

Edge Effects in Amazon Forests: Integrating Remote Sensing and Modelling to Assess Changes in Biomass and Productivity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Degree of Fragmentation from DLR TanDEM-X
2.3. Lidar Data from NASA GEDI
2.4. Individual-Based Forest Model FORMIND
2.5. Comparison with Other Satellite Data
3. Results
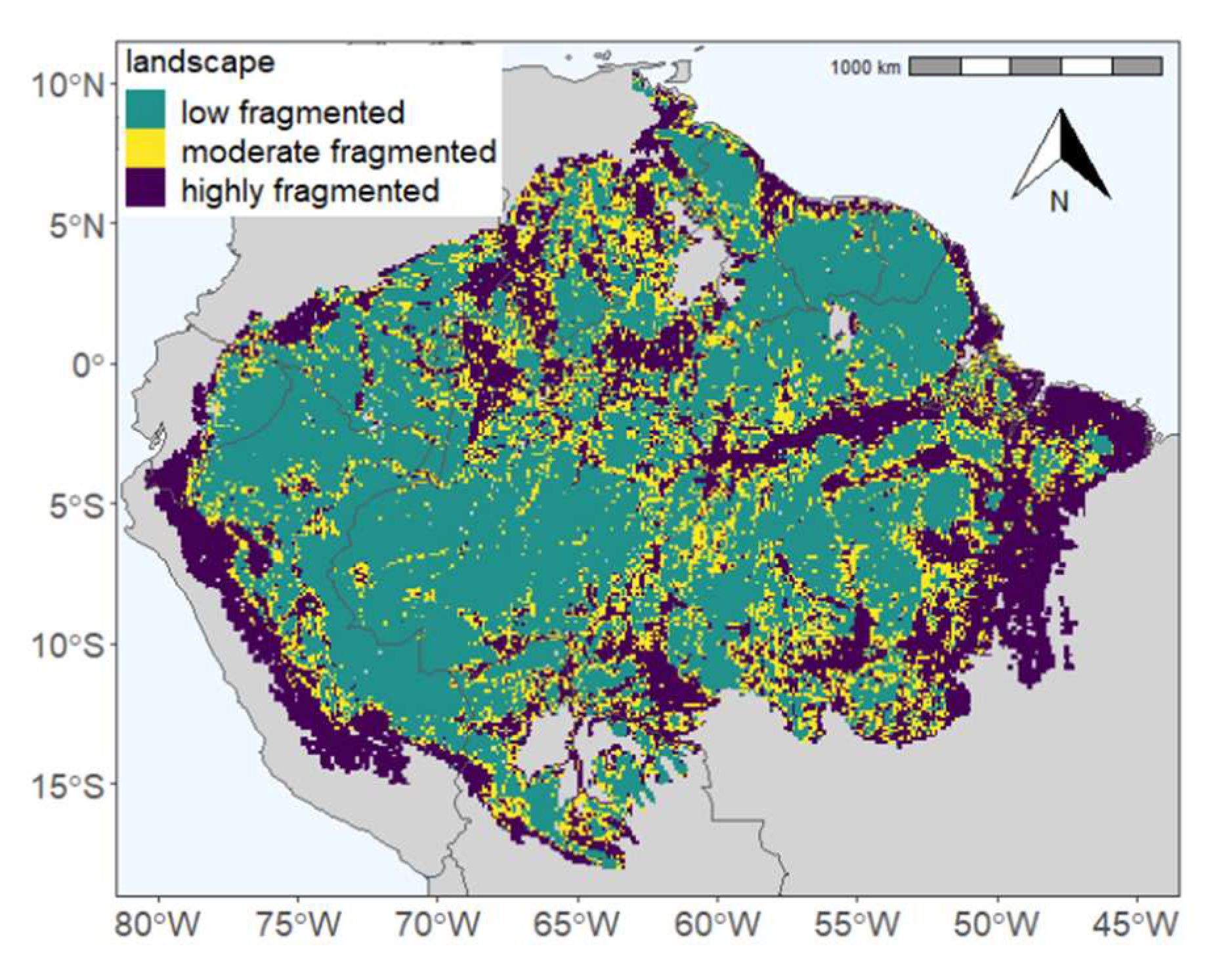
3.1. The Current State of Forest Distances in the Amazon
3.2. Impact of Forest Fragmentation on Amazon Rainforest at Forest Stand Level
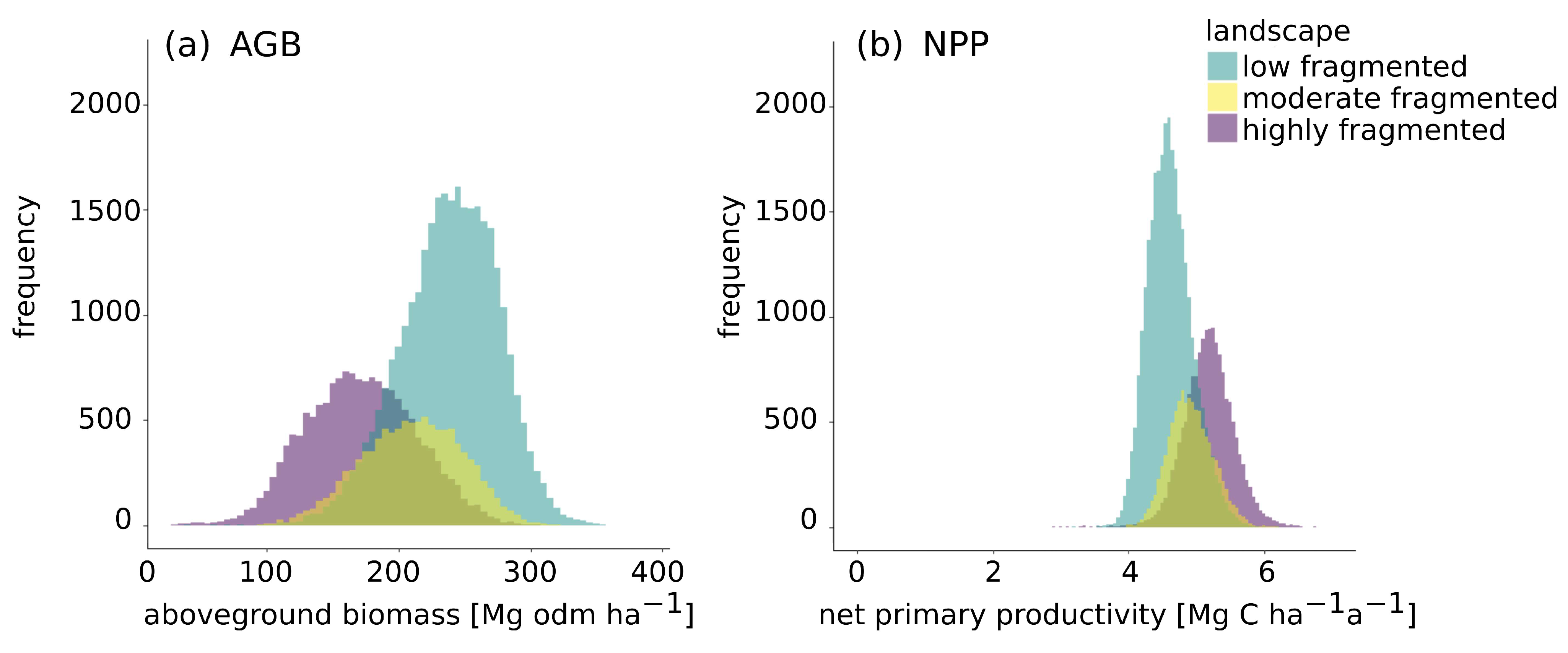
3.3. Impact of Forest Fragmentation on Amazon Biomass and Productivity at Landscape Level
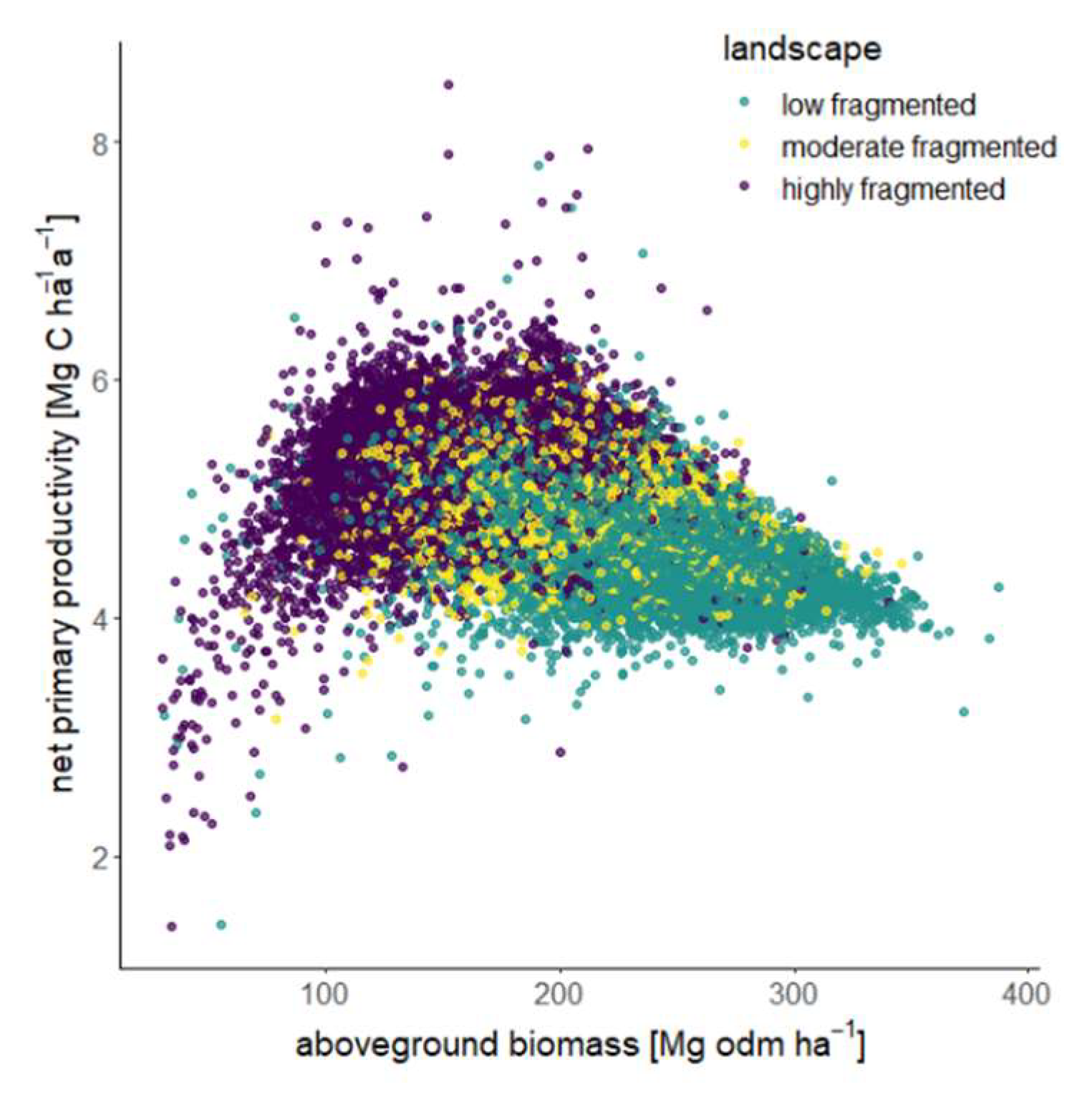
3.4. The Relationship between Forest Properties in Fragmented Landscapes
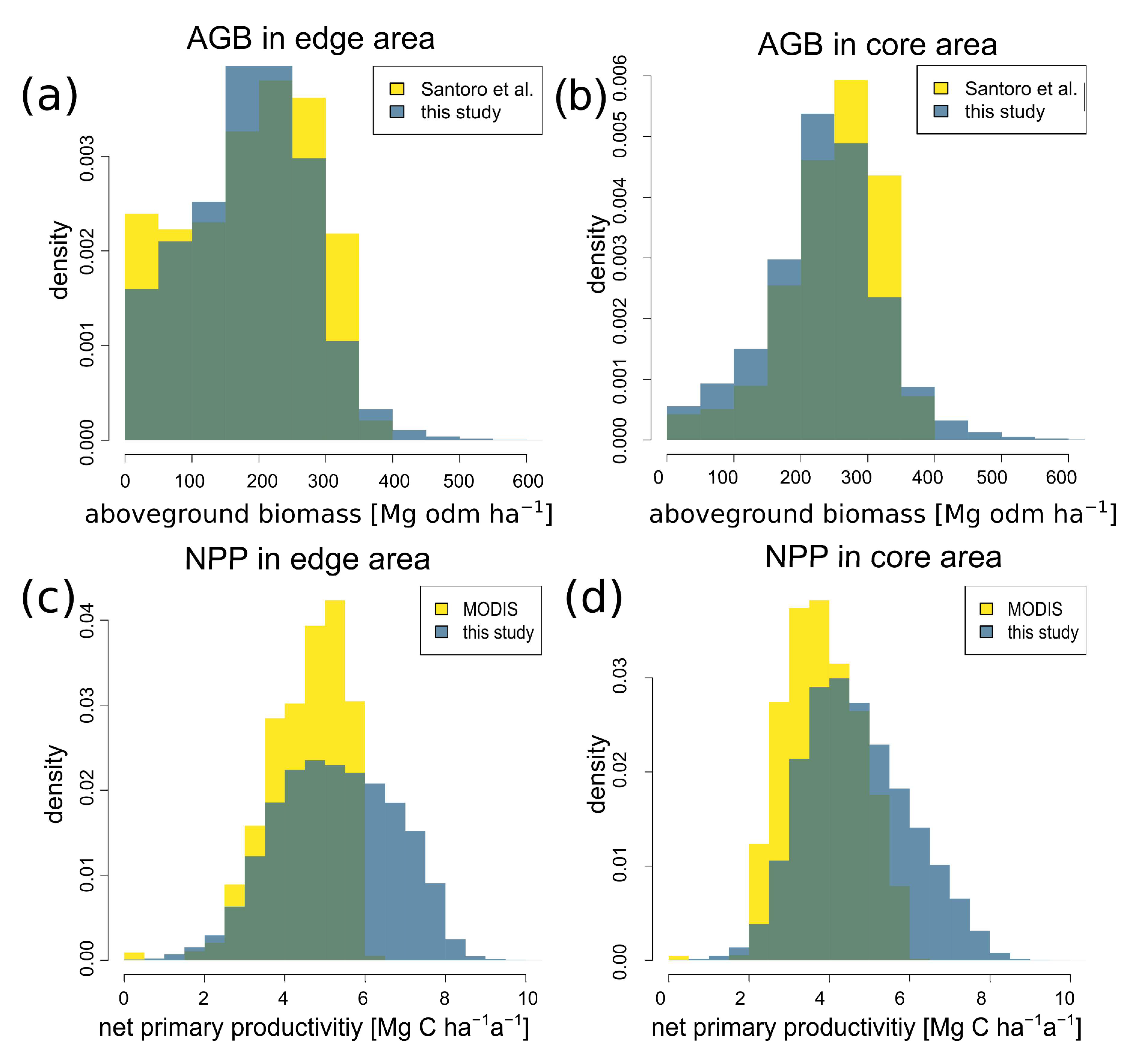
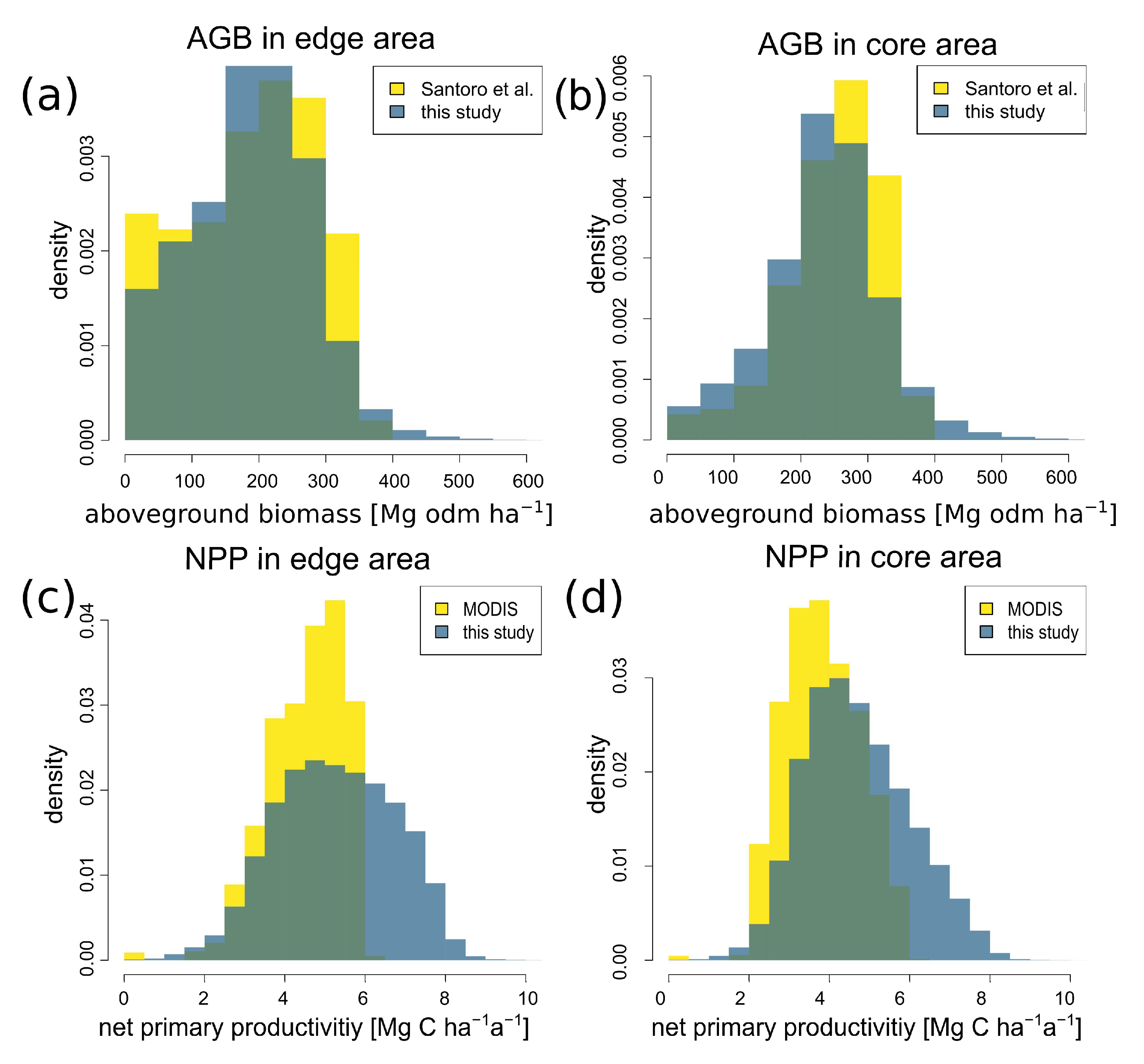
3.5. Comparison of Biomass and Productivity with Other Satellite Products
4. Discussion
4.1. Summary
4.2. Impact of Fragmentation on Edge and Core Forests
4.3. Fragmentation at the Landscape Scale
4.4. Challenges in Combining Remote Sensing and Forest Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pütz, S.; Groeneveld, J.; Henle, K.; Knogge, C.; Martensen, A.C.; Metz, M.; Metzger, J.P.; Ribeiro, M.C.; Dantas de Paula, M.; Huth, A. Long-term carbon loss in fragmented Neotropical forests. Nat. Commun. 2014, 5, 5037. [Google Scholar] [CrossRef] [PubMed]
- Gatti, L.V.; Gloor, M.; Miller, J.B.; Doughty, C.E.; Malhi, Y.; Domingues, L.G.; Basso, L.S.; Martinewski, A.; Correia, C.S.C.; Borges, V.F.; et al. Drought sensitivity of Amazonian carbon balance revealed by atmospheric measurements. Nature 2014, 506, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.; Knapp, N.; Fischer, R. Mapping Amazon forest productivity by fusing GEDI lidar waveforms with an individual-based forest model. Remote Sens. 2021, 13, 4540. [Google Scholar] [CrossRef]
- Brinck, K.; Fischer, R.; Groeneveld, J.; Lehmann, S.; Dantas De Paula, M.; Pütz, S.; Sexton, J.O.; Song, D.X.; Huth, A. High resolution analysis of tropical forest fragmentation and its impact on the global carbon cycle. Nat. Commun. 2017, 8, 14855. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Taubert, F.; Müller, M.S.; Groeneveld, J.; Lehmann, S.; Wiegand, T.; Huth, A. Accelerated forest fragmentation leads to critical increase in tropical forest edge area. Sci. Adv. 2021, 7, eabg7012. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Houghton, R.A.; Skole, D.L.; Nobre, C.A.; Hackler, J.L.; Lawrence, K.T.; Chomentowski, W.H. Annual fluxes or carbon from deforestation and regrowth in the Brazilian Amazon. Nature 2000, 403, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Lapola, D.M.; Pinho, P.; Barlow, J.; Aragão, L.E.O.C.; Berenguer, E.; Carmenta, R.; Liddy, H.M.; Seixas, H.; Silva, C.V.J.; Silva-Junior, C.H.L.; et al. The drivers and impacts of Amazon forest degradation. Science 2023, 379, eabp8622. [Google Scholar] [CrossRef]
- Numata, I.; Cochrane, M.A.; Souza, C.M.; Sales, M.H. Carbon emissions from deforestation and forest fragmentation in the Brazilian Amazon. Environ. Res. Lett. 2011, 6, 044003. [Google Scholar] [CrossRef]
- Laurance, W.F.; Camargo, J.L.C.; Luizao, R.C.C.; Laurance, S.G.; Pimm, S.L.; Bruna, E.M.; Stouffer, P.C.; Williamson, G.B.; Benitez-Malvido, J.; Vasconcelos, H.L.; et al. The fate of Amazonian forest fragments: A 32-year investigation. Biol. Conserv. 2011, 144, 56–67. [Google Scholar] [CrossRef]
- Qie, L.; Lewis, S.L.; Sullivan, M.J.P.; Lopez-Gonzalez, G.; Pickavance, G.C.; Sunderland, T.; Ashton, P.; Hubau, W.; Abu Salim, K.; Aiba, S.-I.; et al. Long-term carbon sink in Borneo’s forests halted by drought and vulnerable to edge effects. Nat. Commun. 2017, 8, 1966. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F.; Camargo, J.L.C.; Fearnside, P.M.; Lovejoy, T.E.; Williamson, G.B.; Mesquita, R.C.G.; Meyer, C.F.J.; Bobrowiec, P.E.D.; Laurance, S.G.W. An Amazonian rainforest and its fragments as a laboratory of global change. Biol. Rev. 2018, 93, 223–247. [Google Scholar] [CrossRef] [PubMed]
- Chaplin-Kramer, R.; Ramler, I.; Sharp, R.; Haddad, N.M.; Gerber, J.S.; West, P.C.; Mandle, L.; Engstrom, P.; Baccini, A.; Sim, S.; et al. Degradation in carbon stocks near tropical forest edges. Nat. Commun. 2015, 6, 10158. [Google Scholar] [CrossRef] [PubMed]
- Dubayah, R.; Blair, J.B.; Goetz, S.; Fatoyinbo, L.; Hansen, M.; Healey, S.; Hofton, M.; Hurtt, G.; Kellner, J.; Luthcke, S.; et al. The Global Ecosystem Dynamics Investigation: High-resolution laser ranging of the Earth’s forests and topography. Sci. Remote Sens. 2020, 1, 100002. [Google Scholar] [CrossRef]
- Rödig, E.; Cuntz, M.; Rammig, A.; Fischer, R.; Taubert, F.; Huth, A. The importance of forest structure for carbon fluxes of the Amazon rainforest. Environ. Res. Lett. 2018, 13, 054013. [Google Scholar] [CrossRef]
- Rödig, E.; Knapp, N.; Fischer, R.; Bohn, F.J.; Dubayah, R.; Tang, H.; Huth, A. From small-scale forest structure to Amazon-wide carbon estimates. Nat. Commun. 2019, 10, 5088. [Google Scholar] [CrossRef] [PubMed]
- Martone, M.; Rizzoli, P.; Wecklich, C.; González, C.; Bueso-Bello, J.-L.; Valdo, P.; Schulze, D.; Zink, M.; Krieger, G.; Moreira, A. The global forest/non-forest map from TanDEM-X interferometric SAR data. Remote Sens. Environ. 2018, 205, 352–373. [Google Scholar] [CrossRef]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Krieger, G.; Moreira, A.; Fiedler, H.; Hajnsek, I.; Werner, M.; Younis, M.; Zink, M. TanDEM-X: A satellite formation for high-resolution SAR interferometry. IEEE Trans. Geosci. Remote Sens. 2007, 45, 3317–3341. [Google Scholar] [CrossRef]
- Bueso Bello, J.L.; González, C.; Martone, M.; Rizzoli, P. TanDEM-X Forest/Non-Forest Map Product Description; DLR Microwaves and Radar Institute: Köln, Germany, 2019. [Google Scholar]
- Hoshen, J.; Klymko, P.; Kopelman, R. Percolation and cluster distribution. III. Algorithms for the site-bond problem. J. Stat. Phys. 1979, 21, 583–600. [Google Scholar] [CrossRef]
- Taubert, F.; Fischer, R.; Groeneveld, J.; Lehmann, S.; Müller, M.S.; Rödig, E.; Wiegand, T.; Huth, A. Global patterns of tropical forest fragmentation. Nature 2018, 554, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Wirt, B.; Armston, J.; Hofton, M.; Luthke, S.; Tang, H. GLOBAL Ecosystem Dynamics Investigation (GEDI) Level 2 User Guide for SDPS PGEVersion 3 (P003) of GEDI L2A Data and SDPS PGEVersion 3 (P003) of GEDI L2B Data Version 2.0 April 2021; GEDI Ecosystem Lidar: Reston, VA, USA, 2021; p. 25. [Google Scholar]
- Fischer, R.; Bohn, F.; Dantas de Paula, M.; Dislich, C.; Groeneveld, J.; Gutiérrez, A.G.; Kazmierczak, M.; Knapp, N.; Lehmann, S.; Paulick, S.; et al. Lessons learned from applying a forest gap model to understand ecosystem and carbon dynamics of complex tropical forests. Ecol. Model. 2016, 326, 124–133. [Google Scholar] [CrossRef]
- Köhler, P.; Huth, A. The effects of tree species grouping in tropical rainforest modelling: Simulations with the individual-based model FORMIND. Ecol. Model. 1998, 109, 301–321. [Google Scholar] [CrossRef]
- Rödig, E.; Cuntz, M.; Heinke, J.; Rammig, A.; Huth, A. Spatial heterogeneity of biomass and forest structure of the Amazon rain forest: Linking remote sensing, forest modelling and field inventory. Glob. Ecol. Biogeogr. 2017, 26, 1292–1302. [Google Scholar] [CrossRef]
- Bohn, F.J.; Frank, K.; Huth, A. Of climate and its resulting tree growth: Simulating the productivity of temperate forests. Ecol. Model. 2014, 278, 9–17. [Google Scholar] [CrossRef]
- Henniger, H.; Huth, A.; Frank, K.; Bohn, F.J. Creating virtual forests around the globe and analysing their state space. Ecol. Model. 2023, 483, 110404. [Google Scholar] [CrossRef]
- Smith, T.M.; Shugart, H.H.; Woodward, F.I.; Burton, P.J. Plant functional types. In Vegetation Dynamics & Global Change; Springer: Boston, MA, USA, 1993; pp. 272–292. [Google Scholar]
- Fischer, R.; Rödig, E.; Huth, A. Consequences of a reduced number of plant functional types for the simulation of forest productivity. Forests 2018, 9, 460. [Google Scholar] [CrossRef]
- Fischer, R.; Knapp, N.; Bohn, F.; Shugart, H.H.; Huth, A. The relevance of forest structure for biomass and productivity in temperate forests: New perspectives for remote sensing. Surv. Geophys. 2019, 40, 709–734. [Google Scholar] [CrossRef]
- Santoro, M.; Cartus, O.; Carvalhais, N.; Rozendaal, D.M.A.; Avitabile, V.; Araza, A.; de Bruin, S.; Herold, M.; Quegan, S.; Rodríguez-Veiga, P.; et al. The global forest above-ground biomass pool for 2010 estimated from high-resolution satellite observations. Earth Syst. Sci. Data 2021, 13, 3927–3950. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.; Heinsch, F.A.; Nemani, R. MODIS-derived terrestrial primary production. In Land Remote Sensing and Global Environmental Change; NASA’s Earth observing system and the science of ASTER and MODIS; Springer: New York, NY, USA, 2011; pp. 635–660. [Google Scholar]
- Park, J.H.; Gan, J.; Park, C. Discrepancies between global forest net primary productivity estimates derived from MODIS and forest inventory data and underlying factors. Remote Sens. 2021, 13, 1441. [Google Scholar] [CrossRef]
- Laurance, W.F.; Bierregaard, R.O. (Eds.) Tropical Forest Remnants. Ecology, Management, and Conservation of Fragmented Communities; Cambridge University Press: Chicago, IL, USA, 1997; p. 616. [Google Scholar]
- Laurance, W.F.; Delamonica, P.; Laurance, S.G.; Vasconcelos, H.L.; Lovejoy, T.E. Conservation—Rainforest fragmentation kills big trees. Nature 2000, 404, 836. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.H.; Vaz, M.C.; Camargo, J.L.C.; Laurance, W.F.; de Andrade, A.; Vicentini, A.; Laurance, S.; Raumonen, P.; Jackson, T.; Zuquim, G.; et al. Edge effects on tree architecture exacerbate biomass loss of fragmented Amazonian forests. Nat. Commun. 2023, 14, 8129. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Laurance, W.F.; Nascimento, H.E.M.; Laurance, S.G.; Andrade, A.C.A.; Fearnside, P.M.; Ribeiro, J.E.L.; Capretz, R.L. Rain forest fragmentation and the proliferation of successional trees. Ecology 2006, 87, 469–482. [Google Scholar] [CrossRef]
- Laurance, W.F. Hyperdynamism in fragmented habitats. J. Veg. Sci. 2002, 13, 595–602. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K.; Pearse, W.D.; Lefebvre, V.; Rosa, I.M.D.; Carreiras, J.M.B.; Lucas, R.M.; Reuman, D.C. Using landscape history to predict biodiversity patterns in fragmented landscapes. Ecol. Lett. 2013, 16, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Numata, I.; Silva, S.S.; Cochrane, M.A.; d’Oliveira, M.V.N. Fire and edge effects in a fragmented tropical forest landscape in the southwestern Amazon. For. Ecol. Manag. 2017, 401, 135–146. [Google Scholar] [CrossRef]
- Caldararu, S.; Palmer, P.I.; Purves, D.W. Inferring Amazon leaf demography from satellite observations of leaf area index. Biogeosciences 2012, 9, 1389–1404. [Google Scholar] [CrossRef]
- Knapp, N.; Huth, A.; Kugler, F.; Papathanassiou, K.; Condit, R.; Hubbell, S.P.; Fischer, R. Model-assisted estimation of tropical forest biomass change: A comparison of approaches. Remote Sens. 2018, 10, 731. [Google Scholar] [CrossRef]
- Smith, C.C.; Barlow, J.; Healey, J.R.; de Sousa Miranda, L.; Young, P.J.; Schwartz, N.B. Amazonian secondary forests are greatly reducing fragmentation and edge exposure in old-growth forests. Environ. Res. Lett. 2023, 18, 124016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Edge (CV) | Core (CV) | Difference between Core and Edge Values | |
|---|---|---|---|
| Aboveground biomass [Mg odm ha−1] | 172 ± 86 (50%) | 235 ± 85 (36%) | −27% |
| Net primary productivity [Mg C ha−1 a−1] | 5.4 ± 1.5 (29%) | 4.7 ± 1.3 (28%) | +13% |
| Gross primary productivity [Mg C ha−1 a−1] | 22 ± 7 (31%) | 24 ± 6 (23%) | −8% |
| Leaf area index [-] | 3.9 ± 1.4 (35%) | 4.6 ± 1.1 (25%) | −15% |
| Highly Fragmented (CV) | Moderate Fragmented (CV) | Low Fragmented (CV) | |
|---|---|---|---|
| Mean aboveground biomass [Mg odm ha−1] | 171 ± 42 (24%) | 210 ± 38 (18%) | 238 ± 37 (16%) |
| Mean net primary productivity [Mg C ha−1 a−1] | 5.2 ± 0.4 (7.6%) | 4.9 ± 0.3 (6.7%) | 4.6 ± 0.3 (7.2%) |
| Mean gross primary productivity [Mg C ha−1 a−1] | 22 ± 3 (14%) | 23 ± 2 (10%) | 24 ± 2 (8%) |
| Mean leaf area index | 3.8 ± 0.6 (16.3%) | 4.3 ± 0.5 (12.5%) | 4.6 ± 0.5 (11.2%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, L.; Huth, A.; Bogdanowski, A.; Müller, M.; Fischer, R. Edge Effects in Amazon Forests: Integrating Remote Sensing and Modelling to Assess Changes in Biomass and Productivity. Remote Sens. 2024, 16, 501. https://doi.org/10.3390/rs16030501
Bauer L, Huth A, Bogdanowski A, Müller M, Fischer R. Edge Effects in Amazon Forests: Integrating Remote Sensing and Modelling to Assess Changes in Biomass and Productivity. Remote Sensing. 2024; 16(3):501. https://doi.org/10.3390/rs16030501
Chicago/Turabian StyleBauer, Luise, Andreas Huth, André Bogdanowski, Michael Müller, and Rico Fischer. 2024. "Edge Effects in Amazon Forests: Integrating Remote Sensing and Modelling to Assess Changes in Biomass and Productivity" Remote Sensing 16, no. 3: 501. https://doi.org/10.3390/rs16030501