1. Introduction
Switchgrass (
Panicum virgatum L.), a native North American warm-season C4 perennial grass has been identified as a potential biofuel feedstock with a promise for production across diverse climates in North America [
1–
3]. Switchgrass is adapted to a wide range of climatic and edaphic conditions from northern Mexico to southern Canada, and from the Atlantic coast to the Rocky Mountains [
3]. It is classified as a dedicated biofuel feedstock due to its high level of productivity over long-term (>10 yr) across varied environmental conditions [
4], suitability for production on marginal land [
3], low nutrient requirements [
1,
5], and positive environmental benefits such as reduced erosion, increased water quality, enhanced soil-carbon sequestration, wildlife habitat and reducing greenhouse gas emissions [
2].
Two major types of switchgrass are found in North America: the low land ecotype is exclusively tetraploid, associated with wet conditions and better adapted to lower latitudes, while the upland ecotype is mainly tetraploid or octaploid, associated with dry conditions and better adapted for mid to northern latitudes [
6,
7]. Because of these distinct differences between and within the upland and lowland ecotypes, it is important to be able to discriminate switchgrass plants. Current methods of identification of specific cultivars are limited to genomic analysis and visual discrimination. Despite genomics analysis only requiring a small amount of sample material, expensive equipment and expertise are needed to make the assessment. While, visual discrimination is possible with trained personnel, results among personnel and locations can vary due to plants of different age and localized effect of light, temperature and moisture.
Remote sensing is a well-known non-destructive method that can play a critical role as a crop stress assessment tool, monitoring nutrient status, disease and weed and insect infestation. The basic concept of remote sensing is the ability to quantify variations due to space and size (spatial variations), variations in reflected or emitted radiation (spectral variations) and variations of reflected or emitted radiation, space and size over time (temporal variations) [
8]. Radiation reflected by vegetation varies in different part of the spectrum due to the vegetation biophysical characteristics. In the visible part of the spectrum (400–700 nm) the amount of reflected or emitted radiation is controlled by the plant pigmentation the chlorophyll, carotenes and xanthophylls [
8]. In the near infrared portion of the spectrum (700–1,350 nm) reflected or emitted radiation is controlled by the internal leaf structures. The middle infrared (Mid-IR) portion of the spectrum (1,350–2,500 nm) reflected or emitted radiation is controlled primarily by
in vivo water content and secondarily by internal leaf structures [
8]. The importance of these parts of the spectrum (Mid-IR) is the high resolution spectral response that is often observed for a crop at leaf or canopy level at different stages of development. As a plant develops and interacts with environmental conditions, reflectance from these areas of the spectrum is affected. Reflectance tends to increase in the near-infrared (NIR; 725–900 nm) as the internal leaf structure of most plant species (
i.e., cotton canopy) reflects more of the energy in this portion, and changes in green peak (550 nm) and in red region (650–690 nm) due to chlorophyll reflectance and adsorption respectively [
9].
The recent advances in ground-based high resolution multispectral, hyperspectral digital cameras, spectroradiometers and several other optical sensors can play a critical role towards a more intelligent crop production system. Narrowbands located in specific portions of the spectrum have been shown to significantly improve discrimination capabilities and classification accuracies for various vegetation and agricultural crops when compared to broadbands such as Landsat Thematic Mapper ™ and Systeme Pour L’Observation de la Terre (SPOT) [
10]. Hyperspectral narrowbands and vegetative indices developed from them are capable of detecting small differences in percentage green cover [
11], crop moisture variations [
12] and discriminating among varieties [
13,
14]. Despite, the improvement of narrowbands over broadband, the large number of bands available with hyperspectral sensors makes analysis complex and time consuming [
14]. Several approaches were used including reflectance from individual narrowbands, various ratios and indices, and multivariate statistical analysis to discriminate among varieties. The use of high resolution hyperspectral leaf reflectance with pigment profiles to discriminate among sugarcane varieties was investigated [
13]. The hyperspectral reflectance data was collected at 350–800 nm at 0.4 nm intervals from the third youngest fully open leaf and plant pigment analysis was done from the same leaf. The authors reported that several single wavelengths ranging from 560 to 720 nm were able to discriminate between selected varieties, multivariate analysis resulted in a 95–100% correct classification for all varieties with leaf reflectance data in comparison to 76–81% correct classification with plant pigment data and 81–86% using vegetative indices [NDVI (Normalize Difference Vegetative Index) and WDRVI (Wide Dynamic Range Vegetative Index)]. Hyperspectral narrowband wavelengths from 375 to 1,075 nm and multiple discriminant analysis were used by Ray
et al. [
14] to identify nine bands (520, 560, 660, 690, 730, 760, 780, 790 and 800 nm) and vegetative indices, simple ratio, ZTM (Zarco Tejada and Miller), Red edge 750/700 and Red edge 740/720 for discriminating among four potato varieties. Likewise, Hatfield and Prueger [
15] used different vegetative indices to quantify differences among varieties of corn and soybean at different growth stages during the growing season. The authors concluded that the ability to quantify differences among the varieties and crops was a function of growth stage and vegetative index.
The use of hyperspectral remote sensing techniques, with high spectral resolutions, in combination with plant pigment analysis may significantly improve the ability to discriminate between and among switchgrass cultivars and ecotypes. The dominant plant pigments are the chlorophylls. These compounds exhibit pronounced absorption in the bluish (400–500 nm) and reddish (600–700 nm) wavelengths of the magnetic spectrum. Other plant pigments such as carotenoids produces yellow or orange reflectance centered at about 450 nm wavelength of the spectrum. Knowledge of pigment pools including those associated with UV-B absorption could improve our understanding of plant stress responses to light, temperature and water, and could also be used to discriminate between species, cultivars and varieties of switchgrass. The objective of this study was to compare the use of hyperspectral narrowbands, hyperspectral narrowband vegetation indices and leaf pigmentation (Chlorophyll a, Chlorophyll b, Carotenoids) to discriminate between 12 switchgrass cultivars and five nitrogen treatments for one of the cultivars (Alamo) at different times during the growing season.
2. Materials and Methods
2.1. Experimental Design
2.1.1. Switchgrass Cultivars
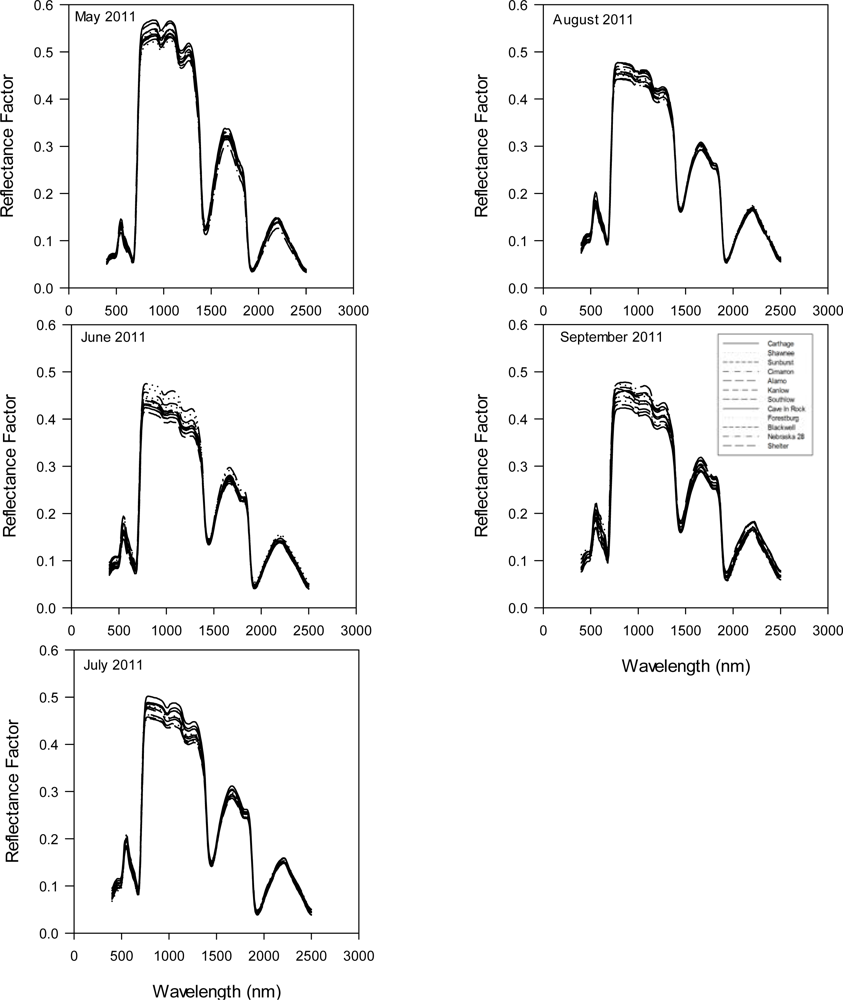
An experiment consisting of twelve cultivars of switchgrass (
Figure 1) with known difference in ecotype, and origin (
Table 1) was established at the Stillwater Agronomy Research Station (36.12°N, 97.09°W) in April 2009 to evaluate biomass yield production among the cultivars. Switchgrass cultivars were planted by seed in plots (6.10 m wide × 7.62 m long) in a randomized complete block design with three replications. Plots were seeded at a rate of 5.04 kg·ha
−1 of pure live seed using a no-till planter. Leaf samples were collected on 24 May, 20 June, 25 July, 24 August and 30 September 2011 from which the spectral and pigment data was obtained (
Table 2).
2.1.2. Nitrogen Treatments
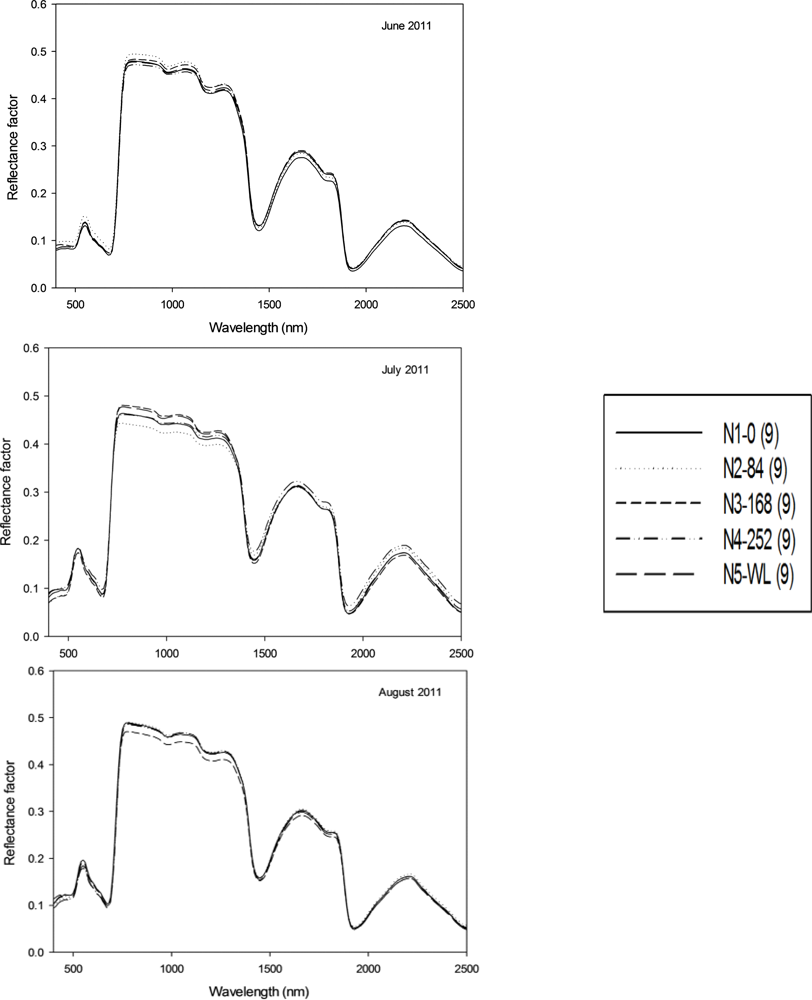
An experiment consisting of five nitrogen treatments (winter legume (hairy vetch), 0, 84, 168 and 252 kg·N·ha
−1) was established at the Stillwater Agronomy Research Station (EFAW Site, 36.13°N, 97.10°W) in a one year old established stand of switchgrass “Alamo” to evaluate the effect of nitrogen treatment on biomass production. Experimental design is a randomized complete block and replicated three times. No nitrogen fertilizer was applied in the establishment year. Plots were fertilized with the different rates of N on 3 June 2011. Leaf samples were collected on 17 June, 27 July and 27 August 2011 from which the spectral and pigment data was obtained (
Table 2).
2.2. Leaf Sampling
Top most fully expanded leaf (6th or 5th) was excised from 6 random plants in each plot and sealed in a plastic bag in an ice chest and transported to the laboratory for spectral and pigment measurements. These samples were collected between 10:00 and 15:00 h local time.
2.3. Spectral Data
Hyperspectral reflectance data was collected using an ASD Field Spec Pro spectroradiometer (Analytical Spectral Devices Inc., Boulder, CO, USA) that consisted of a spectral range of 350–2,500 nm and a 25° field of view. The spectrometer is equipped with three sensors [(visible (400–750 nm) and near infrared-NIR (750–1,100 nm), shortwave infrared-SWIR1 (1,000–1,800 nm) and SWIR2 (1,800–2,500 nm)] with spectral sampling of 3, 10 and 10 nm, respectively. The instrument was periodically calibrated using a standard Spectralon white reference panel (Labsphere Inc., North Sutton, NH, USA). The white reference was measured at 15 min intervals to check the instrument stability for 100% reflectance. To measure leaf reflectance, two leaves were place beside each other to provide a large enough surface area, and sandwiched between the non-reflecting, black body and the light probe. This ensured that no extraneous light entered the sensor during these measurements. Care was taken in placing the leaves beside each other, to ensure that no space or overlapping occurred. Three replicated measurements were made on leaves collected from each plot. Built-in spectral resolution output of the data from the ASD operating system is 1 nm along the whole spectrum. To reduce the amount of data for analysis, spectral data were averaged at 10-nm wavelength intervals (e.g., a band center at 400 was the averaged value between 395–405 nm) giving a total of 211 spectral bands between 400–2,500 nm. Spectral data at start of spectrum due to noise (350–395 nm) and in the atmospheric water absorption spectral regions (1,350–1,420 and 1,800–1,960 nm) were deleted from the data before analysis leaving 186 spectral bands for analysis.
2.4. Pigment Analysis
After reflectance measurements, five of the leaves used for hyperspectral measurements were sampled for plant pigment analysis. The photosynthetic pigments (Chlorophyll a, Chlorophyll b and Carotenoids) were extracted by placing five 38.5 mm
2 leaf discs in a vial with 5 mL of dimethyl sulfoxide and extracting after incubating in a dark room for 24 h. The absorption of the extracts was determined at 664, 648 and 470 nm using the spectrophotometer. The equations by Lichtenthaler [
16] were used to derive the pigment concentrations.
The UV-B absorbing compounds were determined using methods described in Kakani
et al. [
17]. UV-B absorbing compounds were extracted from placing five 38.5 mm
2 leaf discs in a vial with 10 mL of aliquot consisting of a methanol, water and hydrochloric acid in the proportion of 79:20:1 ratio. The vials were incubated at room temperature for 24 h in dark to allow for complete extraction of UV-B absorbing compounds. The absorbance of the extracts from the different cultivars was measured at 330 nm. The content of UV-B absorbing compounds was calculated using the equation [
17], C = 16.05 × A, where A is absorbance at 330 nm and C is concentration of UV-B absorbing compound (μg·mL
−1 of extract).
2.5. Data Analysis
The optimal wavebands that were able to discriminate the target as affected by time of collection were determined based on a comprehensive analysis using principal component analysis (PCA). The goal of the PCA is to identify underlying variables, or factors that explain the pattern of correlations within a set of observed variables. The PCA tends to achieve this by deriving a new set of uncorrelated variables called principal components, thereby reducing the number of variables. The PCA was carried out using the PRINCOMP procedure in SAS [
18].
To evaluate the effect of time of collection in differentiating among the cultivars and N treatments a linear discriminant analysis with cross validation was done for each month. Discriminant function analysis (DA) is a qualitative tool often used to discriminate between two or more groups. To classify observations into a group, a mathematical rule or discriminant function is used to determine to which group an observation belongs based on knowledge of the quantitative variables only. In this study, DA was used to classify the twelve cultivars and five N treatments, by computing a sample’s distance from each class center in Mahalanobis distance (MD) units [
19]. The MD is the parameter that is calculated and used to determine how close to the center of its group is an individual spectrum sample. The MD was calculated using the following equation [
19]:
where
denotes the MD between the cultivars
i and
j,
cov−1denotes the inverse covariance matrix, and
Av(
xi) and
Av(
xi) denote the mean reflection for cultivars
i and
j, respectively. The smallest MD is used to pick the group that the individual fits best. The equation assumes a common variance for the populations from which the groups are derived. Discriminant function analysis was carried out on the first five PCAs resulting from the PCA as they covered most of the variation (99% of variation explained) contained in the raw spectral data.
Selected hyperspectral narrowband vegetation indices that take into account leaf structure, pigmentation and red edge characteristics were computed for each set of spectral data. The vegetation indices computed are shown in
Table 3. Stepwise discriminant analysis (SDA) was carried out to find the best indices which can differentiate switchgrass cultivars and nitrogen treatments at each sampling interval. The SDA is a procedure that reduces the data set to those variables that maximize between statistical group variability while minimizing within group variability. The difference between PCA and SDA is that the PCA creates a new set of uncorrelated variables that defines the axes of greatest variability in the data, while SDA identifies from among the original variables, the best variables that describes differences between given groups. The Wilk’s lambda statistics was used to select the best vegetation indices for differentiating the cultivars and N treatments. Low Wilk’s Lambda valve suggests a great degree of separation. Therefore, index with the lowest Wilk’s lambda value resulted in the greatest separation among the cultivars and N treatments.
Similarly, PCA and DA were performed at each sampling date for the pigment content to determine degree of discrimination. Discriminant analysis was carried out on the two first PCAs as they contain most of the variation (99% of variation explained). The results were compared to determine the approach and the sampling interval that provided the greatest separation. All statistical analyses were performed using SAS (Statistical Analysis System) [
18].
4. Discussion
Optimal wavebands are those bands that have the least correlation among them, high information content and are able to discriminate the target. Currently there is no best approach available to determine the optimal number of bands required to discriminate vegetation characteristics [
10]. Researchers in the past have used various approaches from incorporating reflectance from individual narrowbands, various indices derivatives of reflectance spectra, or combinations of these. Using discriminant analysis of reflectance data resulted with correctly grouping the twelve cultivars into their respective grouping with an accuracy of 80% using cross-validation and 100% using re-substitution methods for the September sampling. Pigment data was unable to discriminate among the cultivars. However, in discriminating among N treatments pigment data was found to be better than the spectral data. Likewise, Johnson
et al. [
13] found greater accuracy (95–100%) in classifying sugarcane varieties with leaf spectral reflectance data in comparison to 76–81% accuracy with plant pigment data. The ability to use spectral reflectance data obtained from a spectroradiometer to discriminate or identify plant varieties or cultivars is based on the leaf spectral characteristics that are related to the leaf pigment profile and structure. The leaf spectral characteristics of plants is affected by many factors such as plant species, leaf maturity, microclimate position of the leaf on the plant [
37], environmental condition in which plant is grown and time of data collection. The amount of light reflected, absorbed or transmitted in the visible (400–700 nm), near infrared (700–1,350 nm) and middle-infrared (1,350–2,500 nm) is primarily controlled by the leaf pigment profile, internal leaf structure, and
in vivo water content respectively [
8]. The PCA found middle infrared to be the dominant waveband for PC1 explaining 63% of the variability, NIR wavebands in May and September, and middle infrared wavebands in June, July and August to be the dominant wavebands for PC2 explaining 22% variability and red wavebands in May, July and August and green wavebands in June and September for PC3 explaining 11% of the variability. Thenkabail
et al. [
10] also found middle infrared wavebands to be the dominant waveband for PC 1 and accounting for a similar 62% of the variability. The middle infrared region of the spectrum dominated the PC1 with a 63% frequency of occurrence suggests that
in vivo water content within leaf was the dominant characteristic for discriminating among the cultivars.
The information generated from vegetation indices depends upon the phenological stage and plant parameter to which the index is most closely related [
15]. This study identified SDA models with different vegetation indices for discriminating among the cultivars at different sampling intervals, Chlorophyll red edge index and EVI in June, red edge ratio in July and PSRI in September, which is indicative of an index influenced by the phenological or plant parameter to which it is most closely related. The PSRI an index proposed to be sensitive to the senescence phase of plant development had the lowest Wilk’s lambda value in differentiating among the cultivars. This index was most sensitive to the senescence phase in being able to discriminate among the cultivars at leaf level. The PSRI defined as (Red
660 − Green
510)/NIR
760, takes advantage of the reflectance relationships in red, green and near infrared regions of the spectrum [
15]. The relatively large Wilk’s lambda value also suggests that the degree of separation was poor in the months of June and July. The low accuracy in classifying the cultivars during these months confirms this. Furthermore, chlorophyll/pigment related indices that most closely match to the leaf chlorophyll content were most dominant in discriminating among the N treatments. Nitrogen concentration in green plants is related to chlorophyll content [
38]. Studies have shown that leaf chlorophyll content can indicate N stress in corn [
39], rice [
40,
41], and sorghum [
42]. The indices identified that discriminated among the N treatments were dominated by chlorophyll/pigment related computed indices (SIPI, MCARI and TCARI), again showing relation to the parameter the index is most closely related. TCARI an index that is sensitive to changes in chlorophyll concentrations takes advantage of the reflectance relationships in the red, red edge and green regions of the spectrum occurred in both July and August model for discrimination. The dominant plant pigments were the chlorophyll and carotenoids explaining over 70% of the variability in PC1 (
Table 4). In general, chlorophyll reflectance and absorption is associated with a green peak (∼550 nm) followed by a decrease in red reflectance (∼650–690 nm). High spectral resolution measurements of chlorophyll in the red edge region (700–795 nm) was found to detect trace quantities of green vegetation [
43]. A leaf simulated reflectance analysis using PROSPECT model was conducted by Haboudane
et al. [
38]. They reported a negative correlation between TCARI and chlorophyll concentration over a range of (10–70 μg/cm
2), and a positive one at concentration below 10 μg/cm
2. This indicates that TCARI is highly sensitive to low concentration of chlorophyll.





{kind=link}
{kind=link}
{kind=link}