chicks. Avian Pathol. 1989, 18, 631–642. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; Dam, G.B.; de Laar, J.M.; Biermann, Y.; Verstegen, I.; Edens, F.; Schrier, C.C. Detection and characterization of a new astrovirus in chicken and turkeys with enteric and locomotion disorders. Avian Pathol. 2011, 40, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhu, A.L.; Yuan, C.L.; Yu, Y.; Zhu, C.X.; Lan, D.L.; Yang, Z.B.; Cui, L.; Hua, X.G. Detection of astrovirus infection in pigeons (Columbia livia) during an outbreak of diarrhoea. Avian Pathol. 2011, 40, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhu, A.L.; Yu, Y.; Yuan, C.L.; Zhu, C.X.; Yang, Z.B.; Cui, L.; Hua, X.G. Complete sequence and genetic characterization of pigeon avian nephritis virus, a member of the family Astroviridae. Arch. Virol. 2011, 156, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Pan, M.; Wang, X.; Xu, Y.; Xie, X.; Knowles, N.J.; Yang, H.; Zhang, D. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J. Gen. Virol. 2009, 90, 1104–1108. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.A.; Calnek, B.W. In Vitro isolation, propagation, and characterization of duck hepatitis virus type III. Avian Dis. 1979, 23, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Liu, N.; Wang, X.; Wang, F.; Zhang, D. Genetic characterization of a novel astrovirus in Pekin ducks. Infect. Genet. Evol. 2015, 32, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Jiang, M.; Wang, M.; Wang, F.; Zhang, B.; Zhang, D. Isolation and detection of duck astrovirus CPH: Implications for epidemiology and pathogenicity. Avian Pathol. 2016, 45, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Honkavuori, K.S.; Briese, T.; Krauss, S.; Sanchez, M.D.; Jain, K.; Hutchison, S.K.; Webster, R.G.; Lipkin, W.I. Novel coronavirus and astrovirus in Delaware Bay shorebirds. PLoS ONE 2014, 9, e93395. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Yaung, K.N.; Joyner, P.H.; Keatts, L.; Borthwick, S.; Neves, E.S.; San, S.; Gilbert, M.; Smith, G.J. Detection of a novel astrovirus from a black-naped monarch (Hypothymis azurea) in Cambodia. Virol. J. 2015, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Kofstad, T.; Jonassen, C.M. Screening of feral and wood pigeons for viruses harbouring a conserved mobile viral element: Characterization of novel astroviruses and picornaviruses. PLoS ONE 2011, 6, e25964. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Yang, Y.; Wang, G.S.; Shao, X.Q.; Zhang, S.Q.; Wang, F.X.; Tan, B.; Tian, F.L.; Cheng, S.P.; Wen, Y.J. Detection and characterization of avastrovirus associated with diarrhea isolated from minks in China. Food Environ. Virol. 2014, 6, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Kayali, G.; Burnham, A.; Oshansky, C.M.; Thomas, P.G.; Gray, G.C.; Beck, M.A.; Schultz-Cherry, S. Detection of antibodies against turkey astrovirus in humans. PLoS ONE 2014, 9, e96934. [Google Scholar] [CrossRef] [PubMed]
- Morsy El-Senousy, W.; Guix, S.; Abid, I.; Pinto, R.M.; Bosch, A. Removal of astrovirus from water and sewage treatment plants, evaluated by a competitive reverse transcription-PCR. Appl. Environ. Microbiol. 2007, 73, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Babkin, I.V.; Tikunov, A.Y.; Zhirakovskaia, E.V.; Netesov, S.V.; Tikunova, N.V. High evolutionary rate of human astrovirus. Infect. Genet. Evol. 2012, 12, 435–442. [Google Scholar] [CrossRef] [PubMed]
- De Grazia, S.; Martella, V.; Chironna, M.; Bonura, F.; Tummolo, F.; Calderaro, A.; Moschidou, P.; Giammanco, G.M.; Medici, M.C. Nationwide surveillance study of human astrovirus infections in an Italian paediatric population. Epidemiol. Infect. 2013, 141, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Jonassen, T.O.; Monceyron, C.; Lee, T.W.; Kurtz, J.B.; Grinde, B. Detection of all serotypes of human astrovirus by the polymerase chain reaction. J. Virol. Methods 1995, 52, 327–334. [Google Scholar] [CrossRef]
- Wolfaardt, M.; Kiulia, N.M.; Mwenda, J.M.; Taylor, M.B. Evidence of a recombinant wild-type human astrovirus strain from a Kenyan child with gastroenteritis. J. Clin. Microbiol. 2011, 49, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Medici, M.C.; Terio, V.; Catella, C.; Bozzo, G.; Tummolo, F.; Calderaro, A.; Bonura, F.; Di Franco, M.; Banyai, K.; et al. Lineage diversification and recombination in type-4 human astroviruses. Infect. Genet. Evol. 2013, 20, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Sebeny, P.J.; Klena, J.D.; Pimentel, G.; Mansour, A.; Naguib, A.M.; Bruton, J.; Young, S.Y.; Holtz, L.R.; Wang, D. Novel astroviruses in children, Egypt. Emerg. Infect. Dis. 2011, 17, 2391–2393. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Banyai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Maximum-likelihood phylogenetic tree of Mamastrovirus (MAstV) capsid (a) and RNA-dependent RNA polymerase (RdRp) (b) genes. Trees were generated from nucleotide sequences using the maximum-likelihood method with the general time reversible nucleotide substitution model with gamma distribution (GTRG+G) and 1000 bootstrap replicates and nodes with bootstrap support values ≥70 are shown by an asterisk. Proposed species yet to be recognized are designated with a ^ symbol. Strains are colored by host and trees with full taxa names are provided as Supplementary Materials (Figures S1 and S2).
Figure 1.
Maximum-likelihood phylogenetic tree of Mamastrovirus (MAstV) capsid (a) and RNA-dependent RNA polymerase (RdRp) (b) genes. Trees were generated from nucleotide sequences using the maximum-likelihood method with the general time reversible nucleotide substitution model with gamma distribution (GTRG+G) and 1000 bootstrap replicates and nodes with bootstrap support values ≥70 are shown by an asterisk. Proposed species yet to be recognized are designated with a ^ symbol. Strains are colored by host and trees with full taxa names are provided as Supplementary Materials (Figures S1 and S2).
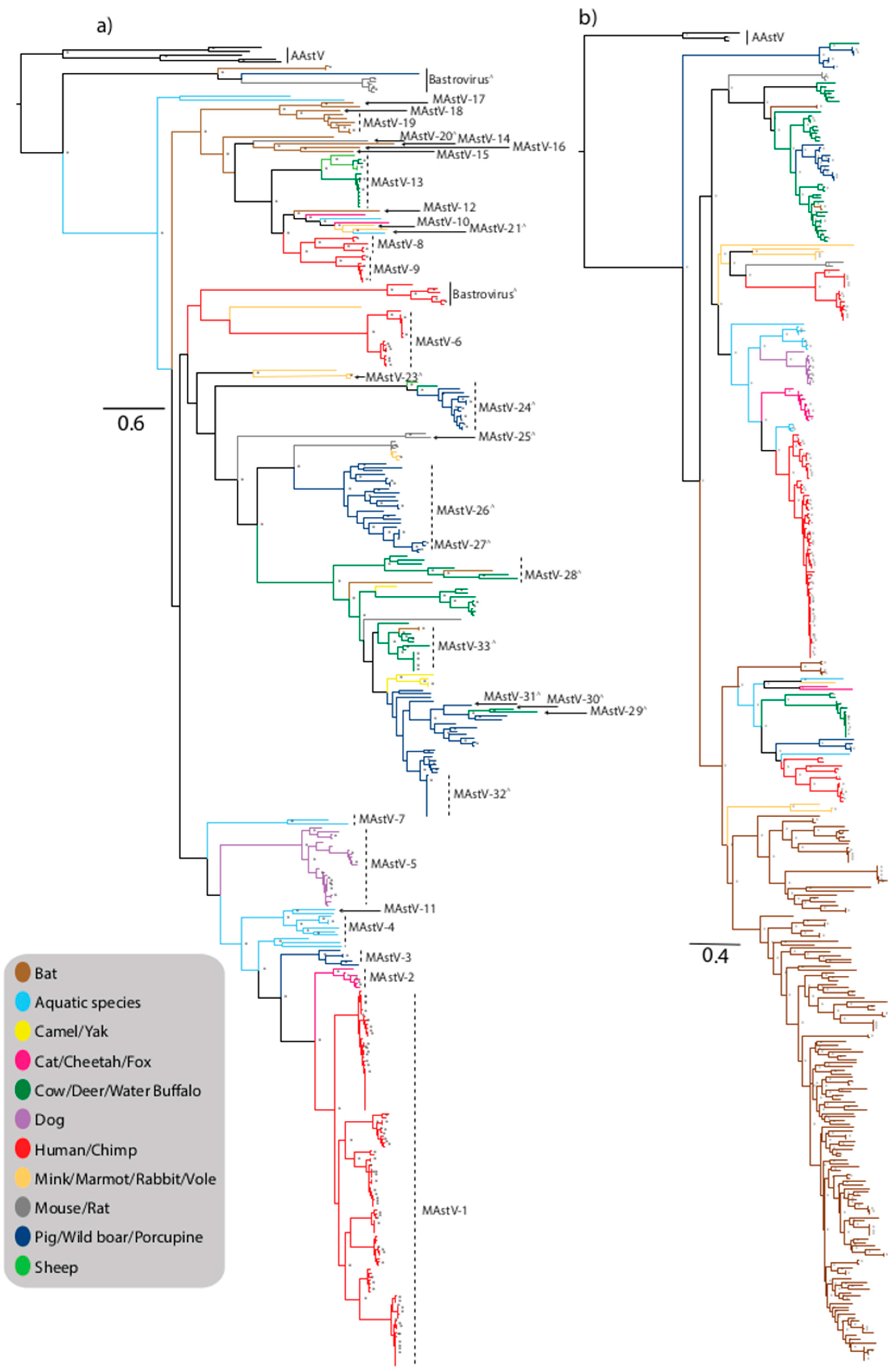
Figure 2.
Maximum-likelihood phylogenetic tree of Avastrovirus (AAstV) (a) capsid and (b) RdRp genes. Trees were generated from nucleotide sequences using the maximum-likelihood method with the GRT+G nucleotide substitution model and 1000 bootstrap replicates and nodes with bootstrap support values ≥70 are shown by an asterisk. Proposed species yet to be recognized are designated with a ^ symbol. Strains are colored by host and trees with full taxa names are provided as Supplementary Materials (Figures S3 and S4).
Figure 2.
Maximum-likelihood phylogenetic tree of Avastrovirus (AAstV) (a) capsid and (b) RdRp genes. Trees were generated from nucleotide sequences using the maximum-likelihood method with the GRT+G nucleotide substitution model and 1000 bootstrap replicates and nodes with bootstrap support values ≥70 are shown by an asterisk. Proposed species yet to be recognized are designated with a ^ symbol. Strains are colored by host and trees with full taxa names are provided as Supplementary Materials (Figures S3 and S4).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. https://doi.org/10.3390/v9050102
AMA Style
Donato C, Vijaykrishna D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses. 2017; 9(5):102. https://doi.org/10.3390/v9050102
Chicago/Turabian StyleDonato, Celeste, and Dhanasekaran Vijaykrishna. 2017. "The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses" Viruses 9, no. 5: 102. https://doi.org/10.3390/v9050102
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.