Validating and Linking the GIMMS Leaf Area Index (LAI3g) with Environmental Controls in Tropical Africa

 ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
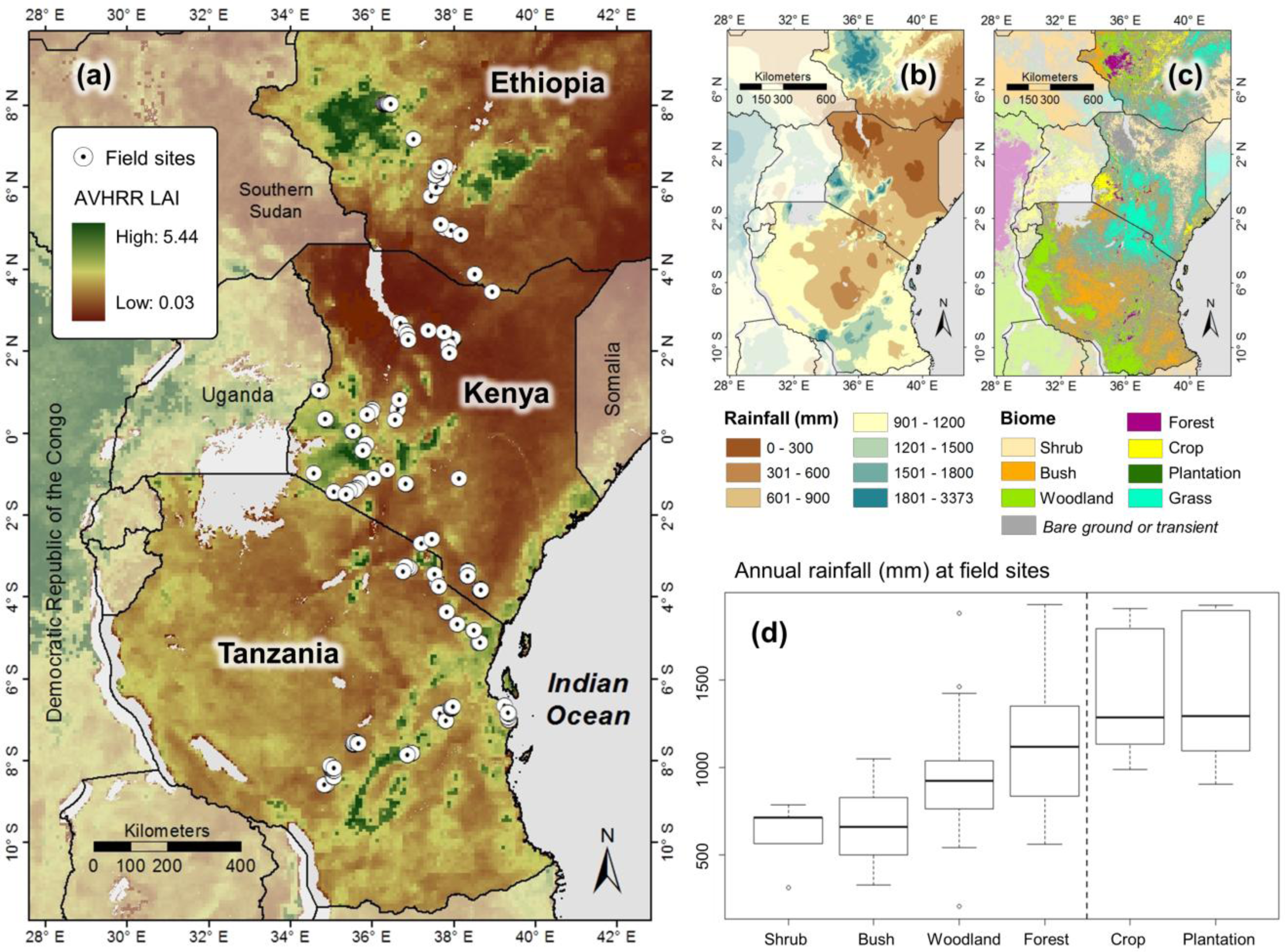
2.1. Study Region
2.2. Ground-Measurements of LAI
2.3. Validation of GIMMS LAI3g
2.4. LAI Response to Environmental Gradients
3. Results
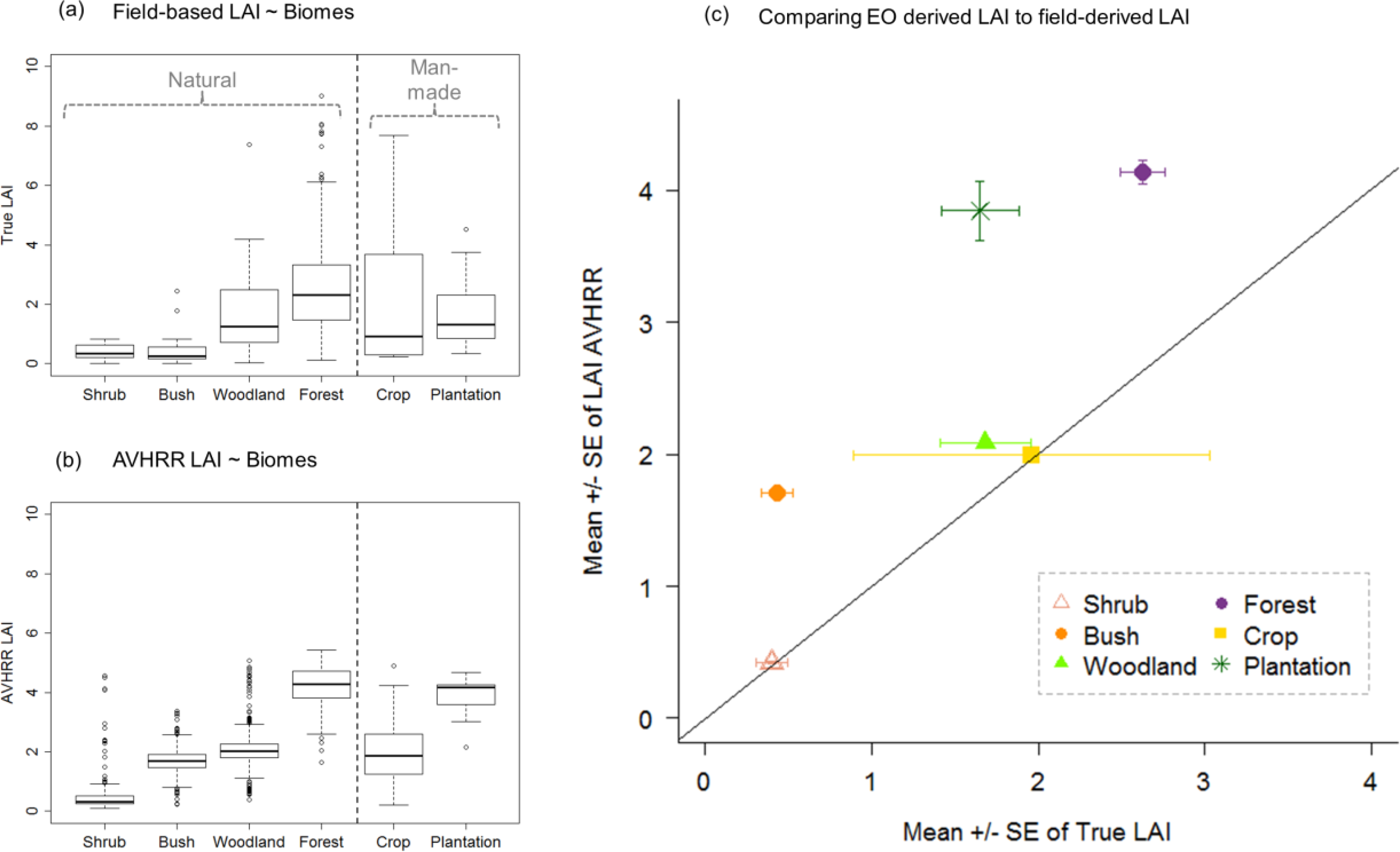
3.1. LAI Distribution within and between Biomes
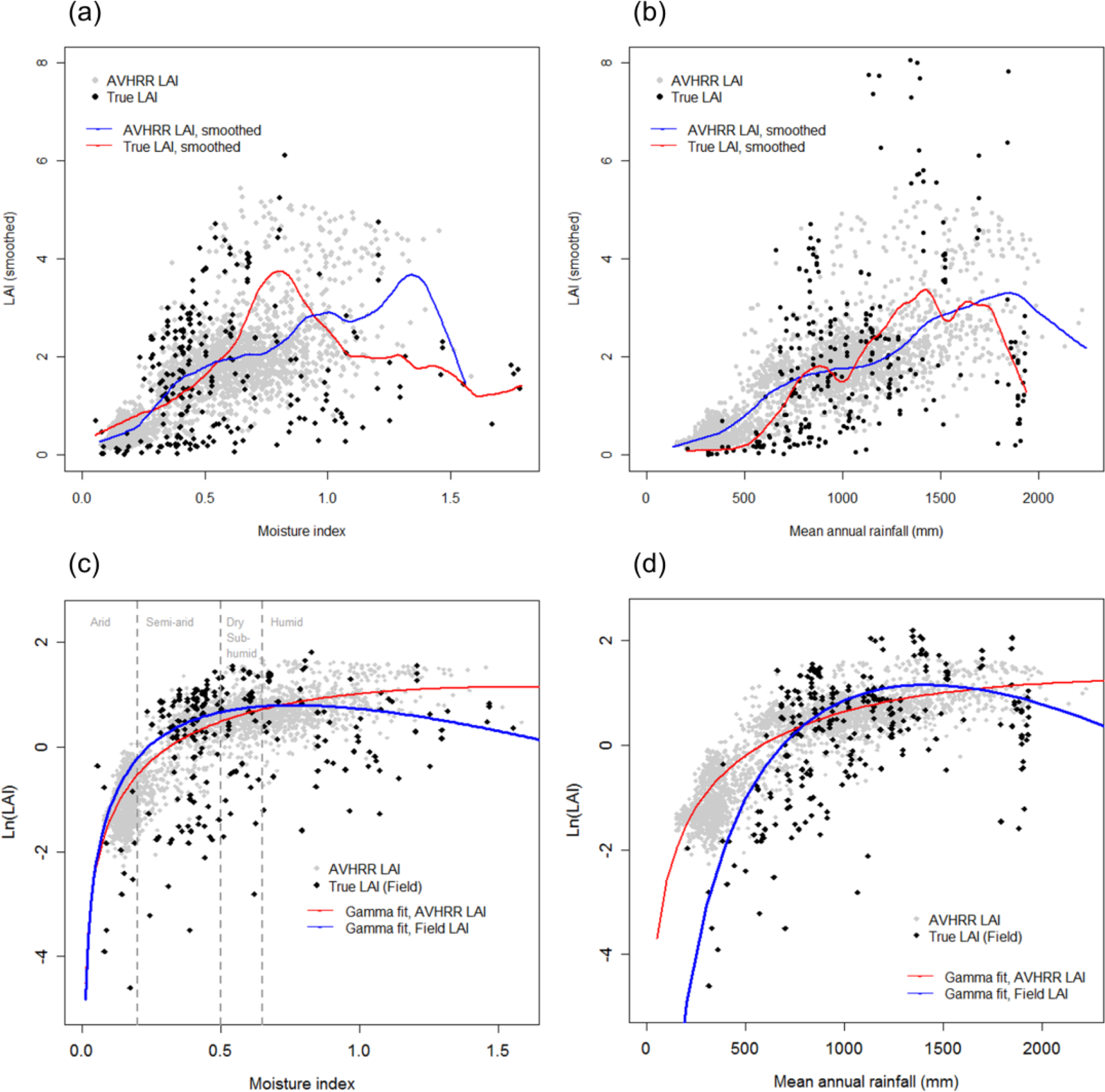
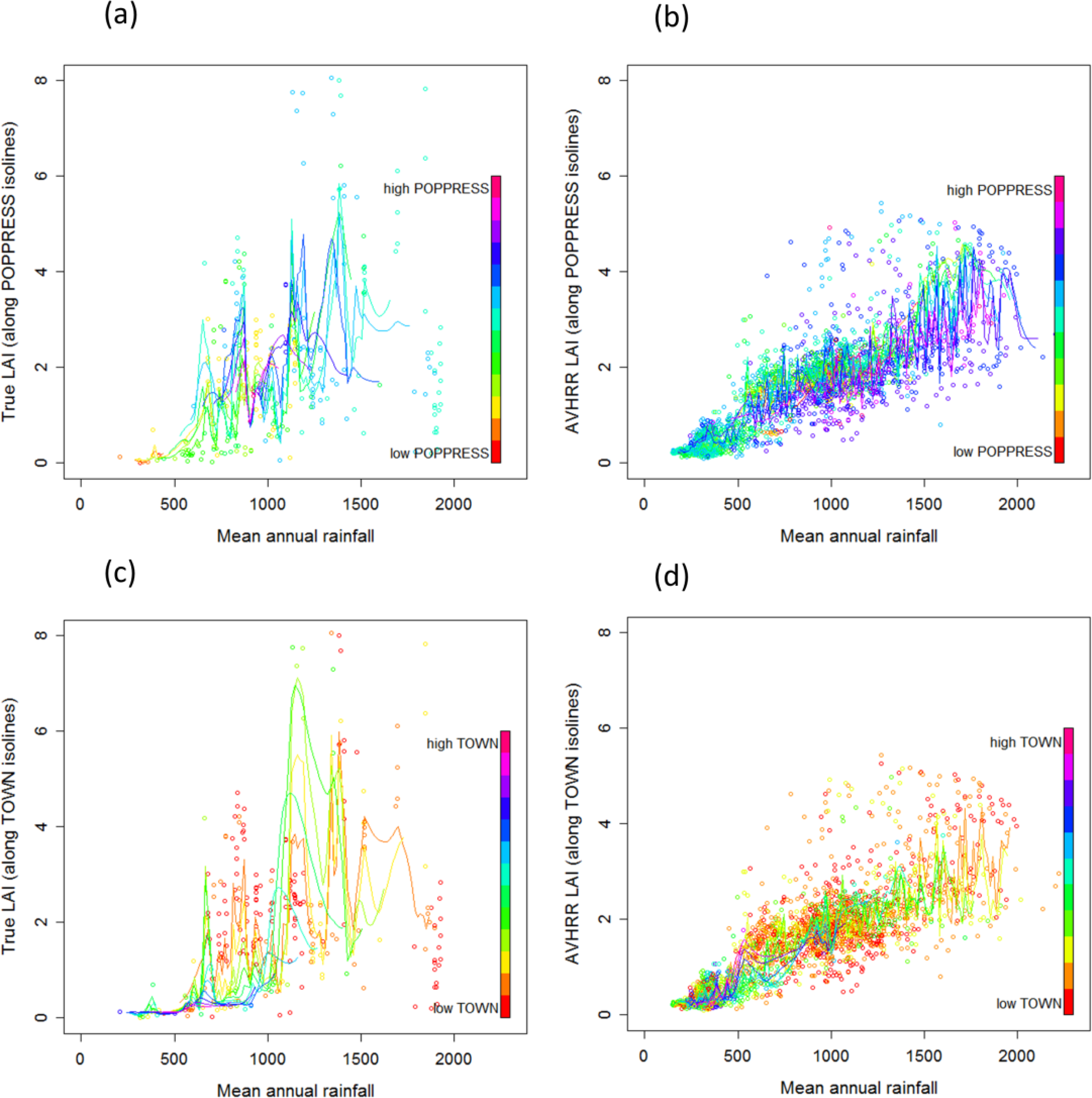
3.2. Distribution of LAI Response to Environmental Variation
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, J.M.; Pavlic, G.; Brown, L.; Cihlar, J.; Leblanc, S.; White, H.; Hall, R.; Peddle, D.; King, D.; Trofymow, J.; et al. Derivation and validation of Canada-wide coarse-resolution leaf area index maps using high-resolution satellite imagery and ground measurements. Remote Sens. Environ 2002, 80, 165–184. [Google Scholar]
- Sabater, J.M.; Rüdiger, C.; Calvet, J.C.; Fritz, N.; Jarlan, L.; Kerr, Y. Joint assimilation of surface soil moisture and LAI observations into a land surface model. Agric. For. Meteorol 2008, 148, 1362–1373. [Google Scholar] [Green Version]
- Sellers, P.; Dickinson, R.; Randall, D.; Betts, A.; Hall, F.; Berry, J.; Collatz, G.; Denning, A.; Mooney, H.; Nobre, C.; et al. Modeling the exchanges of energy, water, and carbon between continents and the atmosphere. Science 1997, 275, 502–509. [Google Scholar]
- Launay, M.; Guérif, M. Ability for a model to predict crop production variability at the regional scale: An evaluation for sugar beet. Agronomie 2003, 23, 135–146. [Google Scholar]
- Neitsch, S.; Arnold, J.; Kiniry, J.; Williams, J.; King, K. Soil and Water Assessment Tool: Theoretical Documentation Version 2005; Blackland Research Centre: Temple, TX, USA, 2005. [Google Scholar]
- Matricardi, E.A.; Skole, D.L.; Pedlowski, M.A.; Chomentowski, W.; Fernandes, L.C. Assessment of tropical forest degradation by selective logging and fire using Landsat imagery. Remote Sens. Environ 2010, 114, 1117–1129. [Google Scholar]
- Pfeifer, M.; Gonsamo, A.; Disney, M.; Pellikka, P.; Marchant, R. Leaf area index for biomes of the Eastern Arc Mountains: Landsat and SPOT observations along precipitation and altitude gradients. Remote Sens. Environ 2012, 118, 103–115. [Google Scholar]
- Fang, H.; Jiang, C.; Li, W.; Wei, S.; Baret, F.; Chen, J.M.; Garcia-Haro, J.; Liang, S.; Liu, R.; Myneni, R.B.; et al. Characterization and intercomparison of global moderate resolution leaf area index (LAI) products: Analysis of climatologies and theoretical uncertainties. J. Geophys. Res.: Biogeosci 2013, 118, 529–548. [Google Scholar]
- Myneni, R.; Hoffman, S.; Knyazikhin, Y.; Privette, J.; Glassy, J.; Tian, Y.; Wang, Y.; Song, X.; Zhang, Y.; Smith, G.; et al. Global products of vegetation leaf area and fraction absorbed PAR from year one of MODIS data. Remote Sens. Environ 2002, 83, 214–231. [Google Scholar]
- Baret, F.; Hagolle, O.; Geiger, B.; Bicheron, P.; Miras, B.; Huc, M.; Berthelot, B.; Niño, F.; Weiss, M.; Samain, O.; et al. LAI, fAPAR and fCover CYCLOPES global products derived from VEGETATION: Part 1: Principles of the algorithm. Remote Sens. Environ 2007, 110, 275–286. [Google Scholar]
- Myneni, R.B.; Ramakrishna, R.; Nemani, R.; Running, S. Estimation of global leaf area index and absorbed PAR using radiative transfer models. IEEE Trans. Geosci. Remote Sens 1997, 35, 1380–1393. [Google Scholar]
- Fernandes, R.; Butson, C.; Leblanc, S.; Latifovic, R. Landsat-5 TM and Landsat-7 ETM+ based accuracy assessment of leaf area index products for Canada derived from SPOT-4 VEGETATION data. Can. J. Remote Sens 2003, 29, 241–258. [Google Scholar]
- Myneni, R.B.; Hall, F.G.; Sellers, P.J.; Marshak, A.L. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remote Sens 1995, 33, 481–486. [Google Scholar]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI) 3g and Fraction of Photosynthetically Active Radiation (FPAR) 3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens 2013, 5, 927–948. [Google Scholar]
- Mao, J.; Shi, X.; Thornton, P.E.; Hoffman, F.M.; Zhu, Z.; Myneni, R.B. Global latitudinal-asymmetric vegetation growth trends and their driving mechanisms: 1982–2009. Remote Sens 2013, 5, 1484–1497. [Google Scholar]
- Cook, B.I.; Pau, S. A global assessment of long-term greening and browning trends in pasture lands using the GIMMS LAI3g dataset. Remote Sens 2013, 5, 2492–2512. [Google Scholar]
- Pfeifer, M.; Platts, P.J. Ground Measurements of Leaf Area Index in Africa, Version 1.5. 2014. Available online: http://www.york.ac.uk/environment/research/kite/resources/ (accessed on 29 January 2014).
- Green, J.M.; Larrosa, C.; Burgess, N.D.; Balmford, A.; Johnston, A.; Mbilinyi, B.P.; Platts, P.J.; Coad, L. Deforestation in an African biodiversity hotspot: Extent, variation and the effectiveness of protected areas. Biol. Conserv 2013, 164, 62–72. [Google Scholar]
- Pfeifer, M.; Burgess, N.D.; Swetnam, R.D.; Platts, P.J.; Willcock, S.; Marchant, R. Protected areas: Mixed success in conserving East Africa’s evergreen forests. PLoS One 2012, 7, e39337. [Google Scholar] [CrossRef]
- Pfeifer, M.; Platts, P.J.; Burgess, N.D.; Swetnam, R.D.; Willcock, S.; Lewis, S.L.; Marchant, R. Land use change and carbon fluxes in East Africa quantified using earth observation data and field measurements. Environ. Conserv 2013, 40, 242–252. [Google Scholar]
- Platts, P.J.; Burgess, N.D.; Gereau, R.; Lovett, J.; Marshall, A.R.; McCLEAN, C.J.; Pellikka, P.K.; Swetnam, R.D.; Marchant, R. Delimiting tropical mountain ecoregions for conservation. Environ. Conserv 2011, 38, 312–324. [Google Scholar]
- Zhang, Y.; Chen, J.M.; Miller, J.R. Determining digital hemispherical photograph exposure for leaf area index estimation. Agric. For. Meteorol 2005, 133, 166–181. [Google Scholar]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol 2004, 121, 19–35. [Google Scholar]
- Gonsamo, A.; Pellikka, P. Methodology comparison for slope correction in canopy leaf area index estimation using hemispherical photography. For. Ecol. Manag 2008, 256, 749–759. [Google Scholar]
- VALERI. Available online: http://w3.avignon.inra.fr/valeri (accessed on 10 August 2013).
- Jonckheere, I.; Muys, B.; Coppin, P. Allometry and evaluation of in situ optical LAI determination in Scots pine: A case study in Belgium. Tree Physiol 2005, 25, 723–732. [Google Scholar]
- Weiss, M.; Baret, F. CAN-EYE V6. 1 USER MANUAL. 2010. Available online: http://www6.paca.inra.fr/can-eye/Download (accessed on 29 January 2014).
- Weiss, M.; Baret, F.; Myneni, R.B.; Pragnère, A.; Knyazikhin, Y. Investigation of a model inversion technique to estimate canopy biophysical variables from spectral and directional reflectance data. Agronomie 2000, 20, 3–22. [Google Scholar]
- Garrigues, S.; Lacaze, R.; Baret, F.; Morisette, J.; Weiss, M.; Nickeson, J.; Fernandes, R.; Plummer, S.; Shabanov, N.; Myneni, R.B.; et al. Validation and intercomparison of global Leaf Area Index products derived from remote sensing data. J. Geophys. Res.: Biogeosci 2008, 113. [Google Scholar] [CrossRef]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ 2010, 114, 168–182. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna; Available online: http://www.r-project.org/ (accessed on 1 September 2013).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol 2005, 25, 1965–1978. [Google Scholar]
- Hargreaves, G.; Allen, R. History and evaluation of Hargreaves evapotranspiration equation. J. Irrig. Drain. Eng 2003, 129, 53–63. [Google Scholar]
- Rodriguez, E.; Morris, C.S.; Belz, J.E. A global assessment of the SRTM performance. Photogramm. Eng. Remote Sens 2006, 72, 249–260. [Google Scholar]
- Greve, M.; Lykke, A.M.; Fagg, C.W.; Bogaert, J.; Friis, I.; Marchant, R.; Marshall, A.R.; Ndayishimiye, J.; Sandel, B.S.; Sandom, C.; et al. Continental-scale variability in browser diversity is a major driver of diversity patterns in acacias across Africa. J. Ecol 2012, 100, 1093–1104. [Google Scholar]
- IUCN; UNEP-WCMC. The World Database on Protected Areas (WDPA); UNEP-WCMC: Cambridge, UK, 2010; Available online: http://www.protectedplanet.net (accessed on 29 October 2013).
- GeoNetwork. Available online: http://www.fao.org/geonetwork/ (accessed on 10 July 2013).
- Linard, C.; Gilbert, M.; Snow, R.W.; Noor, A.M.; Tatem, A.J. Population distribution, settlement patterns and accessibility across Africa in 2010. PLoS One 2012, 7, e31743. [Google Scholar] [CrossRef]
- Tatem, A.J.; Noor, A.M.; von Hagen, C.; di Gregorio, A.; Hay, S.I. High resolution population maps for low income nations: Combining land cover and census in East Africa. PLoS One 2007, 2, e1298. [Google Scholar] [CrossRef]
- Platts, P.J. Spatial Modelling, Phytogeography and Conservation in the Eastern Arc Mountains of Tanzania and Kenya. Ph.D. Thesis, Environment Department, University of York, York, UK. 2012. [Google Scholar]
- Grueber, C.; Nakagawa, S.; Laws, R.; Jamieson, I. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol 2011, 24, 699–711. [Google Scholar]
- Kalácska, M.; Sánchez-Azofeifa, G.A.; Rivard, B.; Calvo-Alvarado, J.C.; Journet, A.; Arroyo-Mora, J.P.; Ortiz-Ortiz, D. Leaf area index measurements in a tropical moist forest: A case study from Costa Rica. Remote Sens. Environ 2004, 91, 134–152. [Google Scholar]
- Van Leeuwen, W.J.D.; Orr, B.J.; Marsh, S.E.; Herrmann, S.M. Multi-sensor NDVI data continuity: Uncertainties and implications for vegetation monitoring applications. Remote Sens. Environ 2006, 100, 67–81. [Google Scholar]
- Morisette, J.T.; Baret, F.; Privette, J.L.; Myneni, R.B.; Nickeson, J.E.; Garrigues, S.; Shabanov, N.V.; Weiss, M.; Fernandes, R.A.; Leblanc, S.G.; et al. Validation of global moderate-resolution LAI products: A framework proposed within the CEOS land product validation subgroup. IEEE Trans. Geosci. Remote Sens 2006, 44, 1804–1817. [Google Scholar]
- Davenport, M.; Nicholson, S. On the relation between rainfall and the normalized difference vegetation index for diverse vegetation types in East Africa. Int. J. Remote Sens 1993, 14, 2369–2389. [Google Scholar]
- Herrmann, S.M.; Anyamba, A.; Tucker, C.J. Recent trends in vegetation dynamics in the African Sahel and their relationship to climate. Glob. Environ. Chang 2005, 15, 394–404. [Google Scholar]
- Pachauri, R.K. Climate Change 2007. Synthesis Report; Contribution of Working Groups I, II and III to the Fourth Assessment Report; Cambridge University Press: Cambridge, UK/New York, NY, USA, 2008. [Google Scholar]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in Africa by coupling MODIS and TRMM instruments. J. Geophys. Res.: Atmos 2005, 110. [Google Scholar] [CrossRef]
- Nicholson, S.; Farrar, T. The influence of soil type on the relationships between NDVI, rainfall, and soil moisture in semiarid Botswana. I. NDVI response to rainfall. Remote Sens. Environ 1994, 50, 107–120. [Google Scholar]
- Tan, B.; Woodcock, C.; Hu, J.; Zhang, P.; Ozdogan, M.; Huang, D.; Yang, W.; Knyazikhin, Y.; Myneni, R.B. The impact of gridding artifacts on the local spatial properties of MODIS data: Implications for validation, compositing, and band-to-band registration across resolutions. Remote Sens. Environ 2006, 105, 98–114. [Google Scholar]
- Wang, Y.; Woodcock, C.E.; Buermann, W.; Stenberg, P.; Voipio, P.; Smolander, H.; Häme, T.; Tian, Y.; Hu, J.; Knyazikhin, Y.; et al. Evaluation of the MODIS LAI algorithm at a coniferous forest site in Finland. Remote Sens. Environ 2004, 91, 114–127. [Google Scholar]
- Buermann, W.; Wang, Y.; Dong, J.; Zhou, L.; Zeng, X.; Dickinson, R.E.; Potter, C.S.; Myneni, R.B. Analysis of a multiyear global vegetation leaf area index data set. J. Geophys. Res.: Atmos 2002, 107. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | N | Date | Country | Camera | Lens | PI |
|---|---|---|---|---|---|---|
| ID01 | 37 | January 2010 | TZA, KEN | Nikon D5000 | Nikkor F2.8 | MP, RAM |
| ID02 | 43 | July 2010 | TZA | Canon EOS 450D | Sigma 4.5 F2.8 | MP, SW * |
| ID03 | 58 | January 2010 | KEN | Nikon D5000 SLR | Sigma 4.5 F2.8 | PKEP |
| ID04 | 23 | January 2007 | KEN | Nikon 8800 VR | Nikon FC-E9 | AG |
| ID05 | 31 | June 2011 | KEN | Canon EOS 450D | Sigma 4.5 F2.8 | MP, ACS * |
| ID06 | 32 | June 2012 | KEN, ETH | Nikon D3100 | Sigma 4.5 F2.8 | MP, PJP, ACS * |
| ID06a | 8 | June 2012 | ETH | Nikon D5000 | Nikkor F2.8 | RAM |
| ID07 | 20 | August 2011 | TZA | Nikon D3100 | Sigma 4.5 F2.8 | HS * |
| ID08 | 11 | November 2012 | KEN | Nikon D800 | Sigma 4.5 F2.8 | RAM |
| ID10 | 31 | May to June 2012 | KEN | Nikon D800 | Sigma 4.5 F2.8 | MP, RAM, PJP, LJ * |
| ID12 | 14 | June 2012 | ETH | Nikon D5000 | Nikkor F2.8 | DD |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | −0.39 | - | |||||||||
| 3 | 0.64 | NS | - | ||||||||
| 4 | −0.16 | 0.80 | 0.13 | ||||||||
| 5 | 0.64 | −0.14 | 0.52 | NS | - | ||||||
| 6 | 0.57 | NS | 0.53 | NS | 0.91 | ||||||
| 7 | 0.48 | NS | 0.47 | 0.19 | 0.18 | 0.18 | - | ||||
| 8 | NS | −0.31 | −0.13 | −0.37 | 0.13 | NS | −0.29 | - | |||
| 9 | 0.42 | −0.22 | 0.38 | 0.28 | 0.38 | 0.31 | 0.43 | NS | - | ||
| 10 | −0.26 | −0.14 | −0.41 | −0.15 | −0.19 | −0.24 | −0.26 | 0.29 | NS | - | |
| 11 | 0.25 | NS | 0.36 | 0.16 | 0.16 | NS | NS | NS | 0.23 | −0.14 | - |
| 12 | −0.17 | NS | NS | 0.12 | −0.25 | −0.28 | NS | −0.45 | ns | ns | −0.21 |
| Number | Mean ± SE | Median | IQR | Max | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | |
| Shrub | 9 | 995 | 0.4 ± 0.1 | 0.4 ± 0.0 | 0.4 | 0.3 | 0.4 | 0.3 | 0.8 | 4.6 |
| Bush | 29 | 705 | 0.4 ± 0.1 | 1.7 ± 0.0 | 0.3 | 1.7 | 0.4 | 0.5 | 2.5 | 3.4 |
| Woodland | 32 | 624 | 1.7 ± 0.3 | 2.1 ± 0.0 | 1.3 | 2.0 | 1.7 | 0.5 | 7.4 | 5.1 |
| Forest | 174 | 80 | 2.6 ± 0.1 | 4.1 ± 0.1 | 2.3 | 4.3 | 1.8 | 0.9 | 9.0 | 5.4 |
| Crop | 7 | 294 | 2.0 ± 1.1 | 2.0 ± 0.1 | 0.5 | 1.9 | 2.3 | 1.3 | 7.7 | 4.9 |
| Plantation | 23 | 11 | 1.6 ± 0.2 | 3.8 ± 0.2 | 1.3 | 4.2 | 1.5 | 0.7 | 4.5 | 4.7 |
| PPT | MI | PET | POP-PRESS | Slope | HERB | MI_DQ | PPT_DQ | TOWN | ELE-VATION | ROADS | TR | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | FI | EO | |
| Shrub | 0.86 | 0.64 | 0.76 | 0.64 | 0.76 | 0.57 | 0.72 | 0.54 | 0.78 | 0.59 | 0.41 | 0.60 | 0.43 | 0.60 | 0.54 | 0.59 | 0.67 | 0.53 | 0.84 | 0.59 | 0.34 | 0.49 | 0.39 | 0.53 |
| Bush | 0.49 | 0.52 | 0.43 | 0.49 | 0.26 | 0.39 | 0.65 | 0.35 | 0.76 | 0.33 | 0.38 | 0.41 | 0.30 | 0.44 | 0.39 | 0.45 | 0.48 | 0.36 | 0.36 | 0.38 | 0.25 | 0.33 | 0.30 | 0.38 |
| Wood | 0.35 | 0.53 | 0.48 | 0.49 | 0.40 | 0.42 | 0.33 | 0.31 | 0.36 | 0.41 | 0.40 | 0.37 | 0.60 | 0.55 | 0.51 | 0.54 | 0.45 | 0.35 | 0.32 | 0.42 | 0.31 | 0.32 | 0.35 | 0.35 |
| Forest | 0.39 | 0.17 | 0.27 | 0.17 | 0.31 | 0.36 | 0.39 | 0.19 | 0.25 | 0.14 | 0.37 | 0.25 | 0.22 | 0.23 | 0.41 | 0.27 | 0.30 | 0.16 | 0.22 | 0.22 | 0.12 | 0.19 | 0.29 | 0.19 |
| Crop | 0.55 | 0.55 | 0.58 | 0.48 | 0.85 | 0.27 | 0.53 | 0.28 | 0.69 | 0.25 | 0.47 | 0.44 | 0.51 | 0.67 | 0.70 | 0.68 | 0.51 | 0.24 | 0.69 | 0.61 | 0.46 | 0.55 | 0.65 | 0.59 |
| Plantation | 0.42 | 0.40 | 0.36 | 0.36 | 0.38 | 0.39 | 0.47 | 0.53 | 0.29 | 0.45 | 0.45 | 0.36 | 0.56 | 0.56 | 0.47 | 0.55 | 0.37 | 0.63 | 0.40 | 0.46 | 0.43 | 0.54 | 0.40 | 0.44 |
| All | 0.40 | 0.66 | 0.23 | 0.64 | 0.36 | 0.41 | 0.33 | 0.51 | 0.32 | 0.48 | 0.23 | 0.48 | 0.18 | 0.66 | 0.32 | 0.66 | 0.39 | 0.46 | 0.16 | 0.59 | 0.14 | 0.39 | 0.33 | 0.40 |
| RVI; LAITrue Models | RVI; LAIavhrr Models | Sign of Estimated GLM Coefficients | ||||
|---|---|---|---|---|---|---|
| GAM | GLM | GAM | GLM | Field | EO | |
| Biome type | 1.0 | 1.0 | 1.0 | 1.0 | Positive | Positive |
| Mean annual rainfall (PPT) | 0.49 | 1.0 a | 0.50 | P1: +; P2: − | ||
| Potential evapotranspiration | 0.50 | 0.28 | 1.0 | 0.22 | Negative | Positive |
| Temperature range | 0.33 | 0.61 | 1.0 | Negative | ||
| Population pressure (POPPRESS) | 0.53 | 1.0 | 1.0 | 0.30 | Positive | Positive |
| Distance to roads | 0.20 | 0.12 | 0.32 | 1.0 | Positive | Positive |
| Distance to towns | 1.0 | 0.11 | 1.0 | 0.57 | Negative | Negative |
| Protected | 1.0 | 1.0 | 1.0 | Positive | ||
| Elevation | 1.0 | 1.0 | 1.0 | Negative | ||
| Slope | 0.62 | 1.0 | 1.0 | Positive | ||
| Latitude/Longitude | 1.0 | 1.0 | 1.0 | 1.0 | Positive | Positive |
| PPT × POPPRESS | 1.0 | - | 1.0 | - | ||
| Biomes:Slope | - | 1.0 | - | - | ||
| AIC | 479.5–481.4 | 591.1–593.1 | 912.6–914.1 | −112.3–−110.4 | ||
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pfeifer, M.; Lefebvre, V.; Gonsamo, A.; Pellikka, P.K.E.; Marchant, R.; Denu, D.; Platts, P.J. Validating and Linking the GIMMS Leaf Area Index (LAI3g) with Environmental Controls in Tropical Africa. Remote Sens. 2014, 6, 1973-1990. https://doi.org/10.3390/rs6031973
Pfeifer M, Lefebvre V, Gonsamo A, Pellikka PKE, Marchant R, Denu D, Platts PJ. Validating and Linking the GIMMS Leaf Area Index (LAI3g) with Environmental Controls in Tropical Africa. Remote Sensing. 2014; 6(3):1973-1990. https://doi.org/10.3390/rs6031973
Chicago/Turabian StylePfeifer, Marion, Veronique Lefebvre, Alemu Gonsamo, Petri K. E. Pellikka, Rob Marchant, Dereje Denu, and Philip J. Platts. 2014. "Validating and Linking the GIMMS Leaf Area Index (LAI3g) with Environmental Controls in Tropical Africa" Remote Sensing 6, no. 3: 1973-1990. https://doi.org/10.3390/rs6031973