
Study on the Relationship between Topological Characteristics of Vegetation Ecospatial Network and Carbon Sequestration Capacity in the Yellow River Basin, China

 , ,
, ,  ,
,
Abstract
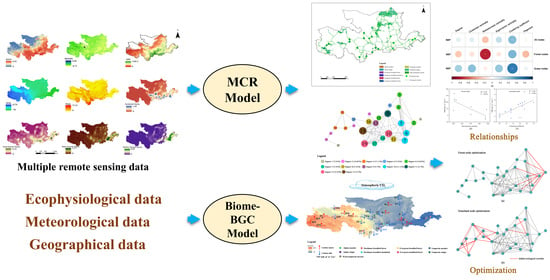
:
1. Introduction
2. Materials and Methods
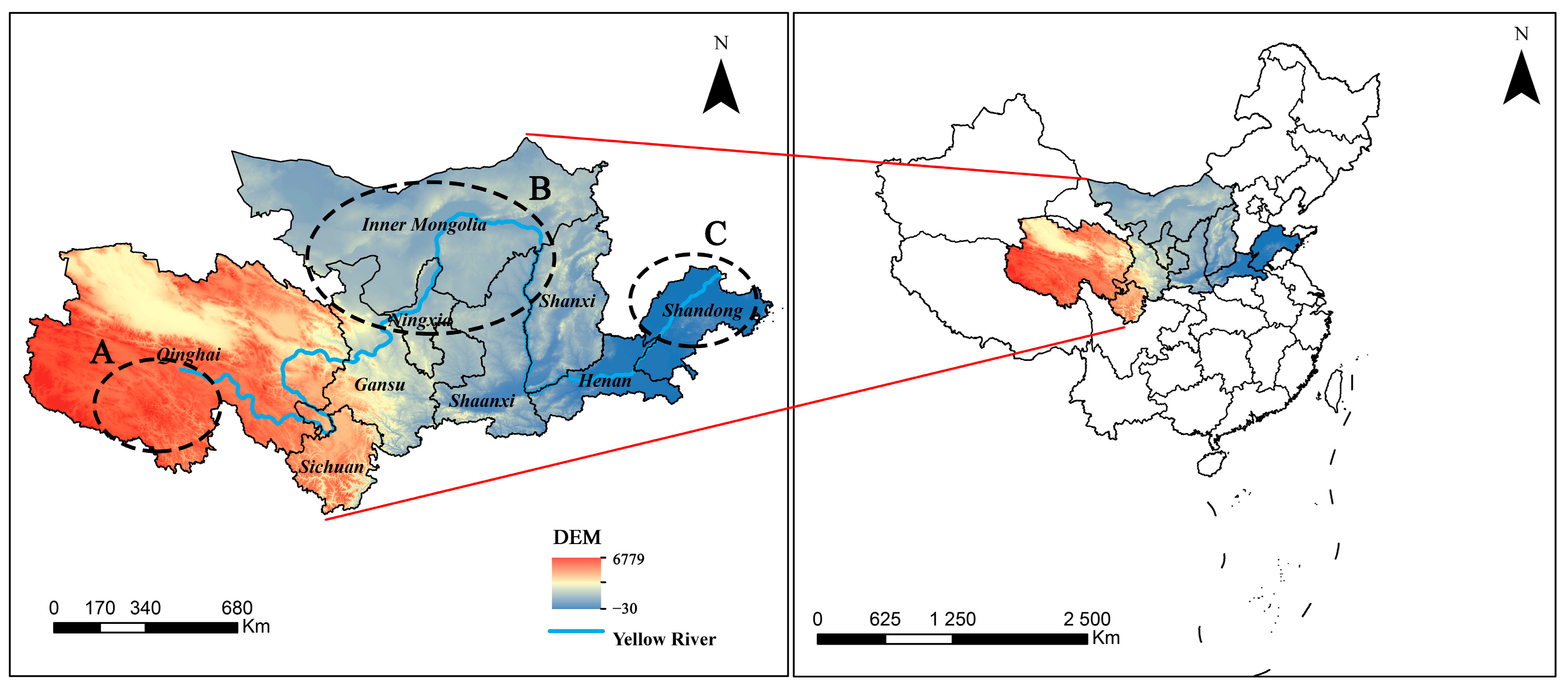
2.1. Study Area
2.1.1. Introduction to the Yellow River Basin
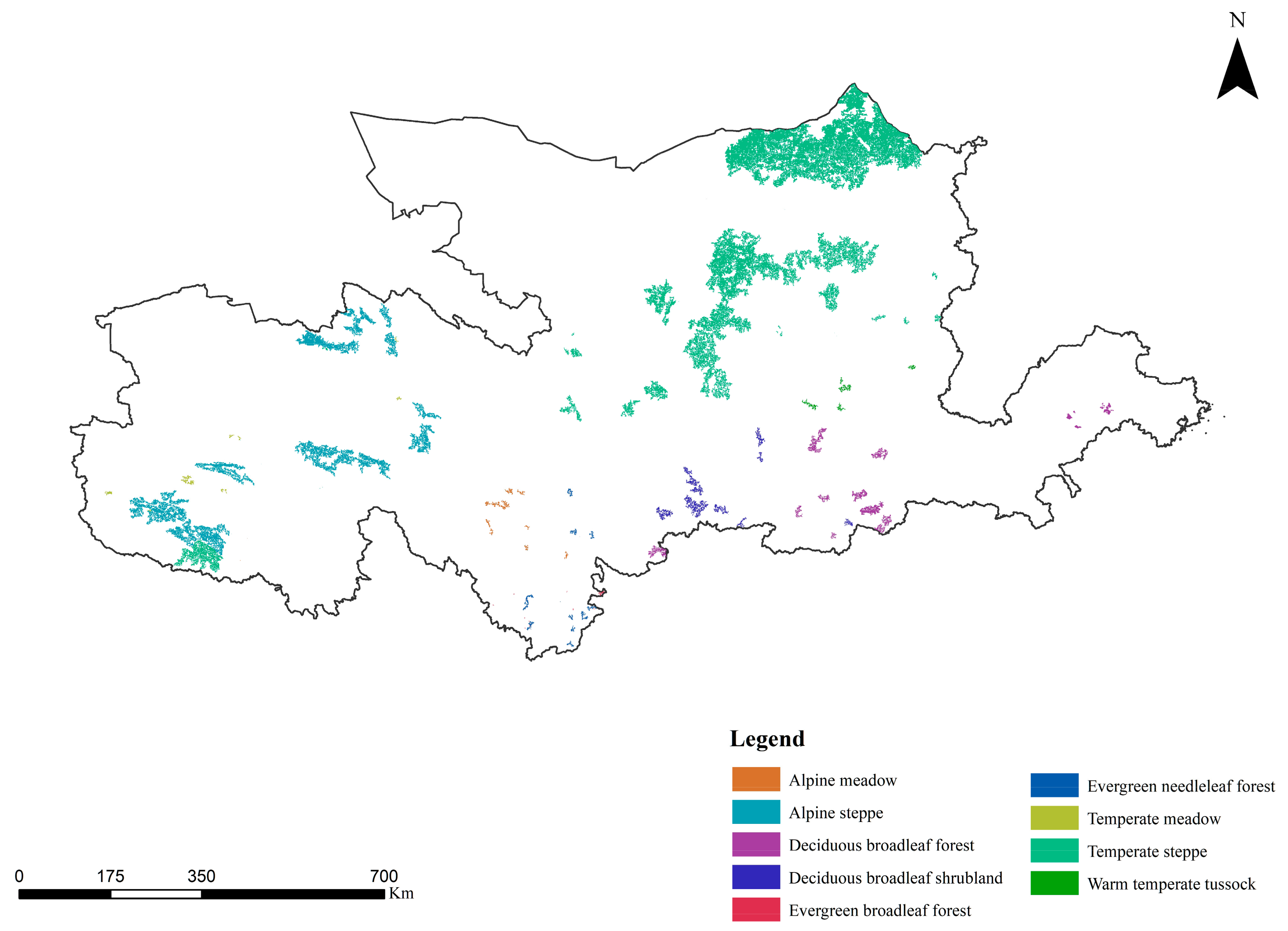
2.1.2. Vegetation Types in the Yellow River Basin
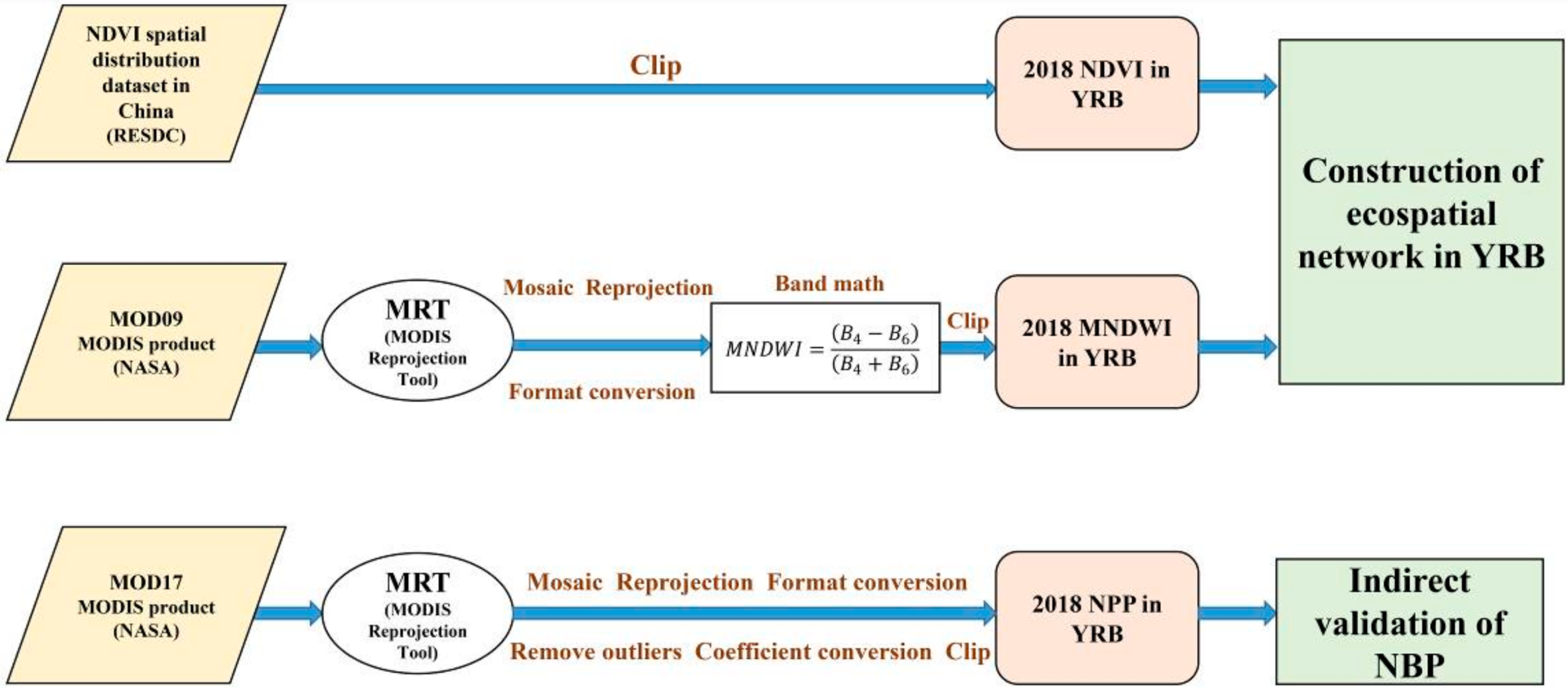
2.2. Data Sources and Processing of MODIS Products
2.2.1. Data Sources and Descriptions
2.2.2. Processing of MODIS Products
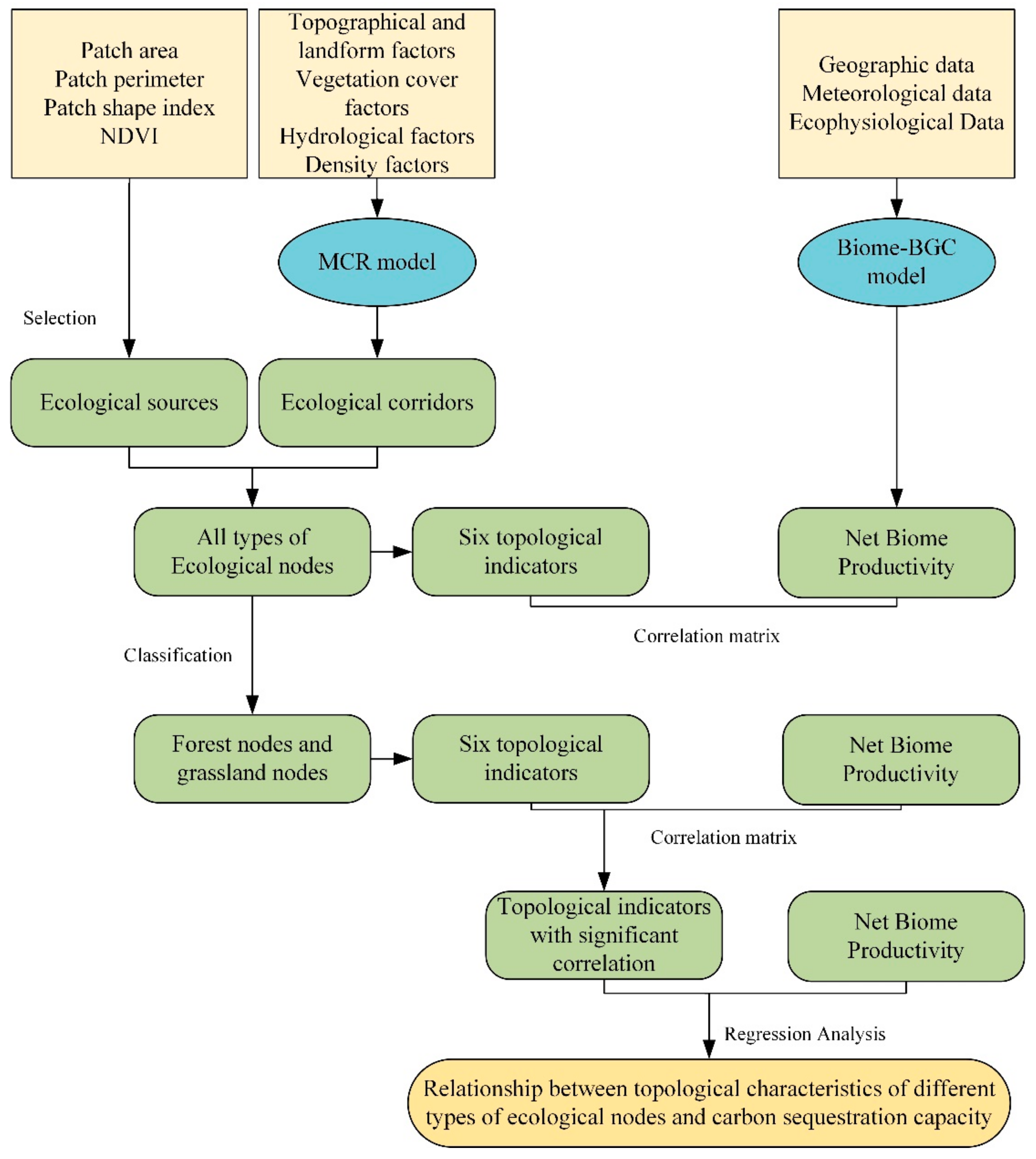
2.3. Construction of Ecospatial Network
2.3.1. Introduction to Ecospatial Network
2.3.2. Identification of Ecological Sources
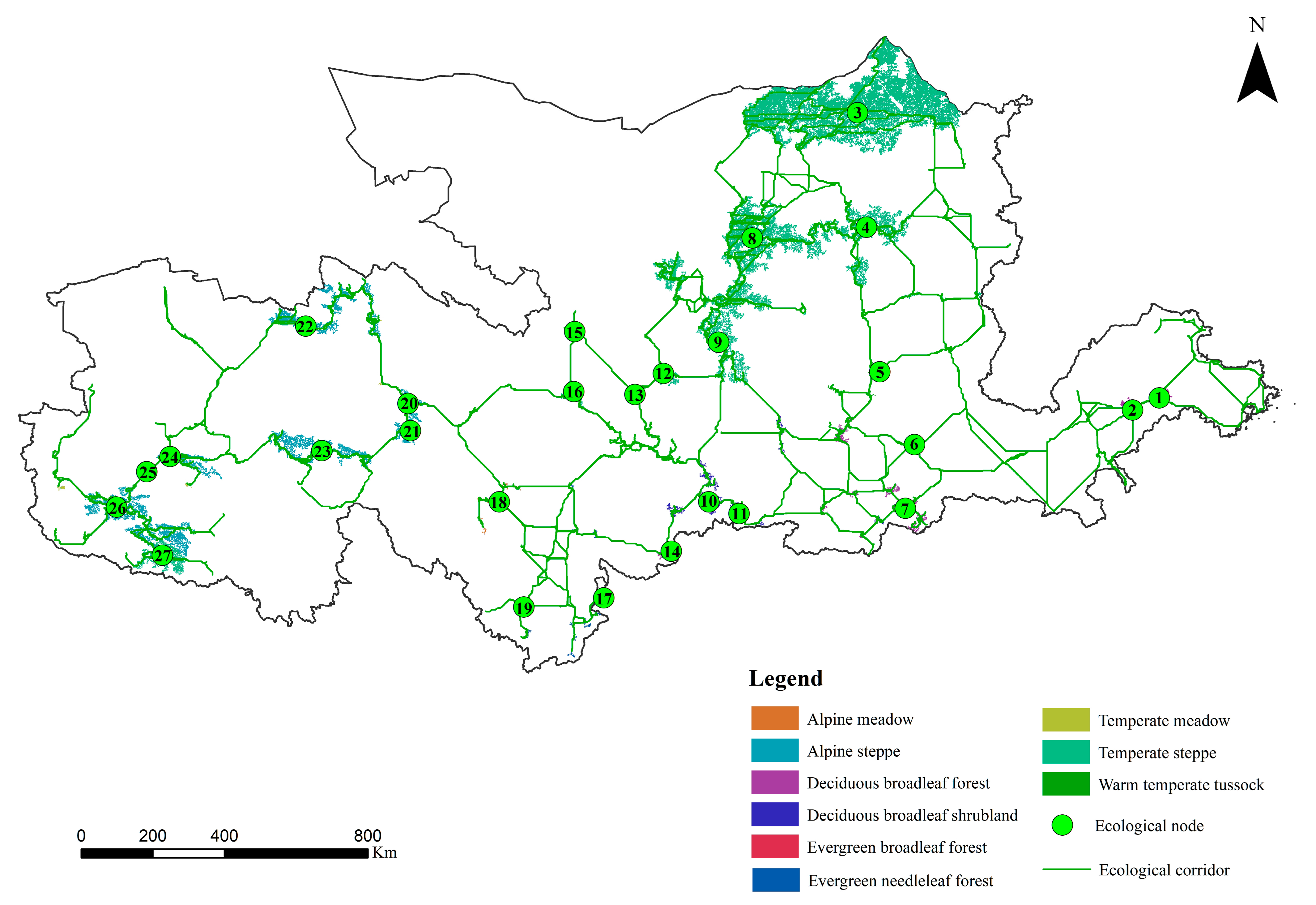
2.3.3. Extraction of Ecological Corridors and Ecological Nodes
2.4. Algorithm for Topological Indicators
2.5. Evaluation of the Indicator of Carbon Sequestration Ability
2.6. Biome-BGC Model and Its Validation
2.6.1. Description of the Biome-BGC Model
2.6.2. Indirect Validation of the Biome-BGC Model
2.7. Statistical Analysis and Technical Routes
3. Results
3.1. Results of Factors for Constructing Ecospatial Networks
3.2. The Construction Process and Results of Vegetation Ecospatial Network in the YRB
3.2.1. Extraction of Ecological Sources
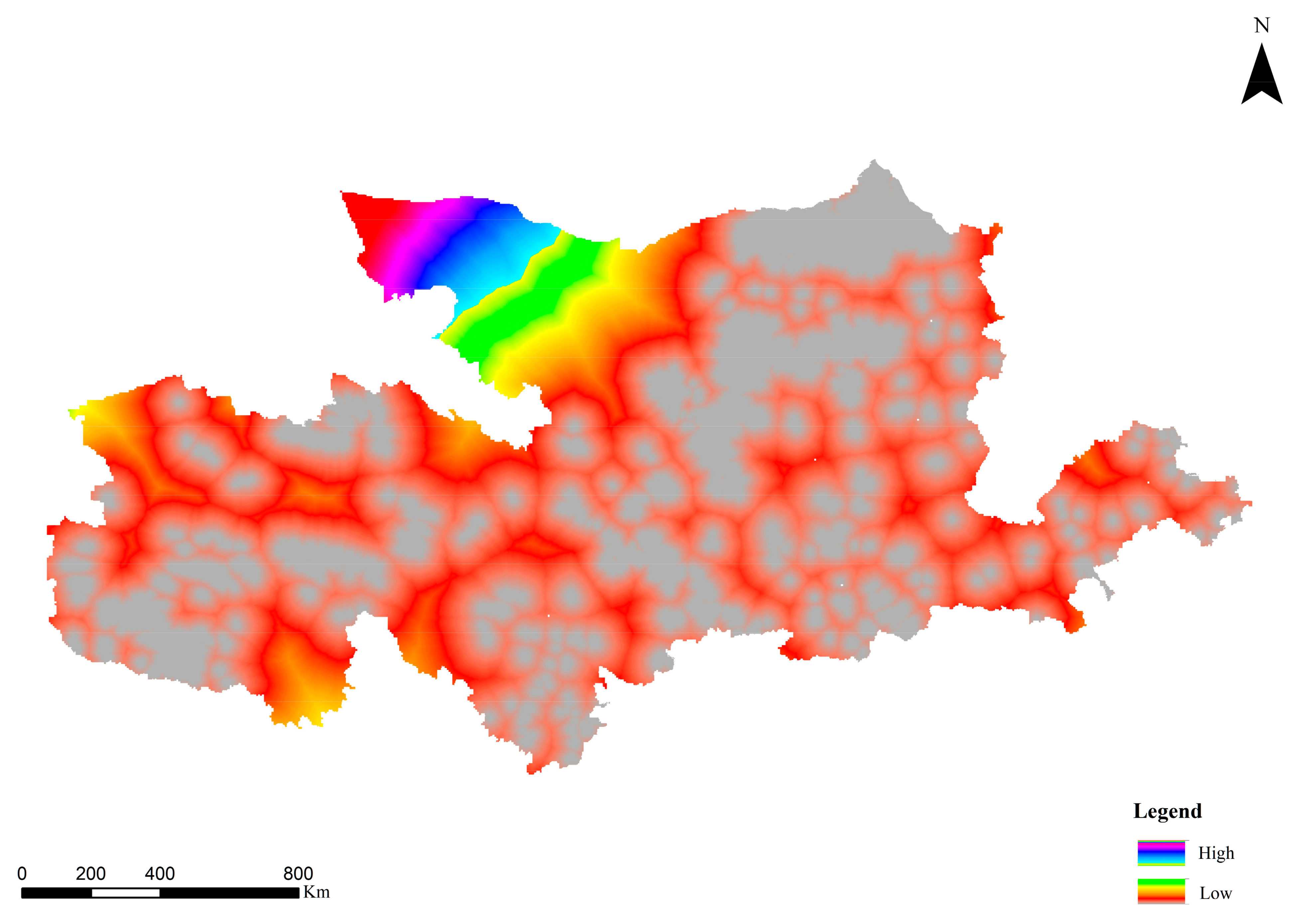
3.2.2. Results of Cumulative Resistance Surface
3.2.3. Construction of Vegetation Ecospatial Network in the YRB
3.3. Results of Topological Indicators
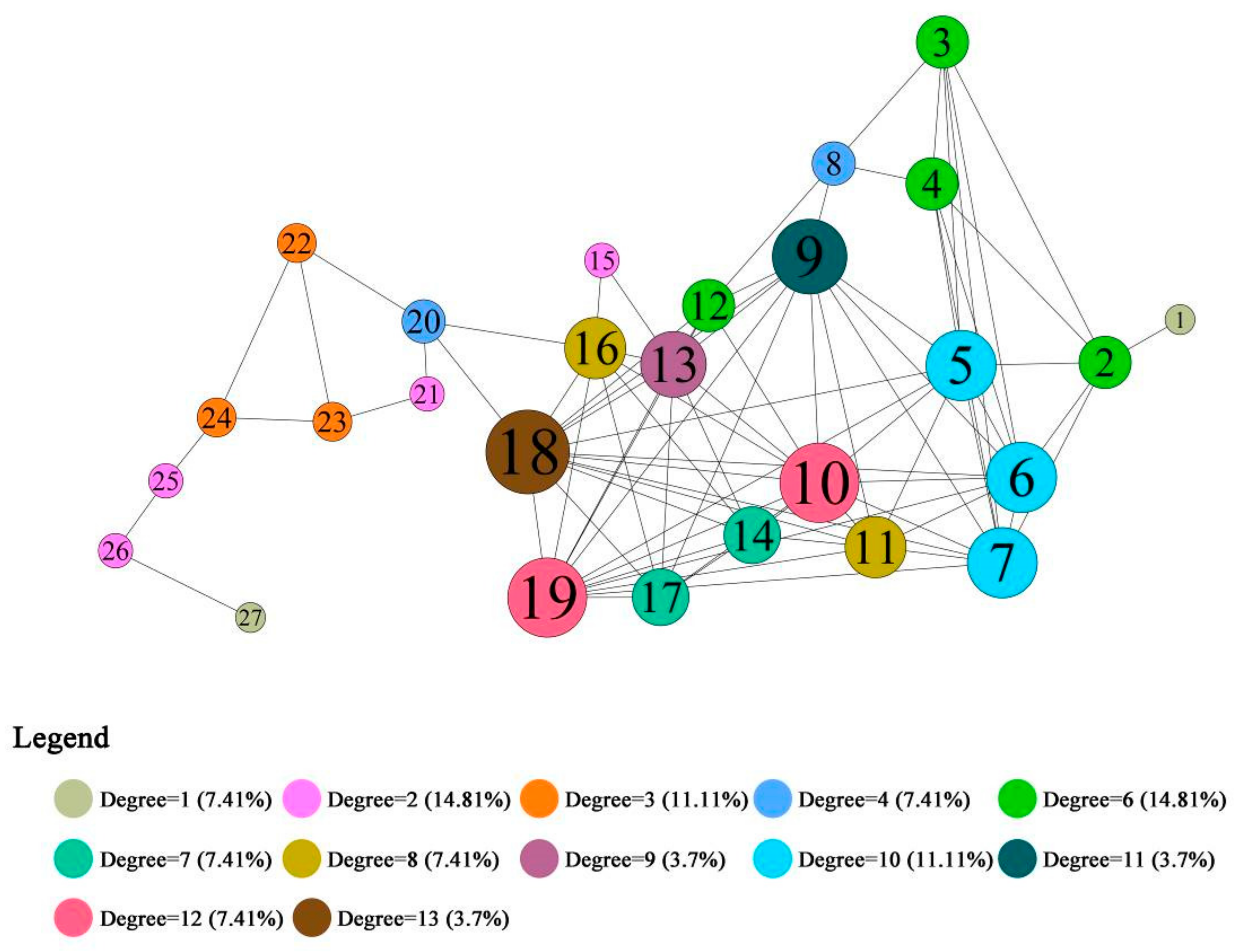
3.3.1. Results of the Topological Indicators Describing the Ecological Nodes
3.3.2. Results of the Topological Indicators Describing the Network
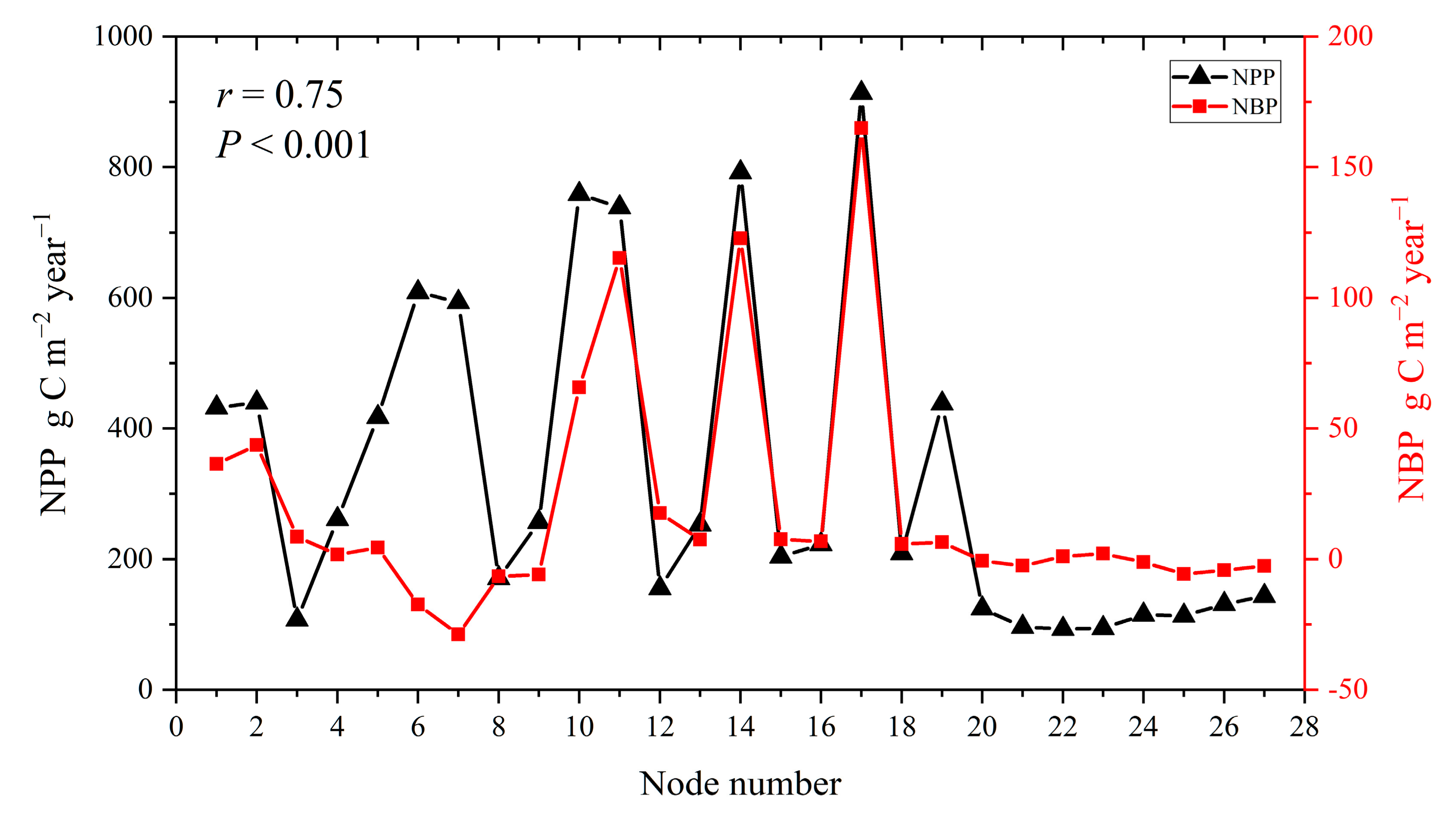
3.4. NBP Simulation Results and Validation
3.4.1. NBP Simulation Results for Ecological Nodes
3.4.2. Results of Indirect Validation of NBP
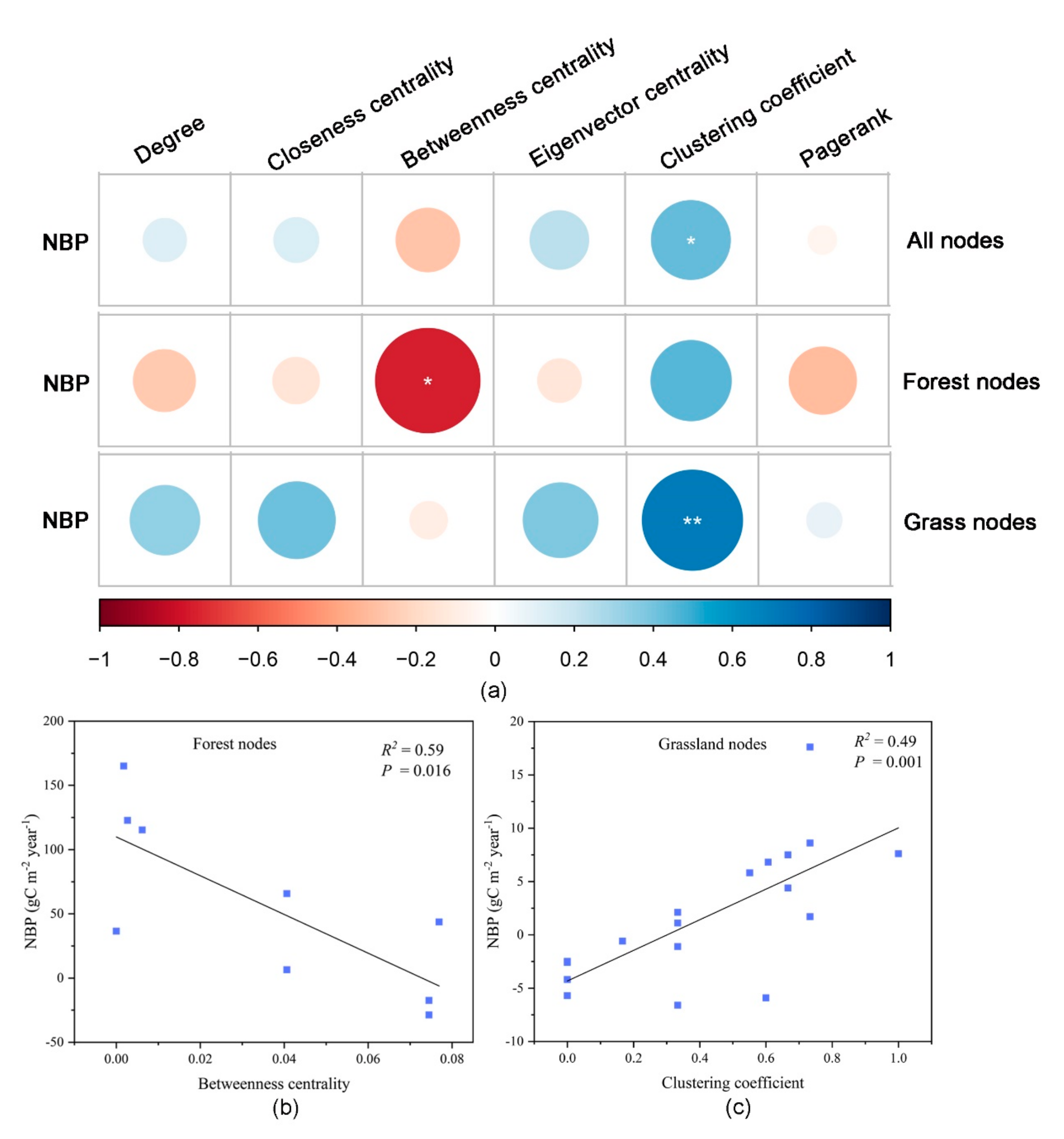
3.5. Relationship between Topological Indicators and Carbon Sequestration Capacity of Ecological Nodes
4. Discussions
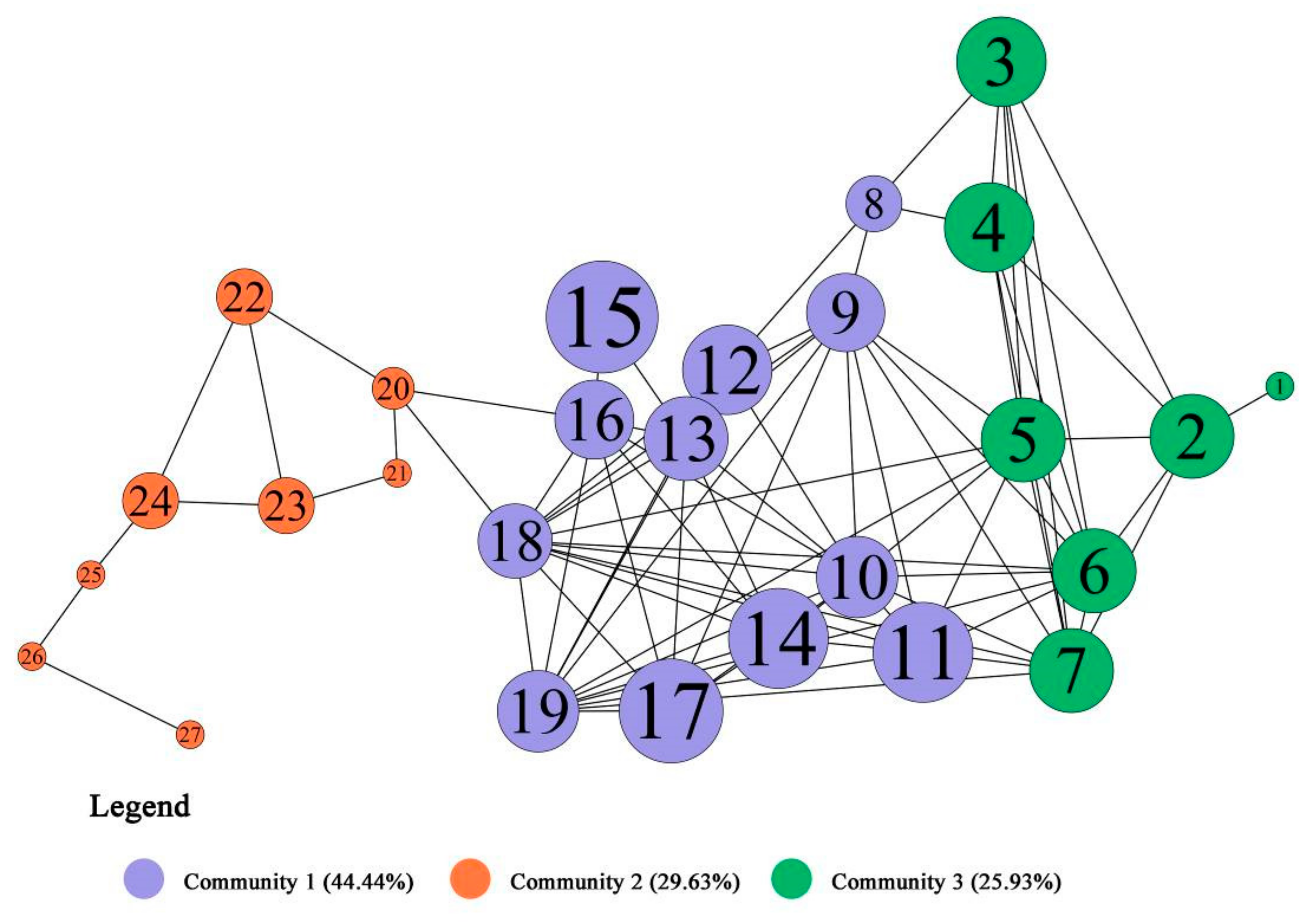
4.1. Community Division and Aggregation of Nodes
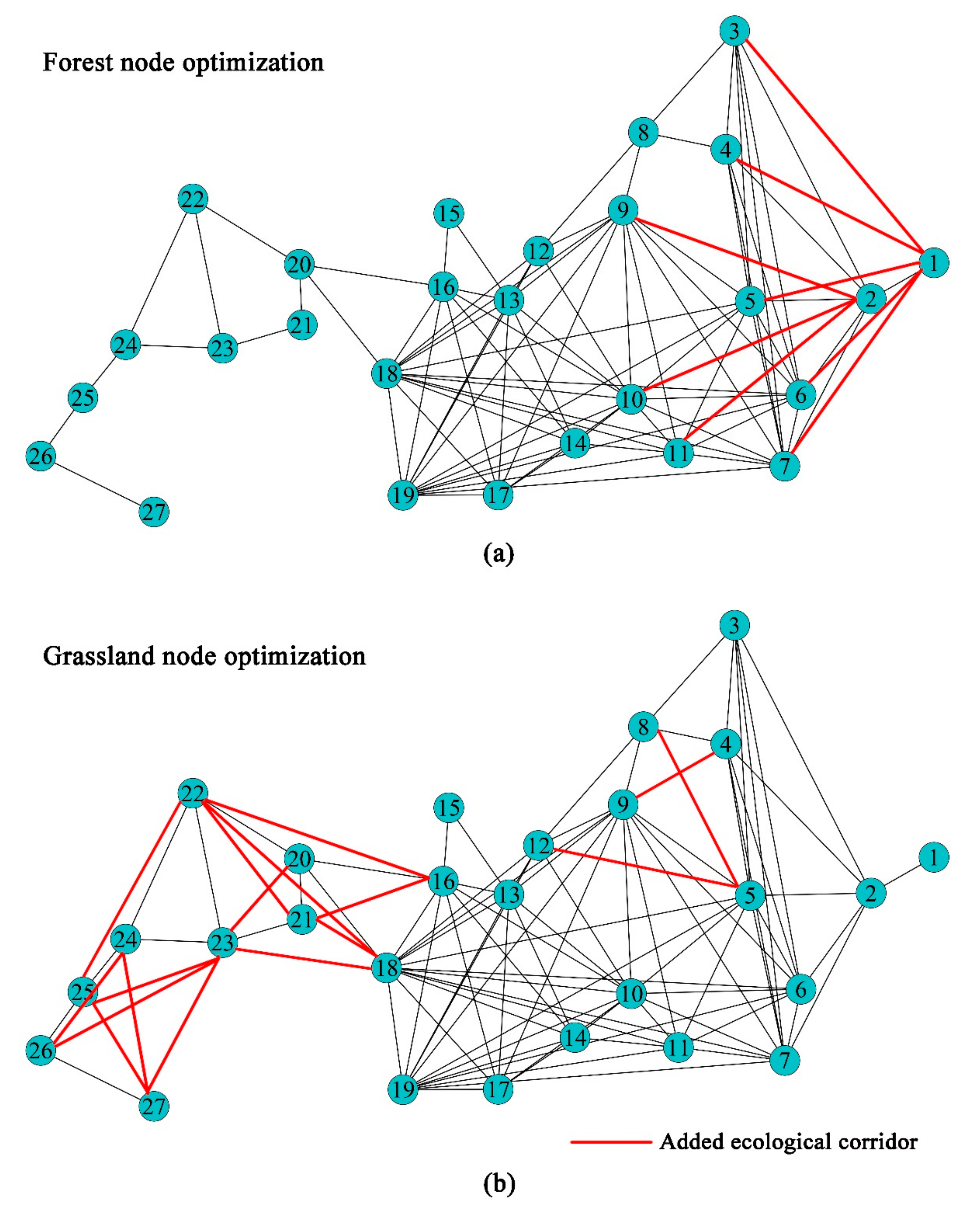
4.2. Suggestions for Optimizing the Vegetation Ecospatial Network in the YRB
4.3. Prospects and Limitations of This Study
5. Conclusions
- (1)
- The vegetation ecospatial network of the YRB in 2018 had a large clustering coefficient, a small-world characteristic, and a relatively stable overall structure. Moreover, the vegetation ecospatial network of the YRB had more carbon sink nodes than carbon source nodes in 2018.
- (2)
- The net carbon sequestration capacity of forest nodes in the YRB had a negative linear correlation with its betweenness centrality; the carbon sequestration capacity of grassland nodes had a positive linear correlation with its clustering coefficient.
- (3)
- For the purpose of increasing the carbon sequestration capacity of vegetation, we put forward optimization suggestions for the vegetation ecospatial network in the YRB. In addition, the eastern region of the YRB is a key optimization area for forest nodes, and the western region is a key optimization area for grass nodes.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_Full_Report.pdf. (accessed on 15 October 2021).
- Broadstock, D.; Ji, Q.; Managi, S.; Zhang, D. Pathways to Carbon Neutrality: Challenges and Opportunities; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of national ecological restoration projects on carbon sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Yu, G.; Liu, L.; Hu, S.; Chapin, F.S. Climate change, human impacts, and carbon sequestration in China. Proc. Natl. Acad. Sci. USA 2018, 115, 4015–4020. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ni, J.; Tenhunen, J. Application of a geographically-weighted regression analysis to estimate net primary production of Chinese forest ecosystems. Glob. Ecol. Biogeogr. 2005, 14, 379–393. [Google Scholar] [CrossRef]
- Ding, Z.L. Research on China’s Carbon Neutrality Framework Roadmap. China Ind. Inf. Technol. 2021, 8, 54–61. [Google Scholar]
- Diodato, N.; Bellocchi, G. Spatial probability modelling of forest productivity indicator in Italy. Ecol. Indic. 2019, 108, 105721. [Google Scholar] [CrossRef]
- Jiang, H.; Apps, M.J.; Zhang, Y.; Peng, C.; Woodard, P.M. Modelling the spatial pattern of net primary productivity in Chinese forests. Ecol. Model. 1999, 122, 275–288. [Google Scholar] [CrossRef]
- Chen, J.M.; Ju, W.; Cihlar, J.; Price, D.; Liu, J.; Chen, W.; Pan, J.; Black, A.; Barr, A. Spatial distribution of carbon sources and sinks in Canada’s forests. Tellus Chem. Phys. Meteorol. 2003, 55, 622–641. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.; Zhang, Y.; Huang, M.; Tao, B.; Guo, R.; Yan, C. Effects of climate warming on net primary productivity in China during 1961–2010. Ecol. Evol. 2017, 7, 6736–6746. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Thompson, J.R. Climate change imposes phenological trade-offs on forest net primary productivity. J. Geophys. Res. Biogeosci. 2017, 122, 2298–2313. [Google Scholar] [CrossRef]
- Tobler, W.R. A Computer Movie Simulating Urban Growth in the Detroit Region. Econ. Geogr. 1970, 46, 234–240. [Google Scholar] [CrossRef]
- Chang, D.H.S.; Gauch, H.G., Jr. Multivariate analysis of plant communities and environmental factors in Ngari, Tibet. Ecology 1986, 67, 1568–1575. [Google Scholar] [CrossRef]
- Wu, Z.; Lin, C.; Su, Z.; Zhou, S.; Zhou, H. Multiple landscape “source–sink” structures for the monitoring and management of non-point source organic carbon loss in a peri-urban watershed. Catena 2016, 145, 15–29. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, D.; Qiao, X.; Jiang, M.; Li, Q.; Gu, Z.; Liu, F. Forest dynamics and carbon storage under climate change in a subtropical mountainous region in central China. Ecosphere 2020, 11, e03072. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Chen, Y.; Tan, Z.; Li, S.; Ai, B. A new landscape index for quantifying urban expansion using multi-temporal remotely sensed data. Landsc. Ecol. 2010, 25, 671–682. [Google Scholar] [CrossRef]
- Costa, L.D.F.; Rodrigues, F.A.; Travieso, G.; Boas, P.R.V. Characterization of complex networks: A survey of measurements. Adv. Phys. 2007, 56, 167–242. [Google Scholar] [CrossRef] [Green Version]
- Bombrun, M.; Dash, J.P.; Pont, D.; Watt, M.S.; Pearse, G.D.; Dungey, H. Forest-Scale Phenotyping: Productivity Characterisation Through Machine Learning. Front. Plant Sci. 2020, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.; Barabási, A.-L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47–97. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Yue, D.; Wang, Y.; Kai, S.; Fang, M.; Ma, H.; Zhang, Q.; Huang, Y. Optimization of ecological node layout and stability analysis of ecological network in desert oasis: A typical case study of ecological fragile zone located at Deng Kou County (Inner Mongolia). Ecol. Indic. 2018, 84, 304–318. [Google Scholar] [CrossRef]
- Guo, H.; Yu, Q.; Pei, Y.; Wang, G.; Yue, D. Optimization of landscape spatial structure aiming at achieving carbon neutrality in desert and mining areas. J. Clean. Prod. 2021, 322, 129156. [Google Scholar] [CrossRef]
- Xu, W.; Wang, J.; Zhang, M.; Li, S. Construction of landscape ecological network based on landscape ecological risk assessment in a large-scale opencast coal mine area. J. Clean. Prod. 2021, 286, 125523. [Google Scholar] [CrossRef]
- Watt, M.S.; Palmer, D.J.; Dungey, H.; Kimberley, M.O. Predicting the spatial distribution of Cupressus lusitanica productivity in New Zealand. For. Ecol. Manag. 2009, 258, 217–223. [Google Scholar] [CrossRef]
- Su, H.Z. Analyzing and Simulating the Growth of Picea Schrenkiana Forests in Xinjiang under Global Climate Change. Ph.D Thesis, Institute of Botany, the Chinese Academy of Sciences, Beijing, China, 2005. [Google Scholar]
- Chen, Y.-P.; Fu, B.-J.; Zhao, Y.; Wang, K.-B.; Zhao, M.M.; Ma, J.-F.; Wu, J.-H.; Xu, C.; Liu, W.-G.; Wang, H. Sustainable development in the Yellow River Basin: Issues and strategies. J. Clean. Prod. 2020, 263, 121223. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Yang, H.; Zhao, Y.; Li, Z.; Wu, J. Capability of Remotely Sensed Drought Indices for Representing the Spatio–Temporal Variations of the Meteorological Droughts in the Yellow River Basin. Remote Sens. 2018, 10, 1834. [Google Scholar] [CrossRef] [Green Version]
- Wohlfart, C.; Liu, G.; Huang, C.; Kuenzer, C. A River Basin over the Course of Time: Multi-Temporal Analyses of Land Surface Dynamics in the Yellow River Basin (China) Based on Medium Resolution Remote Sensing Data. Remote Sens. 2016, 8, 186. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Zhen, L.; Xiao, Y.; Hu, Y. Ecological and socioeconomic effects of ecological restoration in China’s Three Rivers Source Region. Sci. Total Environ. 2019, 650, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Tian, P.; Mu, X.; Jiao, J.; Wang, F.; Gao, P. Quantifying the impact of climate variability and human activities on streamflow in the middle reaches of the Yellow River basin, China. J. Hydrol. 2014, 519, 387–398. [Google Scholar] [CrossRef]
- Guan, B.; Chen, M.; Quirk, T.; Yang, S.; Shang, W.; Li, Y.; Tian, X.; Han, G. Soil seed bank and vegetation differences following channel diversion in the Yellow River Delta. Sci. Total Environ. 2019, 693, 133600. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fu, B.; Piao, S.; Lü, Y.; Ciais, P.; Feng, X.; Wang, Y. Reduced sediment transport in the Yellow River due to anthropogenic changes. Nat. Geosci. 2016, 9, 38–41. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Xiang, F.; Qin, W.; Jiang, W. Vegetation dynamics and the relations with climate change at multiple time scales in the Yangtze River and Yellow River Basin, China. Ecol. Indic. 2020, 110, 105892. [Google Scholar] [CrossRef]
- Wu, B.; Zeng, Y.; Zhao, D. Land cover mapping and above ground biomass estimation in China. In Proceedings of the 2016 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Beijing, China, 10–15 July 2016; pp. 3535–3536. [Google Scholar]
- Chen, L.; Fu, B.; Zhao, W. Source-sink landscape theory and its ecological significance. Front. Biol. China 2008, 3, 131–136. [Google Scholar] [CrossRef]
- Running, S.; Zhao, M. MOD17A3HGF MODIS/Terra Net Primary Production Gap-Filled Yearly L4 Global 500m SIN Grid V006. Available online: https://doi.org/10.5067/MODIS/MOD17A3HGF (accessed on 24 November 2021).
- Xu, H. A study on information extraction of water body with the modified normalized difference water index (MNDWI). J. Remote Sens. 2005, 5, 589–595. [Google Scholar]
- Wu, J.; Zhang, Y.; Zhang, J.; Fan, S.; Yang, C.; Zhang, X. Comparison and analysis of water indexes in muddy coasts based on MODIS data: A case study of the Yellow River Delta coast. Remote Sens. Land Resour. 2019, 3, 242–249. [Google Scholar]
- Shi, F.; Liu, S.; Sun, Y.; An, Y.; Zhao, S.; Liu, Y.; Li, M. Ecological network construction of the heterogeneous agro-pastoral areas in the upper Yellow River basin. Agric. Ecosyst. Environ. 2020, 302, 107069. [Google Scholar] [CrossRef]
- Yu, Q.; Yue, D.; Wang, J.; Zhang, Q.; Li, Y.; Yu, Y.; Chen, J.; Li, N. The optimization of urban ecological infrastructure network based on the changes of county landscape patterns: A typical case study of ecological fragile zone located at Deng Kou (Inner Mongolia). J. Clean. Prod. 2017, 163, S54–S67. [Google Scholar] [CrossRef]
- Su, K.; Yu, Q.; Yue, D.; Zhang, Q.; Yang, L.; Liu, Z.; Niu, T.; Sun, X. Simulation of a forest-grass ecological network in a typical desert oasis based on multiple scenes. Ecol. Model. 2019, 413, 108834. [Google Scholar] [CrossRef]
- Knaapen, J.P.; Scheffer, M.; Harms, B. Estimating habitat isolation in landscape planning. Landsc. Urban. Plan. 1992, 23, 1–16. [Google Scholar] [CrossRef]
- Jiang, W.; Yuan, L.; Wang, W.; Cao, R.; Zhang, Y.; Shen, W. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Koschützki, D.; Lehmann, K.A.; Peeters, L.; Richter, S.; Tenfelde-Podehl, D.; Zlotowski, O. Centrality Indices. In Network Analysis; Brandes, U., Erlebach, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 16–61. [Google Scholar]
- Jing, Y.; Baluja, S. Pagerank for product image search. In Proceeding of the 17th International Conference on World Wide Web, Beijing, China, 21–25 April 2008; pp. 307–316. [Google Scholar]
- Newman, M.E.J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.Y.; Ke, J.H.; Tang, Z.Y.; Chen, A.P. Implications and estimations of four terrestrial productivity parameters. Chin. J. Plant Ecol. 2001, 25, 414. [Google Scholar]
- Allard, V.; Soussana, J.-F.; Falcimagne, R.; Berbigier, P.; Bonnefond, J.; Ceschia, E.; D’Hour, P.; Hénault, C.; Laville, P.; Martin, C.; et al. The role of grazing management for the net biome productivity and greenhouse gas budget (CO2, N2O and CH4) of semi-natural grassland. Agric. Ecosyst. Environ. 2007, 121, 47–58. [Google Scholar] [CrossRef]
- Thornton, P.; Law, B.; Gholz, H.L.; Clark, K.L.; Falge, E.; Ellsworth, D.; Goldstein, A.; Monson, R.; Hollinger, D.; Falk, M.; et al. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric. For. Meteorol. 2002, 113, 185–222. [Google Scholar] [CrossRef]
- Ichii, K.; Hashimoto, H.; Nemani, R.; White, M. Modeling the interannual variability and trends in gross and net primary productivity of tropical forests from 1982 to 1999. Glob. Planet. Chang. 2005, 48, 274–286. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.; Yu, D.; Zhang, Y.; Binley, A. Spatial variations in soil-water carrying capacity of three typical revegetation species on the Loess Plateau, China. Agric. Ecosyst. Environ. 2019, 273, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Ruiz, S.; Maselli, F.; Chiesi, M.; Fibbi, L.; Martinez, B.; Campos-Taberner, M.; García-Haro, F.J.; Gilabert, M.A. Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain. Remote Sens. 2020, 12, 1356. [Google Scholar] [CrossRef]
- Turner, D.P.; Ritts, W.D.; Cohen, W.B.; Gower, S.T.; Running, S.W.; Zhao, M.; Costa, M.; Kirschbaum, A.A.; Ham, J.M.; Saleska, S.R.; et al. Evaluation of MODIS NPP and GPP products across multiple biomes. Remote Sens. Environ. 2006, 102, 282–292. [Google Scholar] [CrossRef]
- Hasenauer, H.; Petritsch, R.; Zhao, M.; Boisvenue, C.; Running, S.W. Reconciling satellite with ground data to estimate forest productivity at national scales. For. Ecol. Manag. 2012, 276, 196–208. [Google Scholar] [CrossRef]
- Li, X.-H.; Sun, O.J. Testing parameter sensitivities and uncertainty analysis of Biome-BGC model in simulating carbon and water fluxes in broadleaved-Korean pine forests. Chin. J. Plant Ecol. 2018, 42, 1131–1144. [Google Scholar] [CrossRef]
- Zhang, Q.B. Study on the Construction and Optimization of Ecological Network in the Northeastern Margin of Ulanbuhe Desert. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Bodin, Ö.; Alexander, S.M.; Baggio, J.; Barnes, M.; Berardo, R.; Cumming, G.S.; Dee, L.E.; Fischer, A.P.; Fischer, M.; Garcia, M.M.; et al. Improving network approaches to the study of complex social–ecological interdependencies. Nat. Sustain. 2019, 2, 551–559. [Google Scholar] [CrossRef]
- Xie, J.; Szymanski, B.K. Community detection using a neighborhood strength driven Label Propagation Algorithm. In Proceedings of the 2011 IEEE Network Science Workshop, New York, NY, USA, 22–24 June 2011. [Google Scholar]
- Goodchild, M.F. The Validity and Usefulness of Laws in Geographic Information Science and Geography. Ann. Assoc. Am. Geogr. 2004, 94, 300–303. [Google Scholar] [CrossRef] [Green Version]
- Xi, J.P. Speech at the Symposium on Ecological Protection and Quality Development of the Yellow River Basin. China Water Resour. 2019, 20, 1–3. [Google Scholar]
- Boisvenue, C.; Running, S. Simulations show decreasing carbon stocks and potential for carbon emissions for Rocky Mountain forests in the next century. Ecol. Appl. 2009, 20, 1302–1319. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.; Robledo-Abad, C.; Harper, R.; Mbow, C.; Ravindranat, N.H.; Sperling, F.; Haberl, H.; Pinto, A.D.S.; Smith, P. Co-benefits, trade-offs, barriers and policies for greenhouse gas mitigation in the agriculture, forestry and other land use (AFOLU) sector. Glob. Chang. Biol. 2014, 20, 3270–3290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Wang, J.; Sun, L.; Lv, C. Construction and optimization of green space ecological networks in urban fringe areas: A case study with the urban fringe area of Tongzhou district in Beijing. J. Clean. Prod. 2020, 276, 124266. [Google Scholar] [CrossRef]
- Nie, W.; Shi, Y.; Siaw, M.J.; Yang, F.; Wu, R.; Wu, X.; Zheng, X.; Bao, Z. Constructing and optimizing ecological network at county and town Scale: The case of Anji County, China. Ecol. Indic. 2021, 132, 108294. [Google Scholar] [CrossRef]
- Huang, X.; Wang, H.; Shan, L.; Xiao, F. Constructing and optimizing urban ecological network in the context of rapid urbanization for improving landscape connectivity. Ecol. Indic. 2021, 132, 108319. [Google Scholar] [CrossRef]
- Fu, B.; Liang, D.; Lu, N. Landscape ecology: Coupling of pattern, process, and scale. Chin. Geogr. Sci. 2011, 21, 385–391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Type | Factor | Level | Ecological Resistance Value |
|---|---|---|---|
| Topography and landform | DEM | <1000 m | 1 |
| 1000–1500 m | 3 | ||
| 1500–3000 m | 5 | ||
| 3000–4500 m | 7 | ||
| >4500 m | 9 | ||
| Slope | 0–2° | 1 | |
| 2–5° | 3 | ||
| 5–8° | 5 | ||
| 8–14° | 7 | ||
| >14° | 9 | ||
| Vegetation cover | NDVI | <0.2 | 9 |
| 0.2–0.4 | 7 | ||
| 0.4–0.6 | 5 | ||
| 0.6–0.8 | 3 | ||
| >0.8 | 1 | ||
| Hydrology | MNDWI | <0.05 | 9 |
| −0.05–0.02 | 7 | ||
| −0.02–0.02 | 5 | ||
| 0.02–0.08 | 3 | ||
| >0.08 | 1 | ||
| Density | Residential density | 0–0.08 | 1 |
| 0.08–0.24 | 3 | ||
| 0.24–0.44 | 5 | ||
| 0.44–0.66 | 7 | ||
| >0.66 | 9 | ||
| Road network | 0–0.02 | 1 | |
| Density | 0.02–0.06 | 3 | |
| 0.06–0.1 | 5 | ||
| 0.1–0.16 | 7 | ||
| >0.16 | 9 | ||
| Water network | <0.17 | 9 | |
| Density | 0.17–0.43 | 7 | |
| 0.43–0.84 | 5 | ||
| 0.84–1.53 | 3 | ||
| >1.53 | 1 | ||
| Railway network density | <0.003 | 1 | |
| 0.003–0.01 | 3 | ||
| 0.01–0.02 | 5 | ||
| 0.02–0.04 | 7 | ||
| >0.04 | 9 |
| Type of Indicator | Name of Indicator | Introduction to the Algorithm | Significance of Indicator | Reference |
|---|---|---|---|---|
| Evaluation of nodes | Degree | The number of ecological corridors owned by an ecological node | Describes the number of connections between an ecological node and other ecological nodes | [17] |
| Clustering coefficient | The ratio of the number of corridors actually existing between the neighboring nodes of a node to the maximum number of corridors that may exist | Indicates the proportion of connectivity between neighboring nodes of an ecological node | [17] | |
| Closeness centrality | The reciprocal of the sum of the shortest distances from a node to all other nodes multiplied by the number of other nodes | Indicates how close the ecological node is to other nodes through the shortest path; at the same time, it also quantifies how much the ecological node is in the geometric center of the network | [43] | |
| Betweenness centrality | The normalized index of the proportion of all the shortest paths in the network that pass through a node | Indicates the proportion or degree to which an ecological node exists on the shortest path of any two nodes in the network | [17] | |
| Eigenvector centrality | Each ecological node is assigned a relative score; connections to nodes with high scores are weighted more than connections to nodes with low scores | The more important the node connected to node A, then the more important node A is; this indicator is used to evaluate the importance of nodes | [43] | |
| PageRank | Rank of the importance of ecological nodes, evolved from eigenvector centrality | Similar to eigenvector centrality, it is an indicator used to evaluate the importance of nodes | [44] | |
| Evaluation of the network | Average degree | Average of the degrees of all ecological nodes in the network | Evaluates the average connectivity of all nodes in the network | [17] |
| Average path length | Average of the shortest distance between any two ecological nodes in the network | Indicates the smoothness of energy flow of the whole ecospatial network | [17] | |
| Average clustering coefficient | Average of the clustering coefficients of all nodes in the network | Indicates whether the distribution of ecological nodes in the network tends to be concentrated or decentralized | [17] | |
| Modularity | Each ecological node in the network is assigned to a different community | Evaluates the effect of network community division | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, M.; Si, G.; Yu, Q.; Huang, H.; Huang, Y.; Liu, W.; Guo, H. Study on the Relationship between Topological Characteristics of Vegetation Ecospatial Network and Carbon Sequestration Capacity in the Yellow River Basin, China. Remote Sens. 2021, 13, 4926. https://doi.org/10.3390/rs13234926
Fang M, Si G, Yu Q, Huang H, Huang Y, Liu W, Guo H. Study on the Relationship between Topological Characteristics of Vegetation Ecospatial Network and Carbon Sequestration Capacity in the Yellow River Basin, China. Remote Sensing. 2021; 13(23):4926. https://doi.org/10.3390/rs13234926
Chicago/Turabian StyleFang, Minzhe, Guoxin Si, Qiang Yu, Huaguo Huang, Yuan Huang, Wei Liu, and Hongqiong Guo. 2021. "Study on the Relationship between Topological Characteristics of Vegetation Ecospatial Network and Carbon Sequestration Capacity in the Yellow River Basin, China" Remote Sensing 13, no. 23: 4926. https://doi.org/10.3390/rs13234926