
Plant Protease Inhibitors as Emerging Antimicrobial Peptide Agents: A Comprehensive Review
 , , ,
, , ,  and
and
Abstract
:1. Introduction
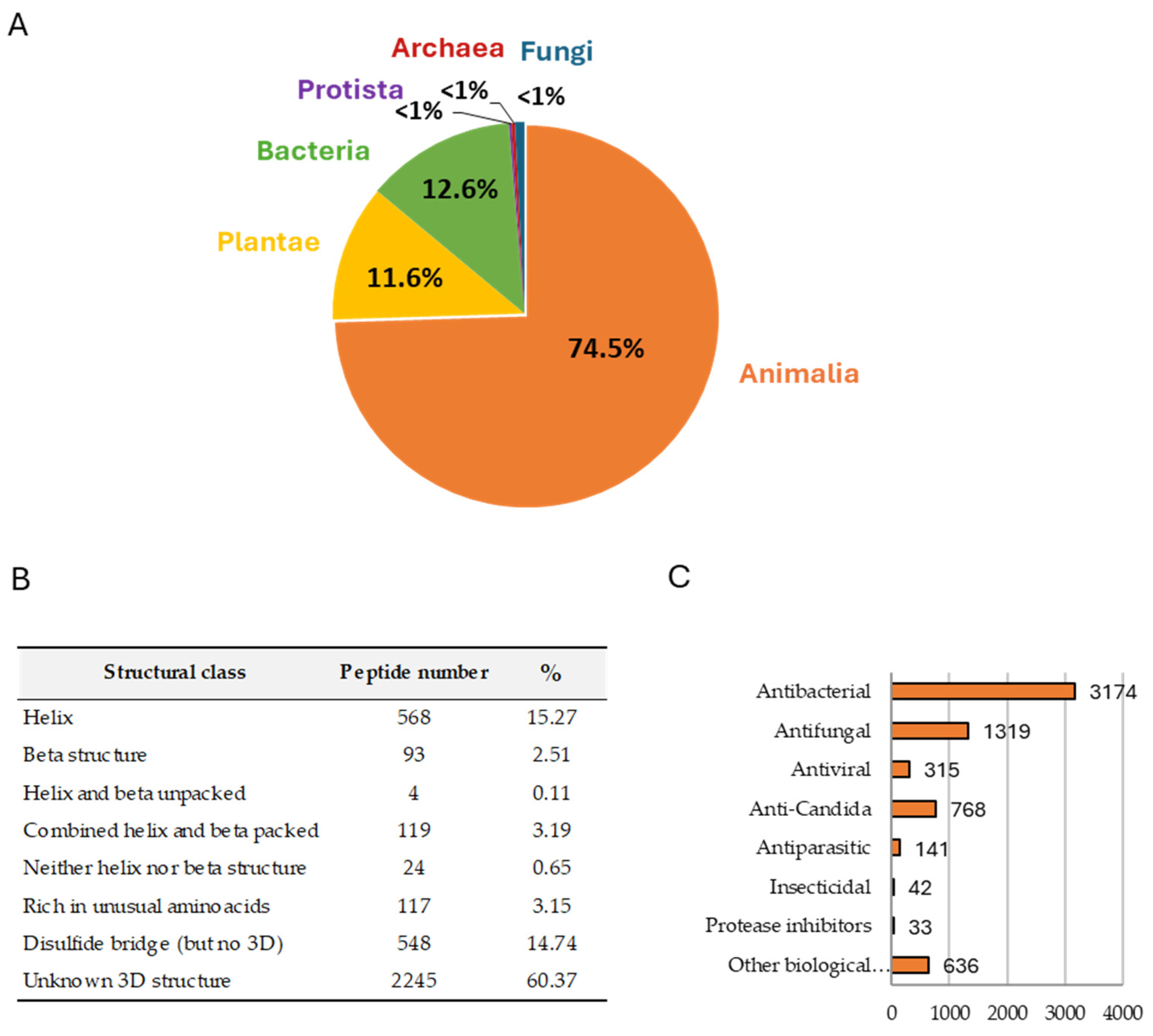
2. Antimicrobial Peptides (AMPs)
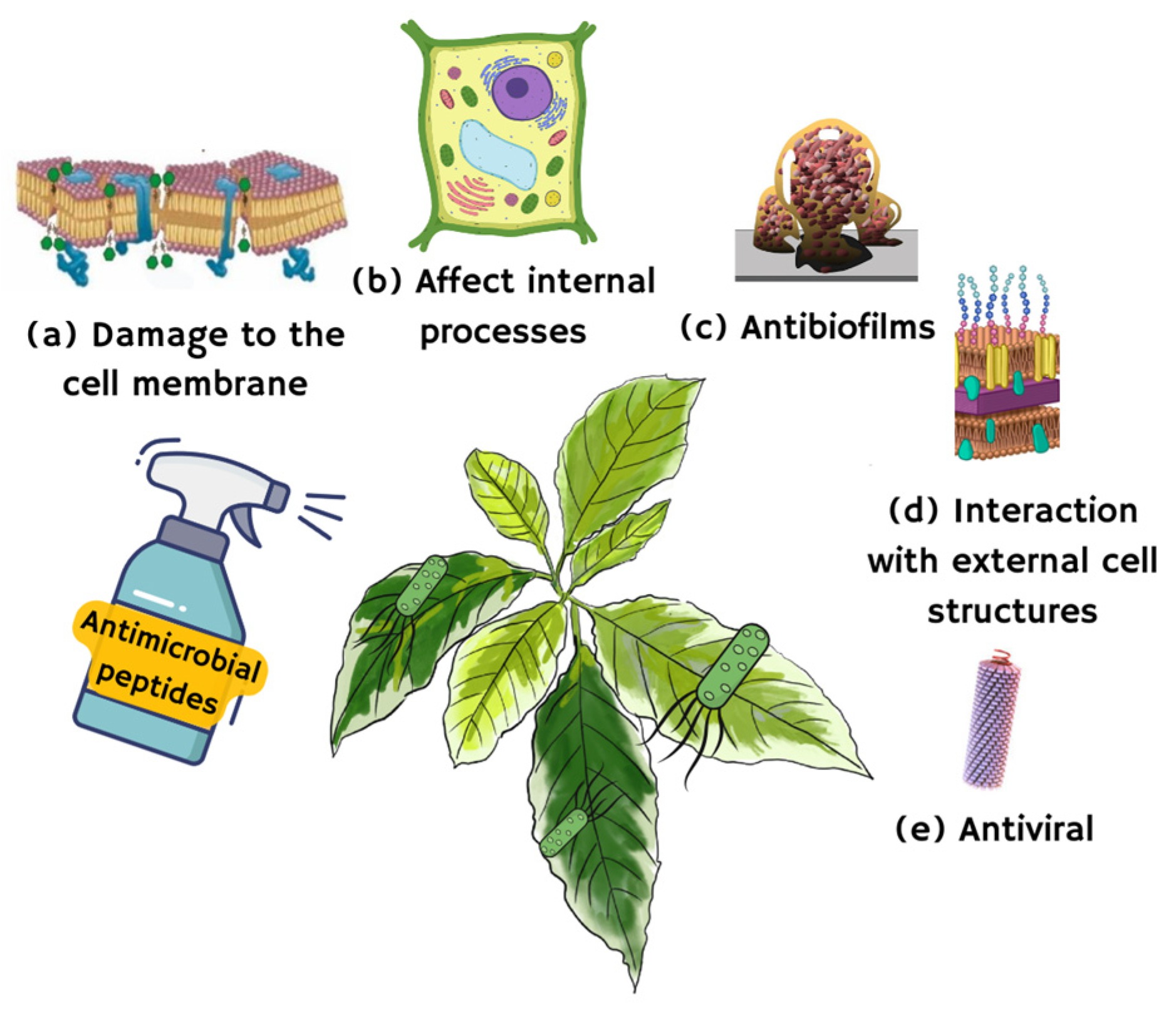
3. Mechanism of Antimicrobial Action of Plant AMPs
4. Peptide Protease Inhibitors (PPIs) Derived from Plants
5. Plant-Derived Peptide Protease Inhibitors with Antimicrobial Properties
5.1. Plant Protease Inhibitors with Antibacterial Activity
5.2. Plant Protease Inhibitors with Antifungal Activity
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lorsbach, B.A.; Sparks, T.C.; Cicchillo, R.M.; Garizi, N.V.; Hahn, D.R.; Meyer, K.G. Natural products: A strategic lead generation approach in crop protection discovery. Pest Manag. Sci. 2019, 75, 2301–2309. [Google Scholar] [CrossRef]
- Bergonzi, M.C.; Heard, C.M.; Garcia-Pardo, J. Bioactive Molecules from Plants: Discovery and Pharmaceutical Applications. Pharmaceutics 2022, 14, 2116. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Haider, R. harmaceutical and Biopharmaceuticals Industries: Revolutionizing Healthcare. Asian J. Nat. Sci. 2023, 2, 11. [Google Scholar]
- Clemons, P.A.; Bodycombe, N.E.; Carrinski, H.A.; Wilson, J.A.; Shamji, A.F.; Wagner, B.K.; Koehler, A.N.; Schreiber, S.L. Small molecules of different origins have distinct distributions of structural complexity that correlate with protein-binding profiles. Proc. Natl. Acad. Sci. USA 2010, 107, 18787–18792. [Google Scholar] [CrossRef] [PubMed]
- Despotovic, M.; de Nies, L.; Busi, S.B.; Wilmes, P. Reservoirs of antimicrobial resistance in the context of One Health. Curr. Opin. Microbiol. 2023, 73, 102291. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.W.K.; Millar, B.C.; Moore, J.E. Antimicrobial Resistance (AMR). Br. J. Biomed. Sci. 2023, 80, 11387. [Google Scholar] [CrossRef] [PubMed]
- Mudenda, S.; Daka, V.; Matafwali, S.K. World Health Organization AWaRe framework for antibiotic stewardship: Where are we now and where do we need to go? An expert viewpoint. Antimicrob. Steward. Healthc. Epidemiol. 2023, 3, e84. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.J.; McEwen, S.A. One Health-Its Importance in Helping to Better Control Antimicrobial Resistance. Trop. Med. Infect. Dis. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Hughes, J.M. Critical Importance of a One Health Approach to Antimicrobial Resistance. Ecohealth 2019, 16, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Meza, M.E.; Galarde-Lopez, M.; Carrillo-Quiroz, B.; Alpuche-Aranda, C.M. Antimicrobial resistance: One Health approach. Vet. World 2022, 15, 743–749. [Google Scholar] [CrossRef] [PubMed]
- WHO. Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 2 April 2024).
- FDA. Drugs. Available online: https://www.fda.gov/drugs (accessed on 2 April 2024).
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stocklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Parisi, M.G.; Moreno, S.; Fernandez, G. Isolation and characterization of a dual function protein from Allium sativum bulbs which exhibits proteolytic and hemagglutinating activities. Plant Physiol. Biochem. 2008, 46, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.C.; Hwang, I.; Cheong, H.; Nah, J.W.; Hahm, K.S.; Park, Y. Protease inhibitors from plants with antimicrobial activity. Int. J. Mol. Sci. 2009, 10, 2860–2872. [Google Scholar] [CrossRef]
- Shamsi, T.N.; Parveen, R.; Fatima, S. Characterization, biomedical and agricultural applications of protease inhibitors: A review. Int. J. Biol. Macromol. 2016, 91, 1120–1133. [Google Scholar] [CrossRef]
- Srivastava, S.; Dashora, K.; Ameta, K.L.; Singh, N.P.; El-Enshasy, H.A.; Pagano, M.C.; Hesham, A.E.; Sharma, G.D.; Sharma, M.; Bhargava, A. Cysteine-rich antimicrobial peptides from plants: The future of antimicrobial therapy. Phytother. Res. 2021, 35, 256–277. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, S.C.; Kim, J.Y.; Lee, S.Y.; Lim, H.T.; Cheong, H.; Hahm, K.S.; Park, Y. Purification and characterization of a heat-stable serine protease inhibitor from the tubers of new potato variety “Golden Valley”. Biochem. Biophys. Res. Commun. 2006, 346, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B. Review: Lessons Learned From Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef] [PubMed]
- Mazurkiewicz-Pisarek, A.; Baran, J.; Ciach, T. Antimicrobial Peptides: Challenging Journey to the Pharmaceutical, Biomedical, and Cosmeceutical Use. Int. J. Mol. Sci. 2023, 24, 9031. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Kundu, R. Cationic Amphiphilic Peptides: Synthetic Antimicrobial Agents Inspired by Nature. ChemMedChem 2020, 15, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, B.; van Belkum, M.J.; Diep, D.B.; Chikindas, M.L.; Ermakov, A.M.; Tiwari, S.K. Halocins, natural antimicrobials of Archaea: Exotic or special or both? Biotechnol. Adv. 2021, 53, 107834. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.I.; Mergani, A.; Aklilu, E.; Kamaruzzman, N.F. Antimicrobial Peptides: Bringing Solution to the Rising Threats of Antimicrobial Resistance in Livestock. Front. Vet. Sci. 2022, 9, 851052. [Google Scholar] [CrossRef] [PubMed]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A review on antimicrobial peptides databases and the computational tools. Database 2022, 2022, baac011. [Google Scholar] [CrossRef] [PubMed]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos-Silva, C.A.; Zupin, L.; Oliveira-Lima, M.; Vilela, L.M.B.; Bezerra-Neto, J.P.; Ferreira-Neto, J.R.; Ferreira, J.D.C.; de Oliveira-Silva, R.L.; Pires, C.J.; Aburjaile, F.F.; et al. Plant Antimicrobial Peptides: State of the Art, In Silico Prediction and Perspectives in the Omics Era. Bioinform. Biol. Insights 2020, 14, 1177932220952739. [Google Scholar] [CrossRef] [PubMed]
- Taveira, G.B.; Carvalho, A.O.; Rodrigues, R.; Trindade, F.G.; Da Cunha, M.; Gomes, V.M. Thionin-like peptide from Capsicum annuum fruits: Mechanism of action and synergism with fluconazole against Candida species. BMC Microbiol. 2016, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Taveira, G.B.; Mello, E.O.; Carvalho, A.O.; Regente, M.; Pinedo, M.; de La Canal, L.; Rodrigues, R.; Gomes, V.M. Antimicrobial activity and mechanism of action of a thionin-like peptide from Capsicum annuum fruits and combinatorial treatment with fluconazole against Fusarium solani. Biopolymers 2017, 108, e23008. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.; Austerlitz, T.; Bohlmann, T.; Bohlmann, H. The thionin family of antimicrobial peptides. PLoS ONE 2021, 16, e0254549. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zong, X.; Jin, M.; Min, J.; Wang, F.; Wang, Y. Mechanisms and regulation of defensins in host defense. Signal. Transduct. Target. Ther. 2023, 8, 300. [Google Scholar] [CrossRef]
- Tiwari, P.; Srivastava, Y.; Sharma, A.; Vinayagam, R. Antimicrobial Peptides: The Production of Novel Peptide-Based Therapeutics in Plant Systems. Life 2023, 13, 1875. [Google Scholar] [CrossRef]
- Odintsova, T.; Shcherbakova, L.; Slezina, M.; Pasechnik, T.; Kartabaeva, B.; Istomina, E.; Dzhavakhiya, V. Hevein-Like Antimicrobial Peptides Wamps: Structure-Function Relationship in Antifungal Activity and Sensitization of Plant Pathogenic Fungi to Tebuconazole by WAMP-2-Derived Peptides. Int. J. Mol. Sci. 2020, 21, 7912. [Google Scholar] [CrossRef] [PubMed]
- Molesini, B.; Treggiari, D.; Dalbeni, A.; Minuz, P.; Pandolfini, T. Plant cystine-knot peptides: Pharmacological perspectives. Br. J. Clin. Pharmacol. 2017, 83, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Postic, G.; Gracy, J.; Perin, C.; Chiche, L.; Gelly, J.C. KNOTTIN: The database of inhibitor cystine knot scaffold after 10 years, toward a systematic structure modeling. Nucleic Acids Res. 2018, 46, D454–D458. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Rogozhin, E.A. Defense Peptides From the alpha-Hairpinin Family Are Components of Plant Innate Immunity. Front. Plant Sci. 2020, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.A.; Costa, A.S.; Campos, D.C.O.; Batista, A.H.M.; Sales, G.W.P.; Nogueira, N.A.P.; Alves, K.M.M.; Coelho-de-Souza, A.N.; Oliveira, H.D. Lipid transfer protein isolated from noni seeds displays antibacterial activity in vitro and improves survival in lethal sepsis induced by CLP in mice. Biochimie 2018, 149, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Berrocal-Lobo, M.; Segura, A.; Moreno, M.; Lopez, G.; Garcia-Olmedo, F.; Molina, A. Snakin-2, an antimicrobial peptide from potato whose gene is locally induced by wounding and responds to pathogen infection. Plant Physiol. 2002, 128, 951–961. [Google Scholar] [CrossRef]
- Gould, A.; Camarero, J.A. Cyclotides: Overview and Biotechnological Applications. Chembiochem 2017, 18, 1350–1363. [Google Scholar] [CrossRef]
- Jacob, B.; Vogelaar, A.; Cadenas, E.; Camarero, J.A. Using the Cyclotide Scaffold for Targeting Biomolecular Interactions in Drug Development. Molecules 2022, 27, 6430. [Google Scholar] [CrossRef]
- Slezina, M.P.; Odintsova, T.I. Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Curr. Issues Mol. Biol. 2023, 45, 3674–3704. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, J.; Sharma, G.; Kashyap, P. Plant derived antimicrobial peptides: Mechanism of target, isolation techniques, sources and pharmaceutical applications. J. Food Biochem. 2022, 46, e14348. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Monroc, S.; Badosa, E.; Feliu, L.; Planas, M.; Montesinos, E.; Bardaji, E. De novo designed cyclic cationic peptides as inhibitors of plant pathogenic bacteria. Peptides 2006, 27, 2567–2574. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Ghimire, J.; Wu, E.; Wimley, W.C. Mechanistic Landscape of Membrane-Permeabilizing Peptides. Chem. Rev. 2019, 119, 6040–6085. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lim, S.I.; Shin, S.H.; Lim, Y.; Koh, J.W.; Yang, S. Conjugation of Cell-Penetrating Peptides to Antimicrobial Peptides Enhances Antibacterial Activity. ACS Omega 2019, 4, 15694–15701. [Google Scholar] [CrossRef]
- Montesinos, E. Functional Peptides for Plant Disease Control. Annu. Rev. Phytopathol. 2023, 61, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Le, C.F.; Fang, C.M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Pati, D.; Mohapatra, R.; Sahu, B.B.; Singh, P. The Impact of Microbes in Plant Immunity and Priming Induced Inheritance: A Sustainable Approach for Crop protection. Plant Stress 2022, 4, 100072. [Google Scholar] [CrossRef]
- Mehmood, S.; Imran, M.; Ali, A.; Munawar, A.; Khaliq, B.; Anwar, F.; Saeed, Q.; Buck, F.; Hussain, S.; Saeed, A.; et al. Model prediction of a Kunitz-type trypsin inhibitor protein from seeds of Acacia nilotica L. with strong antimicrobial and insecticidal activity. Turk. J. Biol. 2020, 44, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Choi, B.H.; Kwak, J.S.; Kang, C.W.; Lim, H.T.; Cheong, H.S.; Hahm, K.S. Kunitz-type serine protease inhibitor from potato (Solanum tuberosum L. cv. Jopung). J. Agric. Food Chem. 2005, 53, 6491–6496. [Google Scholar] [CrossRef] [PubMed]
- Clemente, M.; Corigliano, M.G.; Pariani, S.A.; Sanchez-Lopez, E.F.; Sander, V.A.; Ramos-Duarte, V.A. Plant Serine Protease Inhibitors: Biotechnology Application in Agriculture and Molecular Farming. Int. J. Mol. Sci. 2019, 20, 1345. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Huber, R. Natural protein proteinase inhibitors and their interaction with proteinases. Eur. J. Biochem. 1992, 204, 433–451. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M., Jr.; Kato, I. Protein inhibitors of proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Cotabarren, J.; Lufrano, D.; Parisi, M.G.; Obregon, W.D. Biotechnological, biomedical, and agronomical applications of plant protease inhibitors with high stability: A systematic review. Plant Sci. 2020, 292, 110398. [Google Scholar] [CrossRef] [PubMed]
- Dzik, S. Complement and Coagulation: Cross Talk Through Time. Transfus. Med. Rev. 2019, 33, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Bonturi, C.R.; Silva Teixeira, A.B.; Rocha, V.M.; Valente, P.F.; Oliveira, J.R.; Filho, C.M.B.; Fatima Correia Batista, I.; Oliva, M.L.V. Plant Kunitz Inhibitors and Their Interaction with Proteases: Current and Potential Pharmacological Targets. Int. J. Mol. Sci. 2022, 23, 4742. [Google Scholar] [CrossRef] [PubMed]
- Varkoly, K.; Beladi, R.; Hamada, M.; McFadden, G.; Irving, J.; Lucas, A.R. Viral SERPINS-A Family of Highly Potent Immune-Modulating Therapeutic Proteins. Biomolecules 2023, 13, 1393. [Google Scholar] [CrossRef] [PubMed]
- Udvardy, A. The role of controlled proteolysis in cell-cycle regulation. Eur. J. Biochem. 1996, 240, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Heutinck, K.M.; ten Berge, I.J.; Hack, C.E.; Hamann, J.; Rowshani, A.T. Serine proteases of the human immune system in health and disease. Mol. Immunol. 2010, 47, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Pearce, G. Systemin, hydroxyproline-rich systemin and the induction of protease inhibitors. Curr. Protein Pept. Sci. 2011, 12, 399–408. [Google Scholar] [CrossRef]
- Coppola, M.; Corrado, G.; Coppola, V.; Cascone, P.; Martinelli, R.; Digilio, M.C.; Pennacchio, F.; Rao, R. Prosystemin Overexpression in Tomato Enhances Resistance to Different Biotic Stresses by Activating Genes of Multiple Signaling Pathways. Plant Mol. Biol. Rep. 2015, 33, 1270–1285. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef]
- Dib, H.X.; de Oliveira, D.G.L.; de Oliveira, C.F.R.; Taveira, G.B.; de Oliveira Mello, E.; Verbisk, N.V.; Chang, M.R.; Correa Junior, D.; Gomes, V.M.; Macedo, M.L.R. Biochemical characterization of a Kunitz inhibitor from Inga edulis seeds with antifungal activity against Candida spp. Arch. Microbiol. 2019, 201, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.L.; Valadares, N.F.; Moraes, D.I.; Rosa, J.C.; Araujo, H.S.; Beltramini, L.M. Physico-chemical and antifungal properties of protease inhibitors from Acacia plumosa. Phytochemistry 2009, 70, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Cid-Gallegos, M.S.; Corzo-Rios, L.J.; Jimenez-Martinez, C.; Sanchez-Chino, X.M. Protease Inhibitors from Plants as Therapeutic Agents—A Review. Plant Foods Hum. Nutr. 2022, 77, 20–29. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, M.A.G.; Amaral, N.O.; Alvares, A.; de Oliveira, S.A.; Mehdad, A.; Honda, D.E.; Bessa, A.S.M.; Ramada, M.H.S.; Naves, L.M.; Pontes, C.N.R.; et al. Blood pressure-lowering effects of a Bowman-Birk inhibitor and its derived peptides in normotensive and hypertensive rats. Sci. Rep. 2020, 10, 11680. [Google Scholar] [CrossRef] [PubMed]
- Gitlin-Domagalska, A.; Maciejewska, A.; Debowski, D. Bowman-Birk Inhibitors: Insights into Family of Multifunctional Proteins and Peptides with Potential Therapeutical Applications. Pharmaceuticals 2020, 13, 421. [Google Scholar] [CrossRef] [PubMed]
- Bronsoms, S.; Villanueva, J.; Canals, F.; Querol, E.; Aviles, F.X. Analysis of the effect of potato carboxypeptidase inhibitor pro-sequence on the folding of the mature protein. Eur. J. Biochem. 2003, 270, 3641–3650. [Google Scholar] [CrossRef] [PubMed]
- Arolas, J.L.; Ventura, S. Protease inhibitors as models for the study of oxidative folding. Antioxid. Redox Signal. 2011, 14, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, L.S.; Murugan, K. Antimicrobial activity of protease inhibitor from leaves of Coccinia grandis (L.) Voigt. Indian J. Exp. Biol. 2011, 49, 366–374. [Google Scholar] [PubMed]
- Macedo, M.L.; Ribeiro, S.F.; Taveira, G.B.; Gomes, V.M.; de Barros, K.M.; Maria-Neto, S. Antimicrobial Activity of ILTI, a Kunitz-Type Trypsin Inhibitor from Inga laurina (SW.) Willd. Curr. Microbiol. 2016, 72, 538–544. [Google Scholar] [CrossRef] [PubMed]
- de Souza Nascimento, A.M.; de Oliveira Segundo, V.H.; Felipe Camelo Aguiar, A.J.; Piuvezam, G.; Souza Passos, T.; Florentino da Silva Chaves Damasceno, K.; de Araujo Morais, A.H. Antibacterial action mechanisms and mode of trypsin inhibitors: A systematic review. J. Enzyme. Inhib. Med. Chem. 2022, 37, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Lelio, I.D.; Romanelli, A.; Gualtieri, L.; Molisso, D.; Ruocco, M.; Avitabile, C.; Natale, R.; Cascone, P.; Guerrieri, E.; et al. Tomato Plants Treated with Systemin Peptide Show Enhanced Levels of Direct and Indirect Defense Associated with Increased Expression of Defense-Related Genes. Plants 2019, 8, 395. [Google Scholar] [CrossRef] [PubMed]
- Dabhade, A.R.; Mokashe, N.U.; Patil, U.K. Purification, characterization, and antimicrobial activity of nontoxic trypsin inhibitor from Albizia amara Boiv. Process Biochem. 2016, 51, 659–674. [Google Scholar] [CrossRef]
- Martins, T.F.; Vasconcelos, I.M.; Silva, R.G.G.; Silva, F.D.A.; Souza, P.F.N.; Varela, A.L.N.; Albuquerque, L.M.; Oliveira, J.T.A. A Bowman-Birk Inhibitor from the Seeds of Luetzelburgia auriculata Inhibits Staphylococcus aureus Growth by Promoting Severe Cell Membrane Damage. J. Nat. Prod. 2018, 81, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Arulpandi, I.; Sangeetha, R. Antibacterial Activity of Fistulin: A Protease Inhibitor Purified from the Leaves of Cassia fistula. ISRN Pharm. 2012, 2012, 584073. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.H.O.; Oliveira, C.F.R.; Rodrigues, M.S.; Neto, S.M.; Boleti, A.P.A.; Taveira, G.B.; Mello, E.O.; Gomes, V.M.; Santos, E.L.D.; Crusca, E., Jr.; et al. Adepamycin: Design, synthesis and biological properties of a new peptide with antimicrobial properties. Arch. Biochem. Biophys. 2020, 691, 108487. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.S.; de Oliveira, C.F.R.; Almeida, L.H.O.; Neto, S.M.; Boleti, A.P.A.; Dos Santos, E.L.; Cardoso, M.H.; Ribeiro, S.M.; Franco, O.L.; Rodrigues, F.S.; et al. Adevonin, a novel synthetic antimicrobial peptide designed from the Adenanthera pavonina trypsin inhibitor (ApTI) sequence. Pathog. Glob. Health 2018, 112, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Gopal, R.; Kim, S.Y.; Seo, C.H.; Lee, H.B.; Cheong, H.; Park, Y. PG-2, a Potent AMP against Pathogenic Microbial Strains, from Potato (Solanum tuberosum L. cv. Gogu Valley) Tubers Not Cytotoxic against Human Cells. Int. J. Mol. Sci. 2013, 14, 4349–4360. [Google Scholar] [CrossRef] [PubMed]
- Cotabarren, J.; Ozon, B.; Claver, S.; Geier, F.; Rossotti, M.; Garcia-Pardo, J.; Obregon, W.D. A Multifunctional Trypsin Protease Inhibitor from Yellow Bell Pepper Seeds: Uncovering Its Dual Antifungal and Hypoglycemic Properties. Pharmaceutics 2023, 15, 781. [Google Scholar] [CrossRef] [PubMed]
- Bacha, A.B.; Jemel, I.; Moubayed, N.M.S.; Abdelmalek, I.B. Purification and characterization of a newly serine protease inhibitor from Rhamnus frangula with potential for use as therapeutic drug. 3 Biotech 2017, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Manojlovic, N.T.; Solujic, S.; Sukdolak, S.; Milosev, M. Antifungal activity of Rubia tinctorum, Rhamnus frangula and Caloplaca cerina. Fitoterapia 2005, 76, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Costa, H.P.; Oliveira, J.T.; Sousa, D.O.; Morais, J.K.; Moreno, F.B.; Monteiro-Moreira, A.C.; Viegas, R.A.; Vasconcelos, I.M. JcTI-I: A novel trypsin inhibitor from Jatropha curcas seed cake with potential for bacterial infection treatment. Front. Microbiol. 2014, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shao, C.; Fang, Y.; Wang, J.; Dong, N.; Shan, A. Binding loop of sunflower trypsin inhibitor 1 serves as a design motif for proteolysis-resistant antimicrobial peptides. Acta Biomater. 2021, 124, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Cotabarren, J.; Claver, S.; Payrol, J.A.; Garcia-Pardo, J.; Obregon, W.D. Purification and Characterization of a Novel Thermostable Papain Inhibitor from Moringa oleifera with Antimicrobial and Anticoagulant Properties. Pharmaceutics 2021, 13, 512. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.A.; Lima, A.M.; Dede Oliveira BezerraSousa, D.; Nogueira, F.C.; do Sacramento Neto, J.C.; Dias, L.P.; Araujo, N.M.S.; Nagano, C.S.; Junior, H.V.N.; da Silva, C.R.; et al. Chia (Salvia hispanica L.) Seeds Contain a Highly Stable Trypsin Inhibitor with Potential for Bacterial Management Alone or in Drug Combination Therapy with Oxacillin. Probiotics. Antimicrob. Proteins 2023, 15, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Bartova, V.; Barta, J.; Jarosova, M. Antifungal and antimicrobial proteins and peptides of potato (Solanum tuberosum L.) tubers and their applications. Appl. Microbiol. Biotechnol. 2019, 103, 5533–5547. [Google Scholar] [CrossRef] [PubMed]
- Jashni, M.K.; Mehrabi, R.; Collemare, J.; Mesarich, C.H.; de Wit, P.J. The battle in the apoplast: Further insights into the roles of proteases and their inhibitors in plant-pathogen interactions. Front. Plant Sci. 2015, 6, 584. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Meulenhoff, J.S.; Sela-Buurlage, M.; van den Elzen, P.J.; Cornelissen, B.J. Pathogen-induced proteins with inhibitory activity toward Phytophthora infestans. Plant Cell 1991, 3, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Vernekar, J.V.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Novel bifunctional alkaline protease inhibitor: Protease inhibitory activity as the biochemical basis of antifungal activity. Biochem. Biophys. Res. Commun. 2001, 285, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Gutierrez, D.; Frohlich, K.U.; Kroemer, G.; Madeo, F. Metacaspases are caspases. Doubt no more. Cell Death Differ. 2010, 17, 377–378. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic Resistance: One Health One World Outlook. Front. Cell Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef] [PubMed]
- Woods, M.; McAlister, J.A.; Geddes-McAlister, J. A One Health approach to overcoming fungal disease and antifungal resistance. WIREs Mech. Dis. 2023, 15, e1610. [Google Scholar] [CrossRef] [PubMed]
- FAO. Antimicrobial Resistance. Available online: https://www.fao.org/antimicrobial-resistance/key-sectors/animal-production/en/ (accessed on 2 April 2024).
- Barcenas, O.; Pintado-Grima, C.; Sidorczuk, K.; Teufel, F.; Nielsen, H.; Ventura, S.; Burdukiewicz, M. The dynamic landscape of peptide activity prediction. Comput. Struct. Biotechnol. J. 2022, 20, 6526–6533. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Song, L.; Zhu, S.; Fu, X.; Li, X.; He, C.; Li, J. Machine learning assisted rational design of antimicrobial peptides based on human endogenous proteins and their applications for cosmetic preservative system optimization. Sci. Rep. 2024, 14, 947. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Family | Plant Source | Name | Type of PPI | MW (kDa) | Activity | Target Microorganisms | Ref. |
|---|---|---|---|---|---|---|---|
| Fabaceae (Leguminosae) | Inga edulis | Inga Edulis Trypsin Inhibitor (IETI) | Kunitz | 19.7 | Antifungal | Candida tropicalis and Candida buinensis | [76] |
| Inga laurina | Inga Laurina Trypsin Inhibitor (ILTI) | Kunitz | 20 | Antifungal | C. tropicalis and C. buinensis | [84] | |
| Acacia plumosa | Acacia plumosa Trypsin Inhibitor (ApTI A, B, C) | Kunitz | 20 | Antifungal | Aspergillus niger, Thielaviopsis paradoxa, and Colletotrichum spp. | [77] | |
| Acacia nilotica | Acacia nilotica Trypsin Inhibitor (AnTI) | Trypsin inhibitor | 21 | Antibacterial | G+: Staphylococcus aureus and Bacillus subtilis, G−: Escherichia coli, and Pseudomonas aeruginosa | [62] | |
| Albizia amara | Proteinaceous protease inhibitor (API) | Unknown | 49 | Antibacterial | G+: B. subtilis G−: P. aeruginosa | [87] | |
| Antifungal | C. albicans | ||||||
| Luetzelburgia auriculata | Luetzelburgia auriculata Bowman-Birk protease inhibitor (LzaBBI) | Bowman-Birk | 14.3 | Antibacterial | G+: S. aureus | [88] | |
| Cassia fistula | Fistulin | Trypsin inhibitor | 4 | Antibacterial | G+: S. aureus, and B. subtilis G−: Klebsiella pneumoniae, E. coli, and P. aeruginosa | [89] | |
| Adenanthera pavonina | Adepamycin | Trypsin inhibitor | 2.4 | Antibacterial | G+: S. aureus G−: E. coli, Klebsiella oxytoca, K. pneumoniae, and P. aeruginosa | [90,91] | |
| Antifungal | C. albicans and C. tropicalis | ||||||
| Solanaceae | Solanum tuberosum L. Cv. Golden Valley | Potide-G | Kunitz | 5.57 | Antibacterial | G+: S. aureus, and Listeria monocytogenes G−: E. coli | [26] |
| Antifungal | C. albicans and Rhizoctonia solani | ||||||
| S. tuberosum L. Cv. Gogu Valley | Peptide G2 (PG-2) | Kunitz | 3.2 | Antibacterial | G+: S. aureus | [92] | |
| Antifungal | C. albicans | ||||||
| S. tuberosum L. Cv. L. Jopung | Antifungal protein J (AFP-J) | Kunitz | 13.5 | Antifungal | C. albicans, Trichosporon beigelii, and Saccharomyces cerevisiae | [63] | |
| Capsicum annuum L. | Yellow Bell Pepper Trypsin Inhibitor (YBPTI) | Trypsin inhibitor | Antifungal | C. albicans | [93] | ||
| Cucurbitaceae | Coccinia grandis (L.) Voigt. | Coccinia grandis (L.) inhibitor | Protease inhibitor | 14.3 | Antibacterial | G+: S. aureus, and B. subtilis G−: K. pneumoniae, Proteus vulgaris, and E. coli | [83] |
| Antifungal | C. albicans, Mucor indicus, Penicillium notatum, Aspergillus flavus Aspergillus oryzae, and Cryptococcus neoformans | ||||||
| Rhamnaceae | Rhamnus frangula | RflP-1 | Kunitz | 22.5 | Antibacterial | G+: Bacillus sp. and Bacillus licheniformis | [94,95] |
| Euphorbiaceae | Jatropha curcas | Jatropha curcas Trypsin inhibitor I (JcTI-I) | Trypsin inhibitor | 10.2 | Antibacterial | G+: S. aureus G−: Salmonella enterica | [96] |
| Asteraceae | Helianthus annuus | Sunflower Trypsin Inhibitor 1 (SFTI1) | Trypsin inhibitor | Antibacterial | G+: Staphylococcus epidermidis, S. aureus, and Enterococcus faecalis G−: E. coli, P. aeruginosa, and Salmonella typhimurium | [97] | |
| Moringaceae | Moringa oleifera | Moringa oleifera Protein Inhibitor (MoPI) | Phytocystatin | 19 | Antibacterial | G+: S. aureus, and E. faecalis | [98] |
| Lamiaceae | Salvia hispanica | Salvia hispanica Trypsin Inhibitor (ShTI) | Trypsin inhibitor | 11 | Antibacterial | G+: Methicillin-resistant S. aureus | [99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, M.G.; Ozón, B.; Vera González, S.M.; García-Pardo, J.; Obregón, W.D. Plant Protease Inhibitors as Emerging Antimicrobial Peptide Agents: A Comprehensive Review. Pharmaceutics 2024, 16, 582. https://doi.org/10.3390/pharmaceutics16050582
Parisi MG, Ozón B, Vera González SM, García-Pardo J, Obregón WD. Plant Protease Inhibitors as Emerging Antimicrobial Peptide Agents: A Comprehensive Review. Pharmaceutics. 2024; 16(5):582. https://doi.org/10.3390/pharmaceutics16050582
Chicago/Turabian StyleParisi, Mónica G., Brenda Ozón, Sofía M. Vera González, Javier García-Pardo, and Walter David Obregón. 2024. "Plant Protease Inhibitors as Emerging Antimicrobial Peptide Agents: A Comprehensive Review" Pharmaceutics 16, no. 5: 582. https://doi.org/10.3390/pharmaceutics16050582